The Effects of β-Hydroxy-β-Methylbutyrate (HMB) on Chemotaxis, Phagocytosis, and Oxidative Burst of Peripheral Blood Granulocytes and Monocytes in Goats

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Design
2.3. Sample Collection
2.4. Determination of the Chemotactic Activity of Blood Granulocytes and Monocytes in Goats with the MIGRATEST® Kit
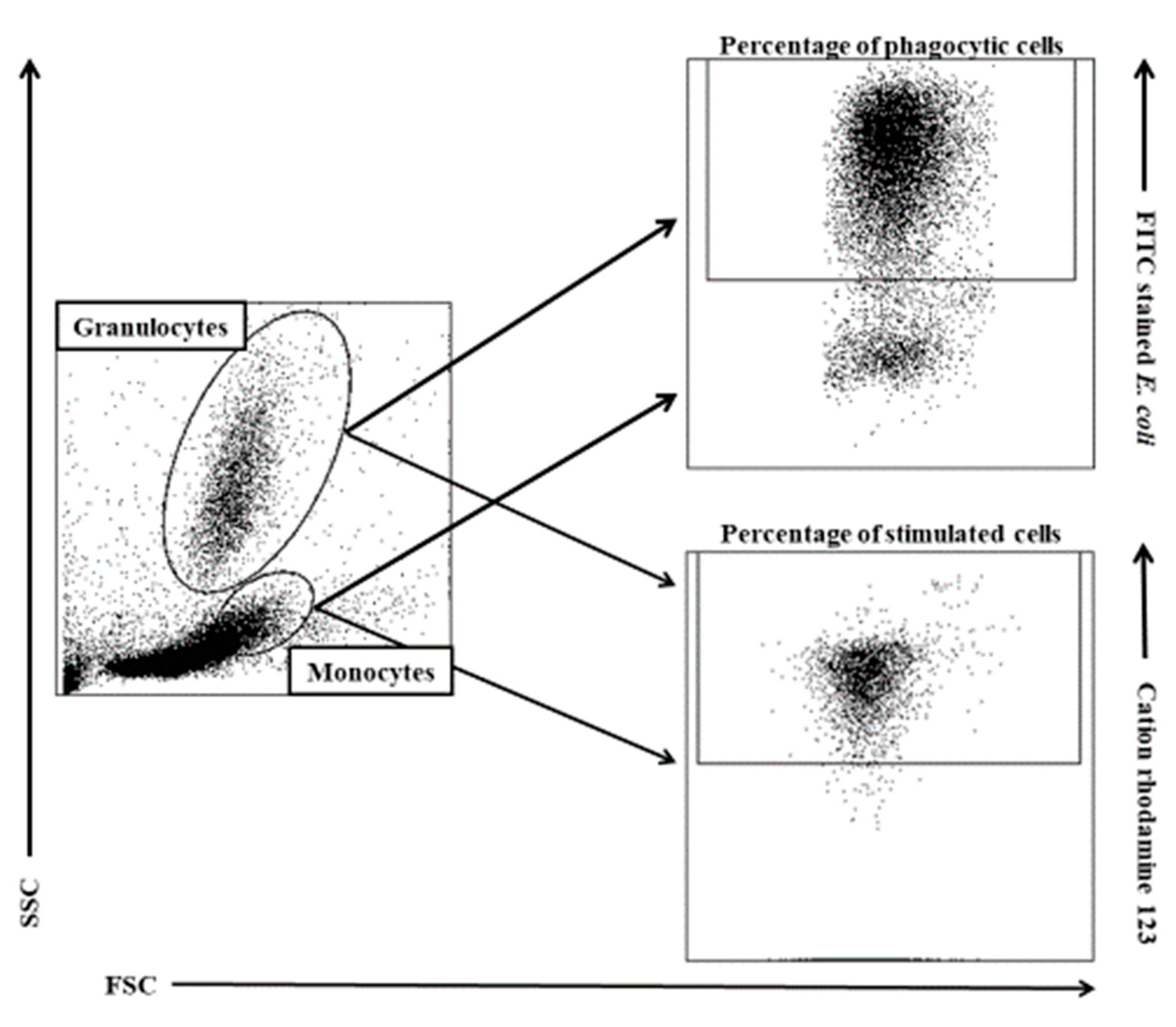
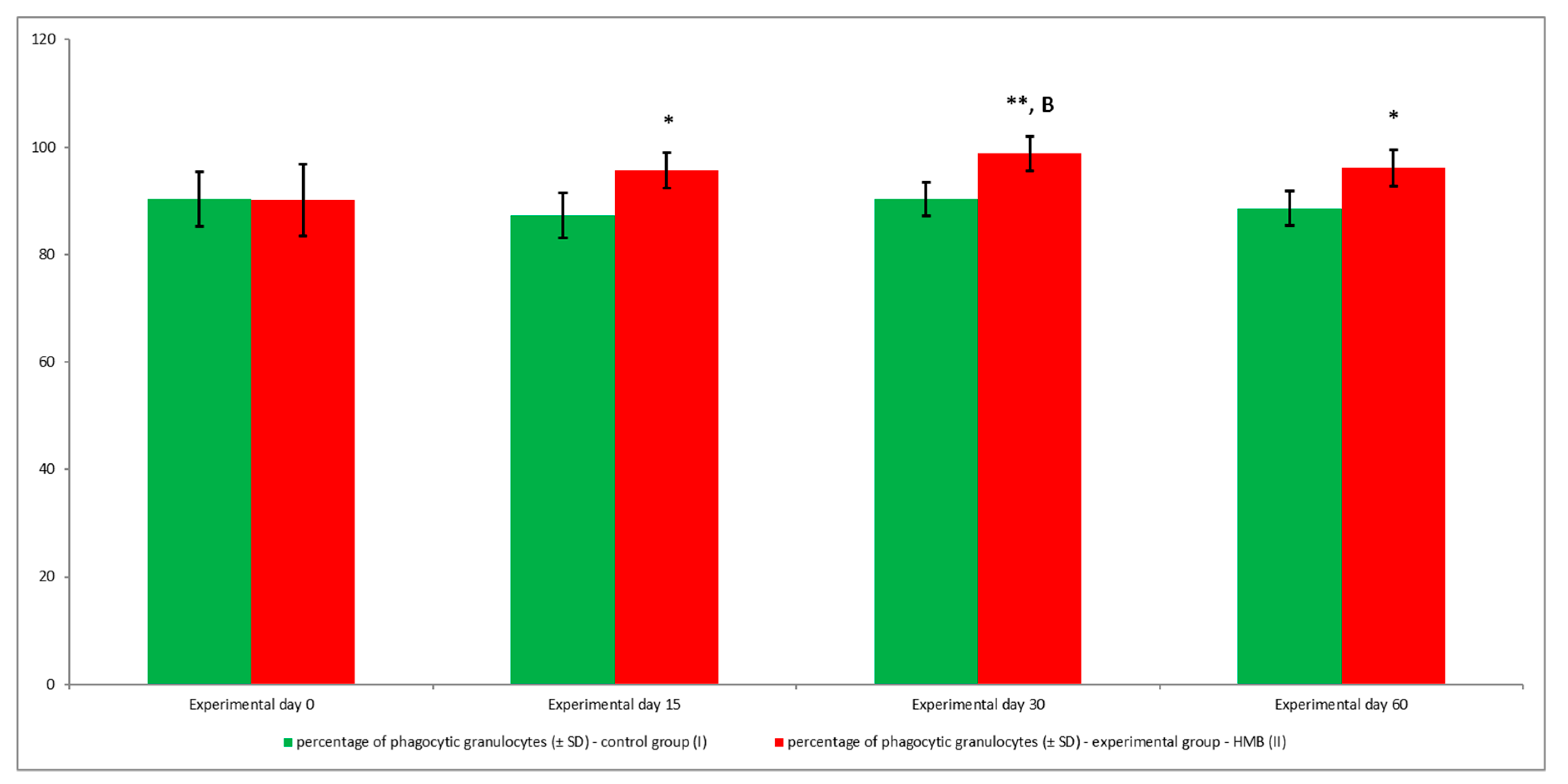
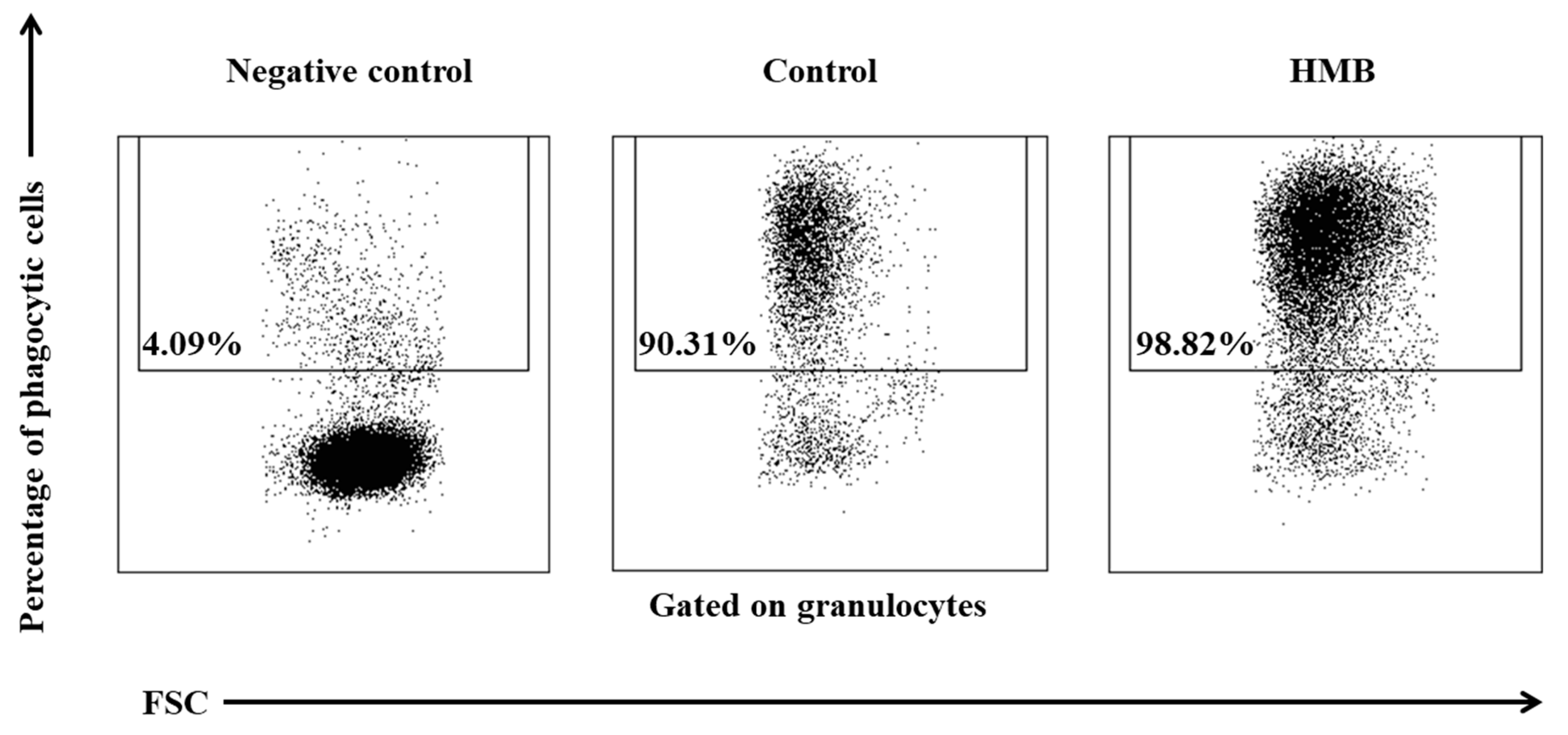
2.5. Determination of the Phagocytic Activity of Blood Granulocytes and Monocytes in Goats with the PHAGOTEST® Kit
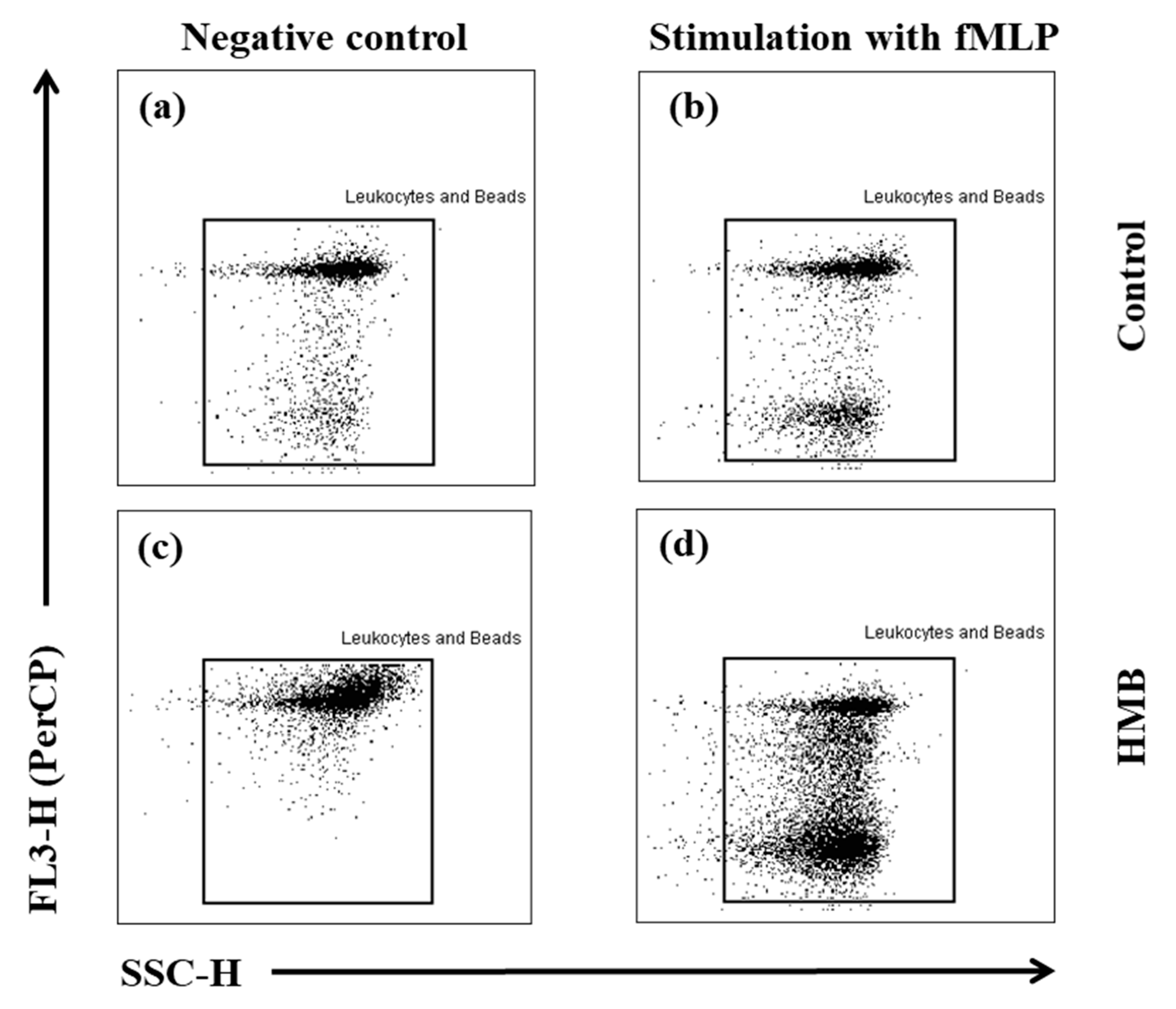
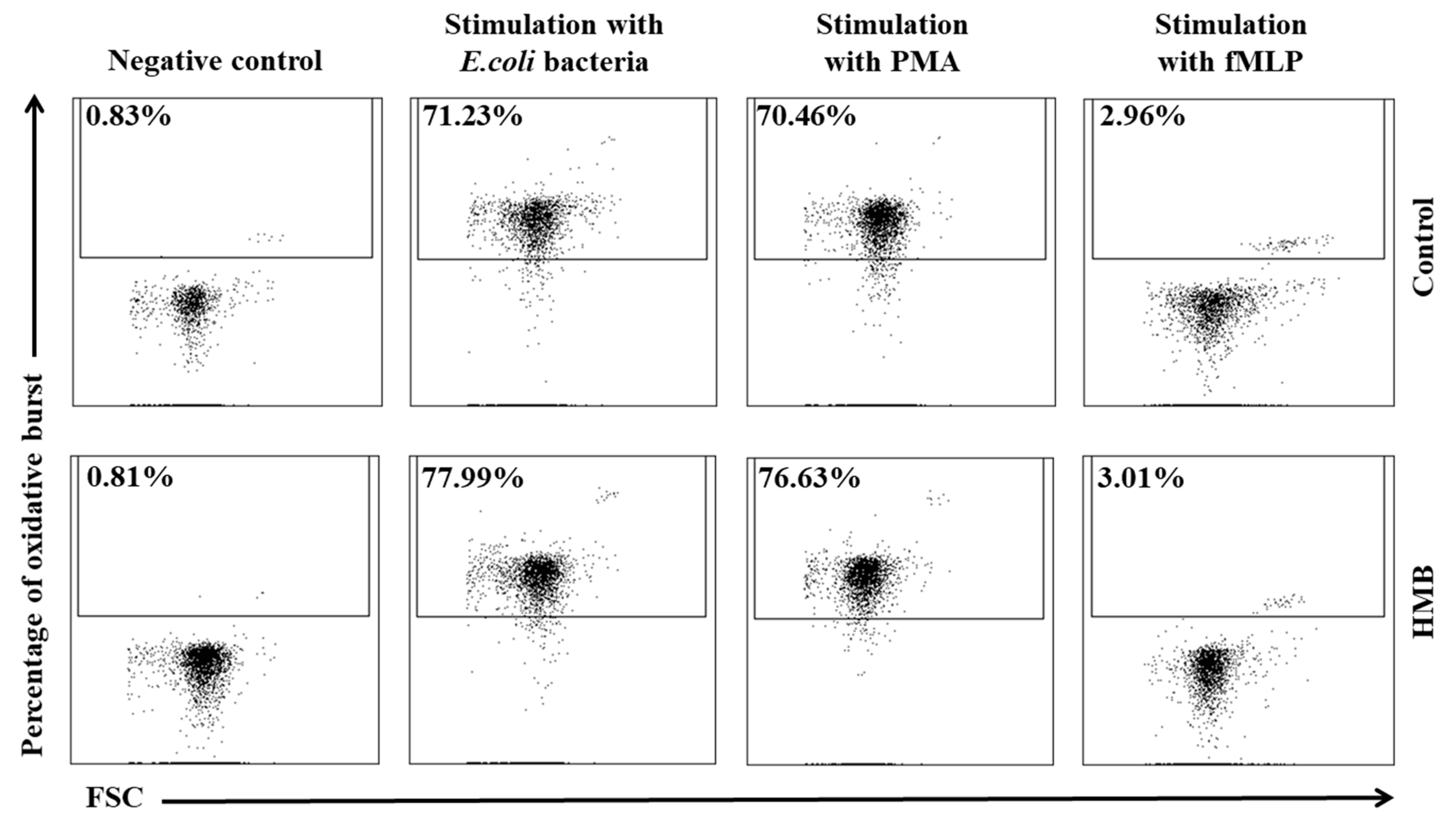
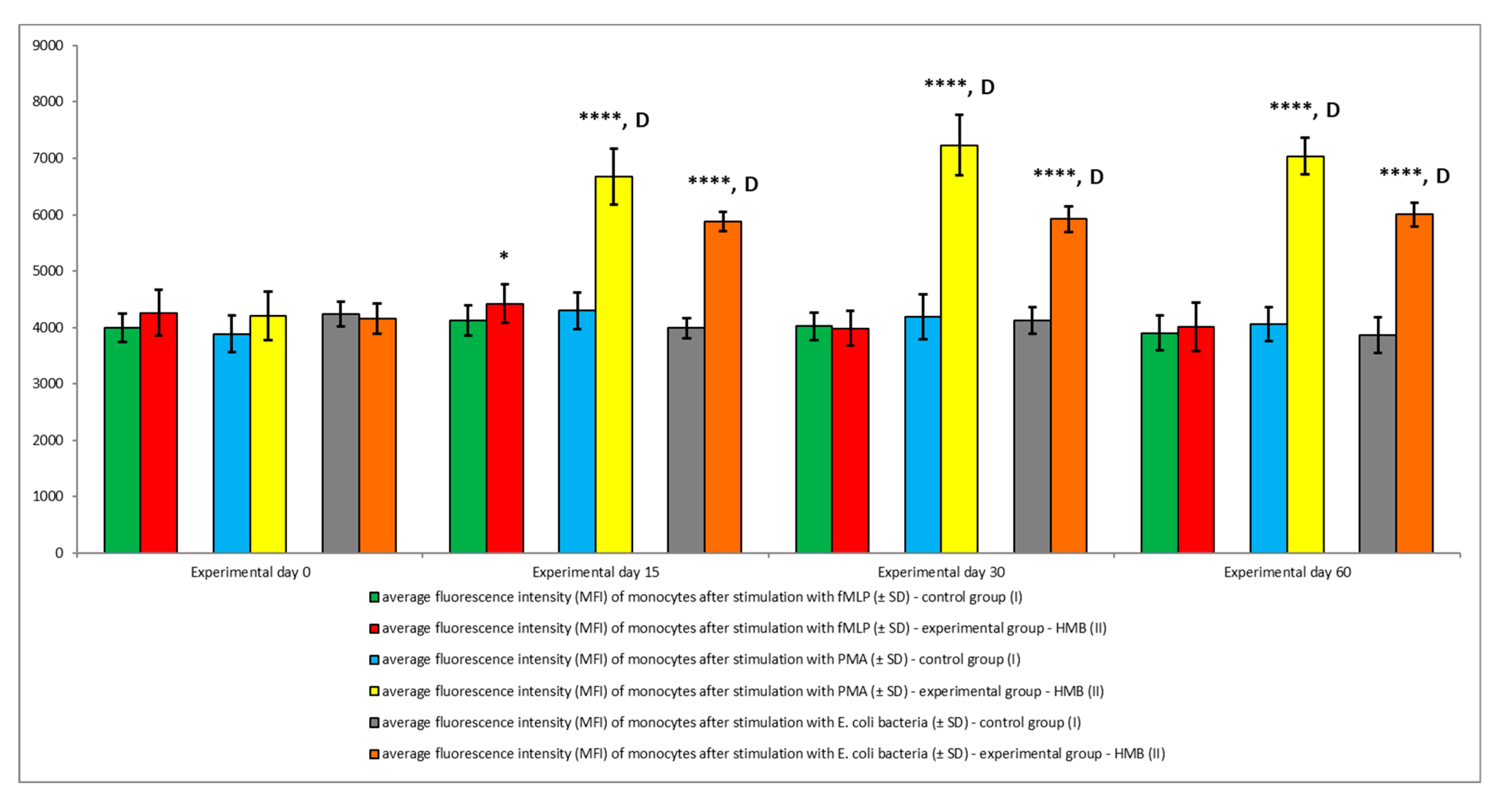
2.6. Determination of the Oxidative Metabolism of Blood Granulocytes and Monocytes in Goats with the BURSTTEST® Kit
2.7. FACS Acquisition and Analysis
2.8. Statistical Analysis
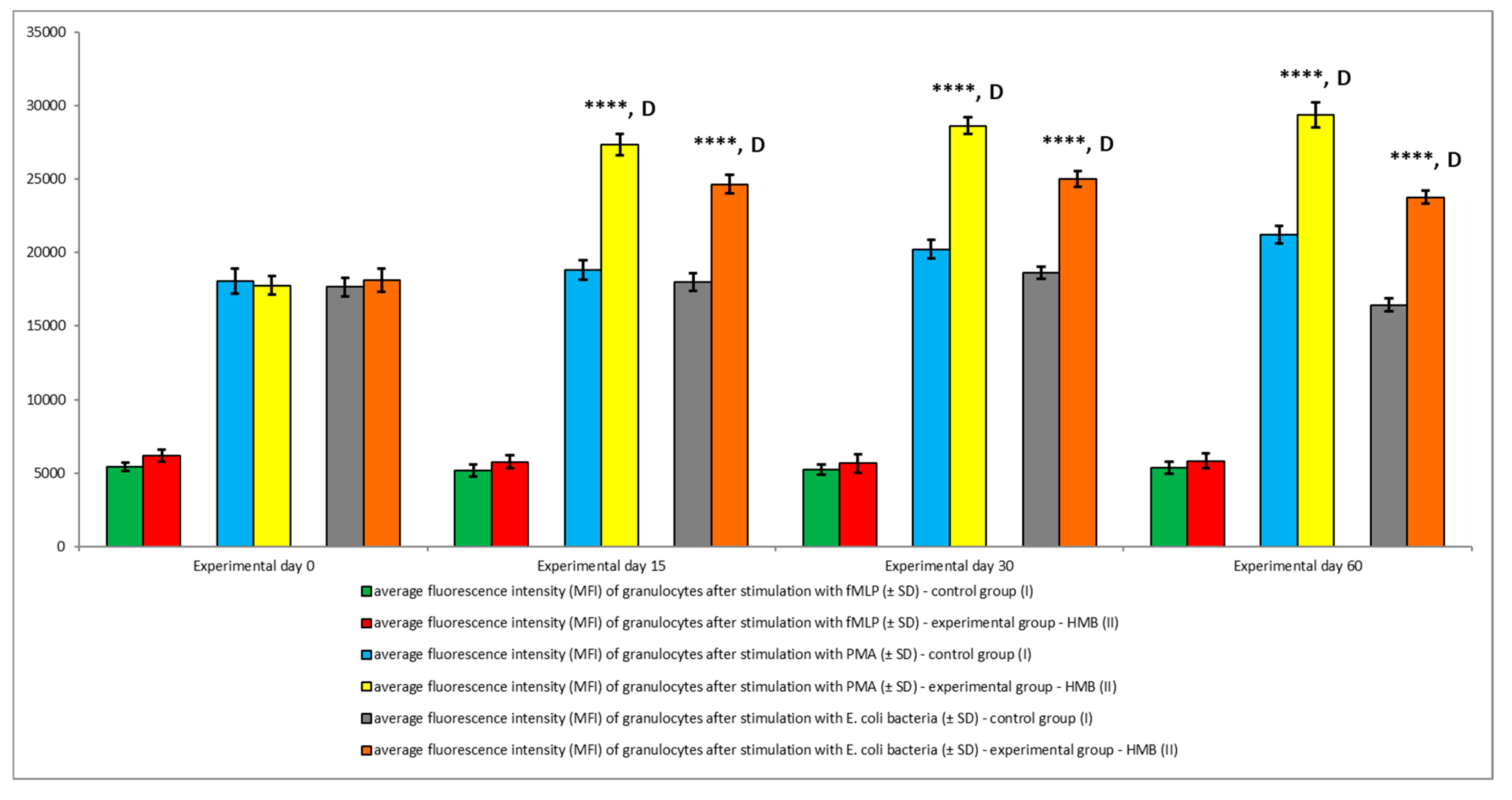
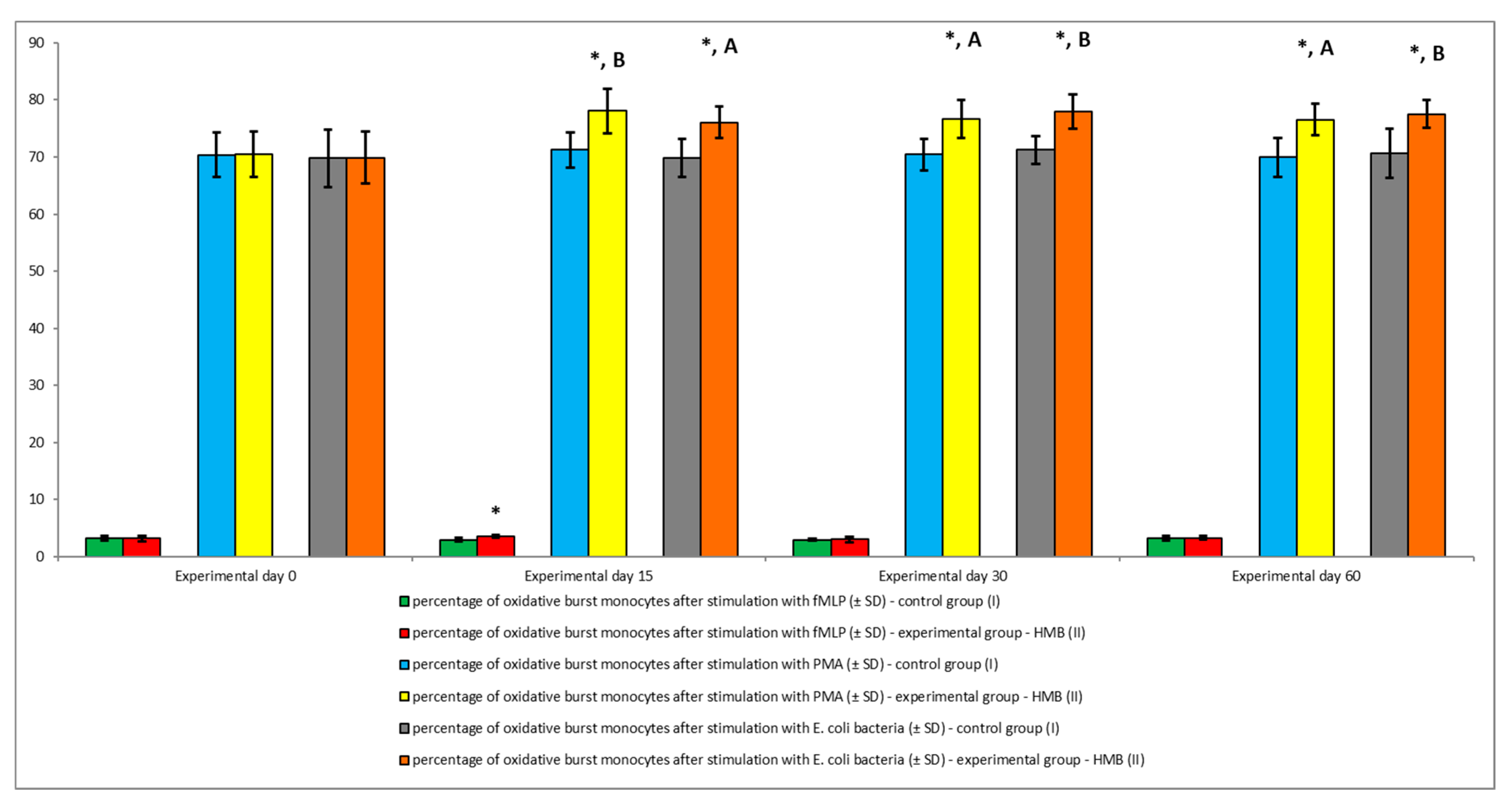
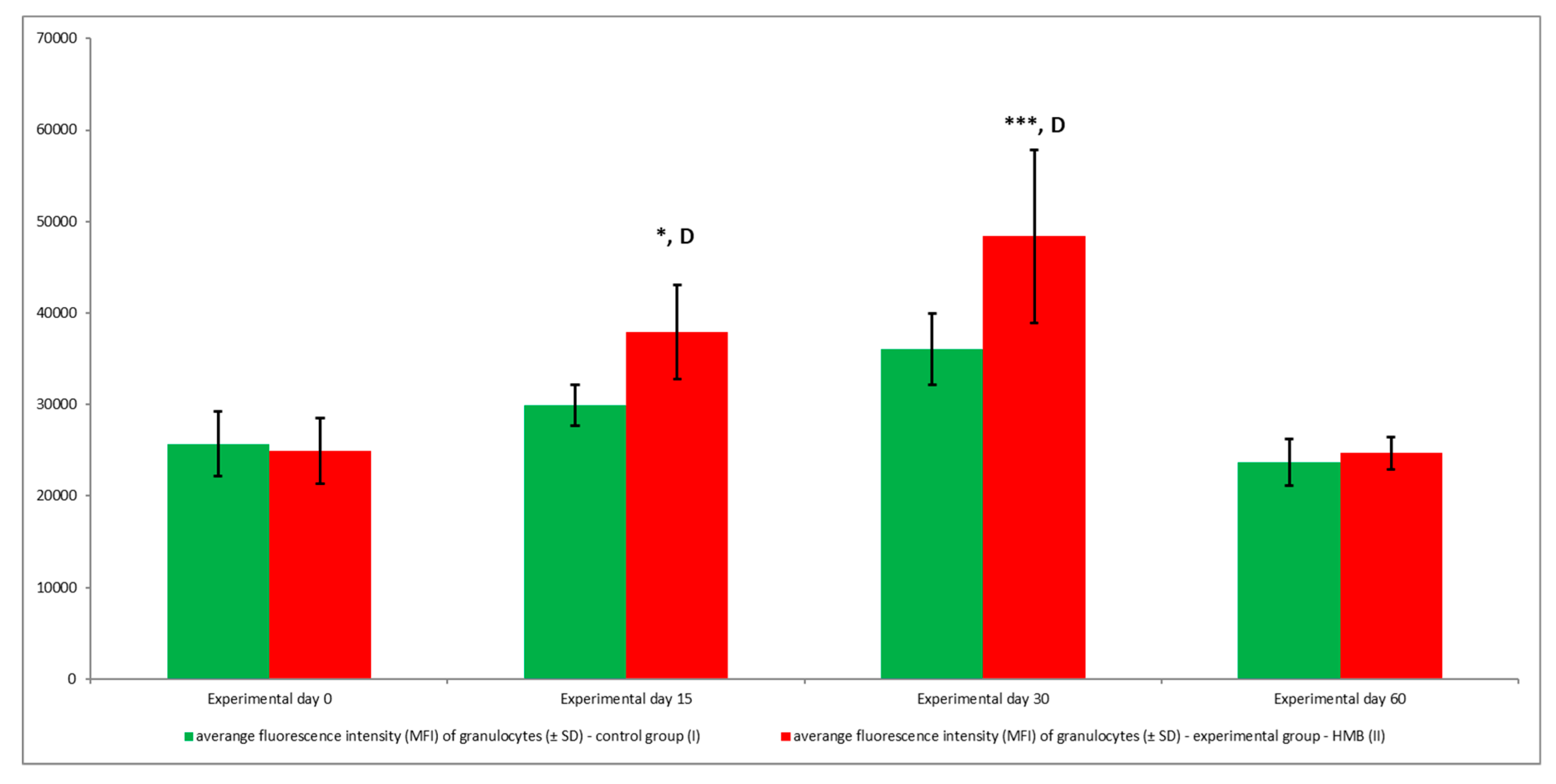
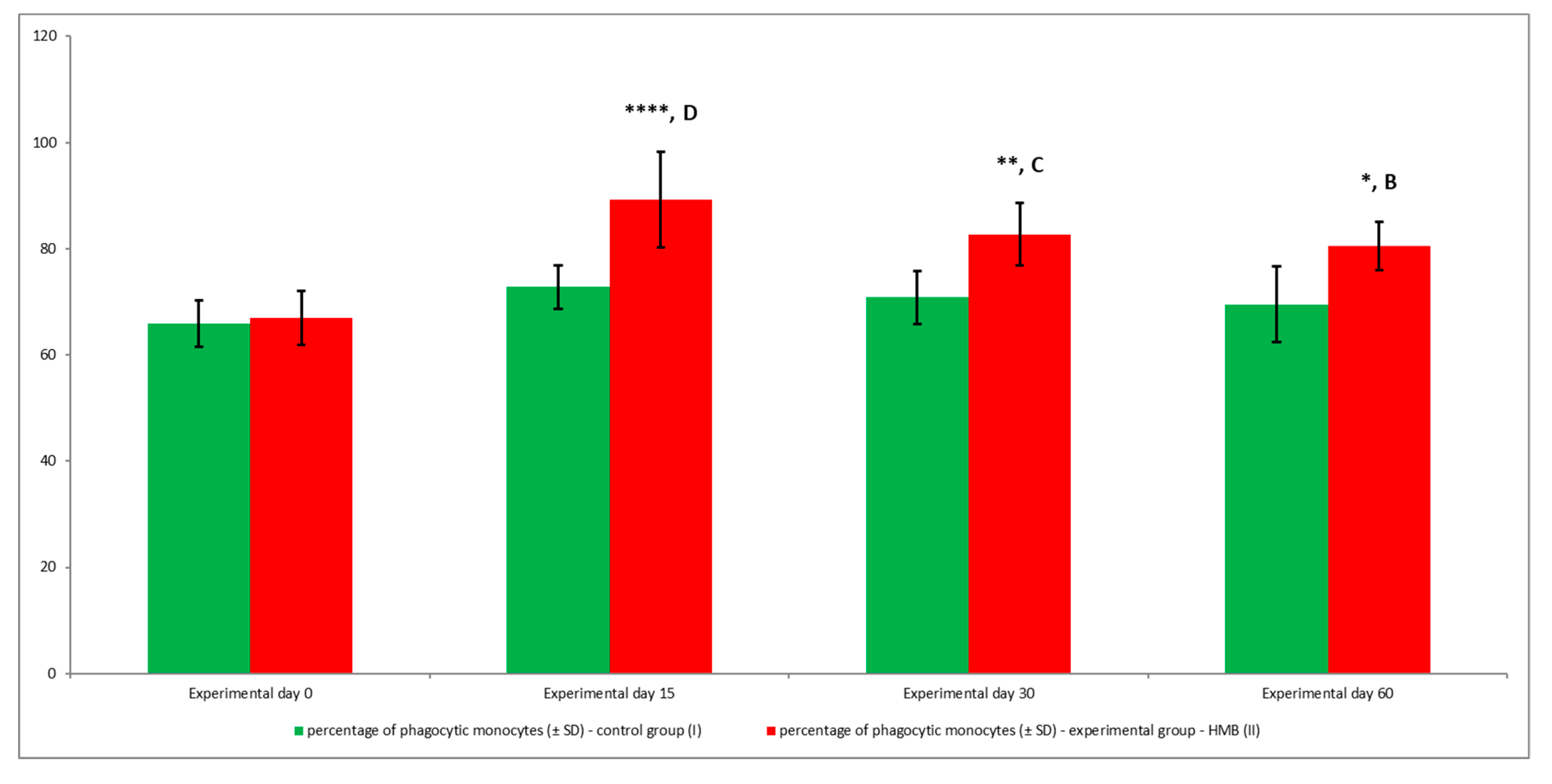
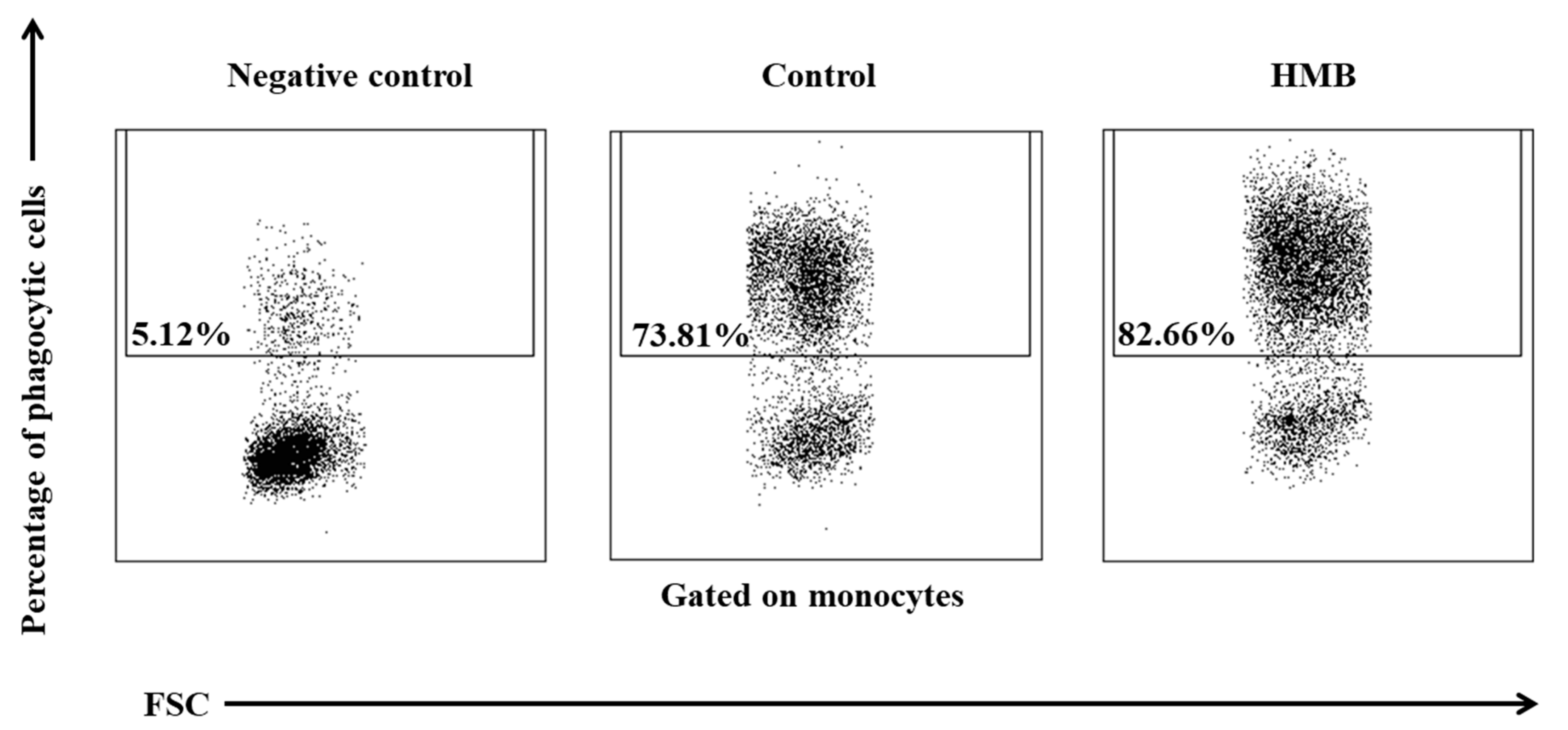
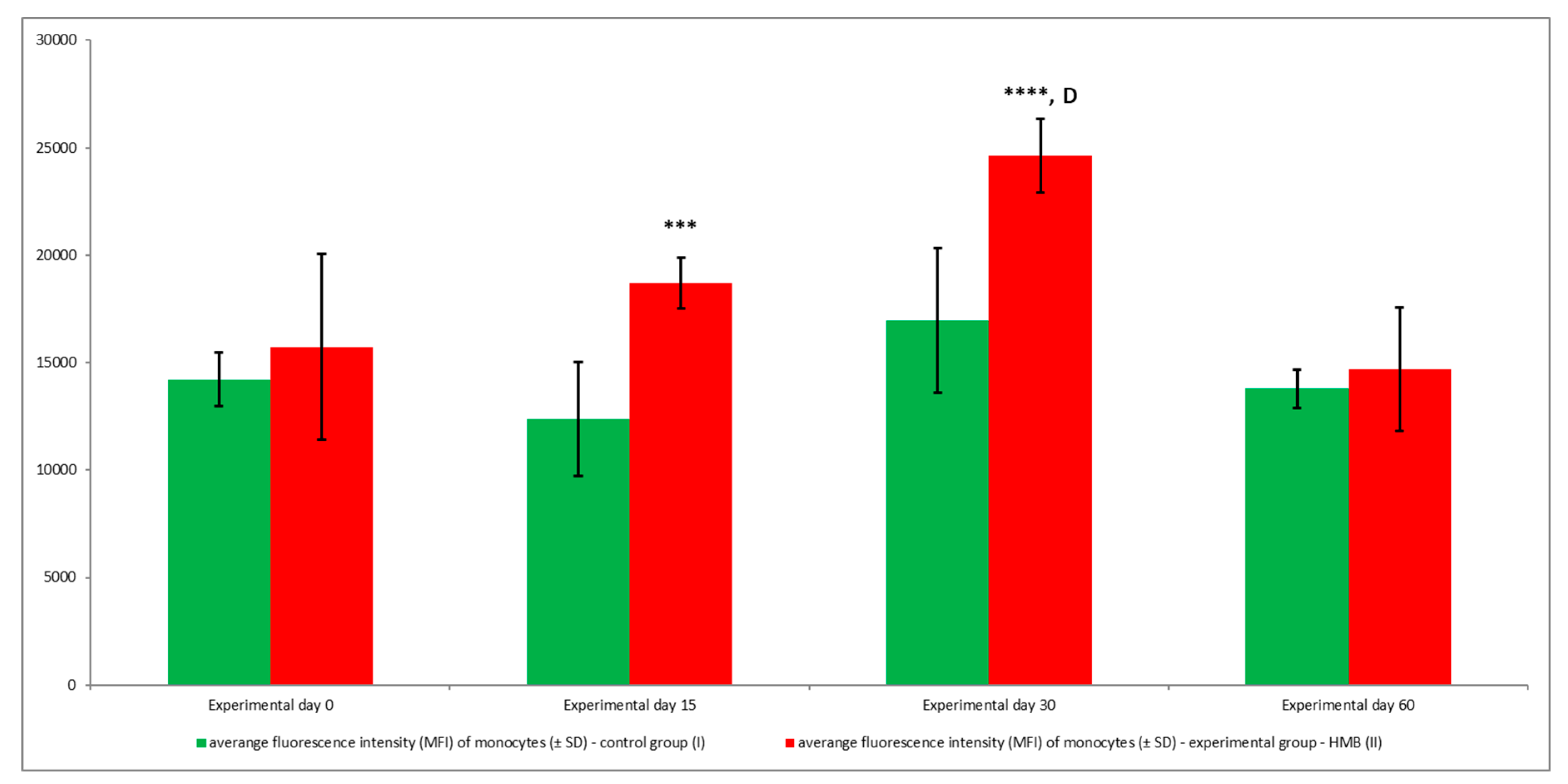
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bielecka, M.; Smoragiewicz, W.; Siwicki, A.K.; Wójcik, R.; Biedrzycka, E.; Orłowski, A.; Kask, S.; Jankowski, J.; Karska-Wysocki, B.; Ham, D. The effect of various probiotic strains or Avilamycin feed additive on immune defense markers and acute-phase response to Salmonella infection in chickens. Probiotics Antimicrob. Prot. 2010, 2, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Wójcik, R. Effect of Biolex-MB40 on phagocytic activity and oxidative metabolism of peripheral blood granulocytes and monocytes in lambs. Pol. J. Vet. Sci. 2014, 17, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Arai, S. Recent trends in functional food science and the industry in Japan. Biosci. Biotechnol. Biochem. 2002, 66, 2017–2029. [Google Scholar] [CrossRef]
- Małaczewska, J.; Wójcik, R.; Jung, L.; Siwicki, A.K. The effect of Biolex Beta-HP on the selected parameters of biochemical, specific and non-specific humoral and cellular immunity in rats. Bull. Vet. Inst. Pulawy 2010, 54, 75–80. [Google Scholar]
- Siwicki, A.K.; Małaczewska, J.; Kazuń, B.; Wójcik, R. Immunomodulating effect of methisoprinol on the pronephros macrophage and lymphocyte activity after suppression induced by infectious haematopoietic necrosis virus (IHNV) in rainbow trout (Oncorhynchus mykiss). Acta Vet. Brno 2008, 77, 631–635. [Google Scholar] [CrossRef]
- Święcicka-Grabowska, G.; Wójcik, R. Immunity against viruses in chickens after applying natural and synthetic immunity stimulator. Pol. J. Vet. Sci. 2003, 6, 60–63. [Google Scholar]
- Wójcik, R.; Milewski, S.; Małaczewska, J.; Tański, Z.; Brzostowski, H.; Siwicki, A.K. Defence mechanisms of the offspring of ewes fed a diet supplemented with yeast (Saccharomyces cerevisiae) during pregnancy and lactation. Centr. Eur. J. Immunol. 2008, 33, 197–201. [Google Scholar]
- Ząbek, K.; Milewski, S.; Wójcik, R.; Siwicki, A.K. Effect of β-1,3/1,6-D-glucan in diet on productivity and humoral and cellular defence mechanisms in sheep. Acta Vet. Brno 2013, 82, 141–146. [Google Scholar] [CrossRef]
- Nissen, S.L.; Abumrad, N.N. Nutritional role of the leucine metabolite β-hydroxy β-methylbutyrate (HMB). Nutr. Biochem. 1997, 8, 300–311. [Google Scholar] [CrossRef]
- Van Koevering, M.; Nissen, S. Oxidation of leucine and α-ketoisocaproate to β-hydroxy-β-methylbutyrate in vivo. Am. J. Physiol. 1992, 262, E27–E31. [Google Scholar] [CrossRef]
- Wilson, J.M.; Grant, S.C.; Lee, S.R. Beta-hydroxy-beta-methylbutyrate blunts negative age-related changes in body composition, functionality and myofiber dimensions in rats. J. Int. Soc. Sports Nutr. 2012, 9, 18. [Google Scholar] [CrossRef]
- Tatara, R. Neonatal programming of skeletal development in sheep is mediated by somatotrophic axis function. Exp. Physiol. 2008, 93, 763–772. [Google Scholar] [CrossRef]
- Qiao, X.; Zhang, H.J.; Wu, S.G.; Yue, H.Y.; Zuo, J.J.; Feng, D.Y.; Qi, G.H. Effect of β-hydroxy-β-methylbutyrate calcium on growth, blood parameters, and carcass qualities of broiler chickens. Poult. Sci. 2013, 92, 753–759. [Google Scholar] [CrossRef]
- Arazi, H.; Taati, B.; Suzuki, K. A review of the effects of leucine metabolite (β-hydroxy-β-methylbutyrate) supplementation and resistance training on inflammatory markers: A new approach to oxidative stress and cardiovascular risk factors. Antioxidants (Basel) 2018, 7, 148. [Google Scholar] [CrossRef]
- Gepner, Y.; Varanoske, A.N.; Boffey, D.; Hoffman, J.R. Benefits of β-hydroxy-β-methylbutyrate supplementation in trained and untrained individuals. Res. Sports Med. 2019, 27, 204–218. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Hirayama, K.; Ueda, H.; Ochi, E. Two and four weeks of β-hydroxy-β-methylbutyrate (HMB) supplementations reduce muscle damage following eccentric contractions. J. Am. Coll. Nutr. 2019, 38, 373–379. [Google Scholar]
- Estévez, R.A.; Mostazo, M.G.C.; Rodriguez, E.; Espinoza, J.C.; Kuznar, J.; Jónsson, Z.O.; Guðmundsson, G.H.; Maier, V.H. Inducers of salmon innate immunity: An in vitro and in vivo approach. Fish Shellfish Immunol. 2018, 72, 247–258. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Morand, M.; Fuller, J.C., Jr.; Nissen, S.; Goryczko, K.; Ostaszewski, P.; Kazuń, K.; Głombski, E. Influence of feeding the leucine metabolite β-hydroxy-β-methylbutyrate (HMB) on the non-specific cellular and humoral defence mechanisms of rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2003, 19, 44–48. [Google Scholar] [CrossRef]
- Puchajda-Skowrońska, H.; Siwicki, A.K.; Wójcik, R.; Pudyszak, K. Effects of 3-hydroxy-3-methylbutyrate (HMB) on selected performance indices and non-specific defense mechanisms in geese. Medycyna Wet. 2006, 62, 89–92. [Google Scholar]
- Krakowski, L.; Krzyżanowski, J.; Wrona, Z.; Kostro, K.; Siwicki, A.K. The influence of nonspecific immunostimulation of pregnant sows on the immunological value of colostrum. Vet. Immunol. Immunopathol. 2002, 87, 89–95. [Google Scholar] [CrossRef]
- Ostaszewski, P.; Kozłowska, E.; Siwicki, A.K.; Krzyżanowski, J.; Fuller, J.C., Jr.; Nissen, S. The immunomodulating activity of dietary β-hydroxy-β-methylbutyrate (HMB) in weanling pigs. J. Anim. Sci. 1998, 76 (Suppl. 1), 136. [Google Scholar]
- Wójcik, R.; Małaczewska, J.; Siwicki, A.K.; Miciński, J.; Zwierzchowski, G. The effect of beta-hydroxy-beta-methylbutyrate (HMB) on the proliferative response of blood lymphocytes and the phagocytic activity of blood monocytes and granulocytes in calves. Pol. J. Vet. Sci. 2013, 16, 567–569. [Google Scholar] [CrossRef]
- Wójcik, R.; Małaczewska, J.; Siwicki, A.K.; Miciński, J.; Zwierzchowski, G. The effect of β-hydroxy-β-methylbutyrate (HMB) on selected parameters of humoral immunity in calves. Pol. J. Vet. Sci. 2014, 17, 357–359. [Google Scholar] [CrossRef]
- Nissen, S.; Morrical, D.; Fuller, J.C., Jr. The effects of the leucine catabolite ß-hydroxy-ß-methylbutyrate on the growth and health of growing lambs. J. Anim. Sci. 1994, 77 (Suppl. 1), 243. [Google Scholar]
- Ząbek, K.; Wójcik, R.; Milewski, S.; Małaczewska, J.; Tański, Z.; Siwicki, A.K. Effect of β-hydroxy-β-methylbutyrate acid on meat performance traits and selected indicators of humoral immunity in goats. Jpn. J. Vet. Res. 2016, 64, 247–256. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Innate immunity. In Basic Immunology. Functions and Disorders of the Immune System, 4th ed.; Abbas, A.K., Lichtman, A.H., Pillai, S., Eds.; Elsevier Inc.: Philadelphia, PA, USA, 2014; pp. 25–52. [Google Scholar]
- Kuhlman, G.; Roth, J.A.; Flakoll, P.J.; Vahdehaar, M.J.; Nissen, S. Effects of dietary leucine, α-ketoisocaproate and isovalerate on antibody production and lymphocyte blastogenesis in growing lambs. J. Nutr. 1988, 118, 1564–1569. [Google Scholar] [CrossRef]
- Guilhem, A.; Malcus, C.; Clarivet, B.; Plauchu, H.; Dupuis-Girod, S. Immunological abnormalities associated with hereditary haemorrhagic telangiectasia. J. Intern. Med. 2013, 274, 351–362. [Google Scholar] [CrossRef]
- Ross, G.D.; Cain, J.A.; Myones, B.L.; Newman, S.L.; Lachmann, P.J. Specificity of membrane complement receptor type three (CR3) for beta-glucans. Complement 1987, 4, 61–74. [Google Scholar] [CrossRef]
- Zhou, M.J.; Brown, E.J. CR3 (Mac-1, alpha M beta 2, CD11b/CD18) and Fc gamma RIII cooperate in generation of a neutrophil respiratory burst: Requirement for Fc gamma RIII and tyrosine phosphorylation. J. Cell Biol. 1994, 125, 1407–1416. [Google Scholar] [CrossRef]
- Underhill, D.M.; Ozinsky, A. Phagocytosis of microbes: Complexity in action. Ann. Rev. Immunol. 2002, 20, 825–852. [Google Scholar] [CrossRef]
- Rice, P.J.; Kelley, J.L.; Kogan, G.; Ensley, H.E.; Kalbfleisch, J.H.; Browder, I.W.; Williams, D.L. Human monocyte scavenger receptors are pattern recognition receptors for (1→3)-β-D-glucans. J. Leukoc. Biol. 2002, 72, 140–146. [Google Scholar]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Freeman, S.A.; Grinstein, S. Phagocytosis: Receptors, signal integration, and the cytoskeleton. Immunol. Rev. 2014, 262, 193–215. [Google Scholar] [CrossRef]
- Gordon, S. Phagocytosis: An Immunobiologic Process. Immunity 2016, 44, 463–475. [Google Scholar] [CrossRef]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological defense mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investg. 1973, 52, 741–744. [Google Scholar] [CrossRef]
- Babior, B.M. NADPH oxidase: An update. Blood 1999, 93, 1464–1476. [Google Scholar] [CrossRef]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Ann. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef]
- Faurschou, M.; Borregaard, N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect. 2003, 5, 1317–1327. [Google Scholar] [CrossRef]
- Ostaszewski, P.; Kostiuk, S.; Balasinska, B.; Jank, M.; Papet, I.; Glomot, F. The leucine metabolite 3-hydroxy-3-methylbutyrate (HMB) modifies protein turnover in muscles of laboratory rats and domestic chickens in vitro. J. Anim. Physiol. Anim. Nutr. 2000, 84, 1–8. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Hossain, T.; Hill, D.S.; Phillips, B.E.; Crossland, H.; Williams, J.; Loughna, P.; Churchward-Venne, T.A.; Breen, L.; Phillips, S.M.; et al. Effects of leucine and its metabolite beta-hydroxy-beta-methylbutyrate on human skeletal muscle protein metabolism. J. Physiol. 2013, 591, 2911–2923. [Google Scholar] [CrossRef]
- Van Koevering, M.T.; Dolezal, H.G.; Gill, D.R.; Owens, F.N.; Strasia, C.A.; Buchanan, D.S.; Lake, R.; Nissen, S. Effects of beta-hydroxy-beta-methylbutyrate on performance and carcass quality of meat producing feedlot steers. J. Anim. Sci. 1994, 72, 1927–1935. [Google Scholar] [CrossRef]
- Nissen, S.; Fuller, J.C., Jr.; Sell, J.; Ferket, P.R.; Rives, D.V. The effect of beta-hydroxy-beta-methylbutyrate on growth, mortality, and carcass qualities of broiler chickens. Poult. Sci. 1994, 73, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.T.; Ferket, P.R.; Mozdziak, P.E. The effect of early nutrition on satellite cell dynamics in the young turkey. Poult. Sci. 2005, 84, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.R.; Fragala, M.S.; Jajtner, A.R.; Gonzalez, A.M.; Wells, A.J.; Mangine, G.T.; Robinson, E.H., 4th; McCormack, W.P.; Beyer, K.S.; Pruna, G.J.; et al. β-Hydroxy-β-methylbutyrate (HMB)-free acid attenuates circulating TNF-α and TNFR1 expression post resistance exercise. J. Appl. Physiol. 2013, 115, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, S.; Ogo, A.; Kubota, H.; Teramoto, F.; Hirai, T. β-Hydroxy-β-methylbutyrate Suppresses NF-ĸB Activation and IL-6 Production in TE-1 Cancer Cells. In Vivo 2019, 33, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.R.; Gepner, Y.; Stout, J.R.; Hoffman, M.W.; Ben-Dov, D.; Funk, S.; Daimont, I.; Jajtner, A.R.; Townsend, J.R.; Church, D.D. β-Hydroxy-β-methylbutyrate attenuates cytokine response during sustained military training. Nutr. Res. 2016, 36, 553–563. [Google Scholar] [CrossRef]
- Gonzalez, A.M.; Fragala, M.S.; Jajtner, A.R.; Townsend, J.R.; Wells, A.J.; Beyer, K.S.; Boone, C.H.; Pruna, G.J.; Mangine, G.T.; Bohner, J.D.; et al. Effects of β-hydroxy-β-methylbutyrate free acid and cold water immersion on expression of CR3 and MIP-1β following resistance exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Józefowski, S.; Yang, Z.; Marcinkiewicz, J.; Kobzik, L. Scavenger receptors and β-glucan receptors participate in the recognition of yeasts by murine macrophages. Inflamm. Res. 2012, 61, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Bowdish, D.M.E.; Loffredo, M.S.; Mukhopadhyay, S.; Mantovani, A.; Gordon, S. Macrophage receptors implicated in the “adaptive” form of innate immunity. Microbes Infect. 2007, 9, 1680–1687. [Google Scholar] [CrossRef]
- Dunne, D.W.; Resnick, D.; Greenberg, J.; Krieger, M.; Joiner, K.A. The type I macrophage scavenger receptor binds to gram-positive bacteria and recognizes lipoteichoic acid. Proc. Natl. Acad. Sci. USA 1994, 91, 1863–1867. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; de Beer, M.C.; de Beer, F.C.; van der Westhuyzen, D.R. Serum amyloid A is a ligand for scavenger receptor class B type I and inhibits high density lipoprotein binding and selective lipid uptake. J. Biol. Chem. 2005, 280, 2954–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jozefowski, S.; Sulahian, T.; Arredouani, M.; Kobzik, L. Role of scavenger receptor MARCO in macrophage responses to CpG oligodeoxynucleotides. J. Leukoc. Biol. 2006, 80, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Peiser, L.; De Winther, M.P.; Makepeace, K.; Hollinshead, M.; Coull, P.; Plested, J.; Kodama, T.; Moxon, E.R.; Gordon, S. The class A macrophage scavenger receptor is a major pattern recognition receptor for Neisseria meningitidis which is independent of lipopolysaccharide and not required for secretory responses. Infect. Immun. 2002, 70, 5346–5354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Peiser, L.; Gordon, S. Activation of murine macrophages by Neisseria meningitidis and IFN-gamma in vitro: Distinct roles of class A scavenger and Toll-like pattern recognition receptors in selective modulation of surface phenotype. J. Leukoc. Biol. 2004, 76, 577–584. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Chen, Y.; Sankala, M.; Peiser, L.; Pikkarainen, T.; Kraal, G.; Tryggvason, K.; Gordon, S. MARCO, an innate activation marker of macrophages, is a class A scavenger receptor for Neisseria meningitidis. Eur. J. Immunol. 2006, 36, 940–949. [Google Scholar] [CrossRef]
- Peterson, A.L.; Qureshi, M.A.; Ferket, P.R.; Fuller, J.C., Jr. In vitro exposure with β-hydroxy-β-methylbutyrate enhances chicken macrophage growth and function. Vet. Immunol. Immunopathol. 1999, 67, 67–78. [Google Scholar] [CrossRef]
- Peterson, A.L.; Qureshi, M.A.; Ferket, P.R.; Fuller, J.C. Enhancement of cellular and humoral immunity in young broilers by the dietary supplementation of beta-hydroxy-beta-methylbutyrate. Poult. Sci. 1999, 21, 307–330. [Google Scholar]
- Kunttu, H.M.; Valtonen, E.T.; Suomalainen, L.R.; Vielma, J.; Jokinen, I.E. The efficacy of two immunostimulants against Flavobacterium columnare infection in juvenile rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2009, 26, 850–857. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Fuller, J.C., Jr.; Nissen, S.; Ostaszewski, P.; Studnicka, M. In vitro effects of beta-hydroxy-beta-methylbutyrate (HMB) on cell-mediated immunity in fish. Vet. Immunol. Immunopathol. 2000, 76, 191–197. [Google Scholar] [CrossRef]
















© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójcik, R.; Ząbek, K.; Małaczewska, J.; Milewski, S.; Kaczorek-Łukowska, E. The Effects of β-Hydroxy-β-Methylbutyrate (HMB) on Chemotaxis, Phagocytosis, and Oxidative Burst of Peripheral Blood Granulocytes and Monocytes in Goats. Animals 2019, 9, 1031. https://doi.org/10.3390/ani9121031
Wójcik R, Ząbek K, Małaczewska J, Milewski S, Kaczorek-Łukowska E. The Effects of β-Hydroxy-β-Methylbutyrate (HMB) on Chemotaxis, Phagocytosis, and Oxidative Burst of Peripheral Blood Granulocytes and Monocytes in Goats. Animals. 2019; 9(12):1031. https://doi.org/10.3390/ani9121031
Chicago/Turabian StyleWójcik, Roman, Katarzyna Ząbek, Joanna Małaczewska, Stanisław Milewski, and Edyta Kaczorek-Łukowska. 2019. "The Effects of β-Hydroxy-β-Methylbutyrate (HMB) on Chemotaxis, Phagocytosis, and Oxidative Burst of Peripheral Blood Granulocytes and Monocytes in Goats" Animals 9, no. 12: 1031. https://doi.org/10.3390/ani9121031
APA StyleWójcik, R., Ząbek, K., Małaczewska, J., Milewski, S., & Kaczorek-Łukowska, E. (2019). The Effects of β-Hydroxy-β-Methylbutyrate (HMB) on Chemotaxis, Phagocytosis, and Oxidative Burst of Peripheral Blood Granulocytes and Monocytes in Goats. Animals, 9(12), 1031. https://doi.org/10.3390/ani9121031