The Genus Allium as Poultry Feed Additive: A Review

Abstract
:Simple Summary
Abstract
1. Introduction
2. Overview of Major Bioactive Compounds in Allium
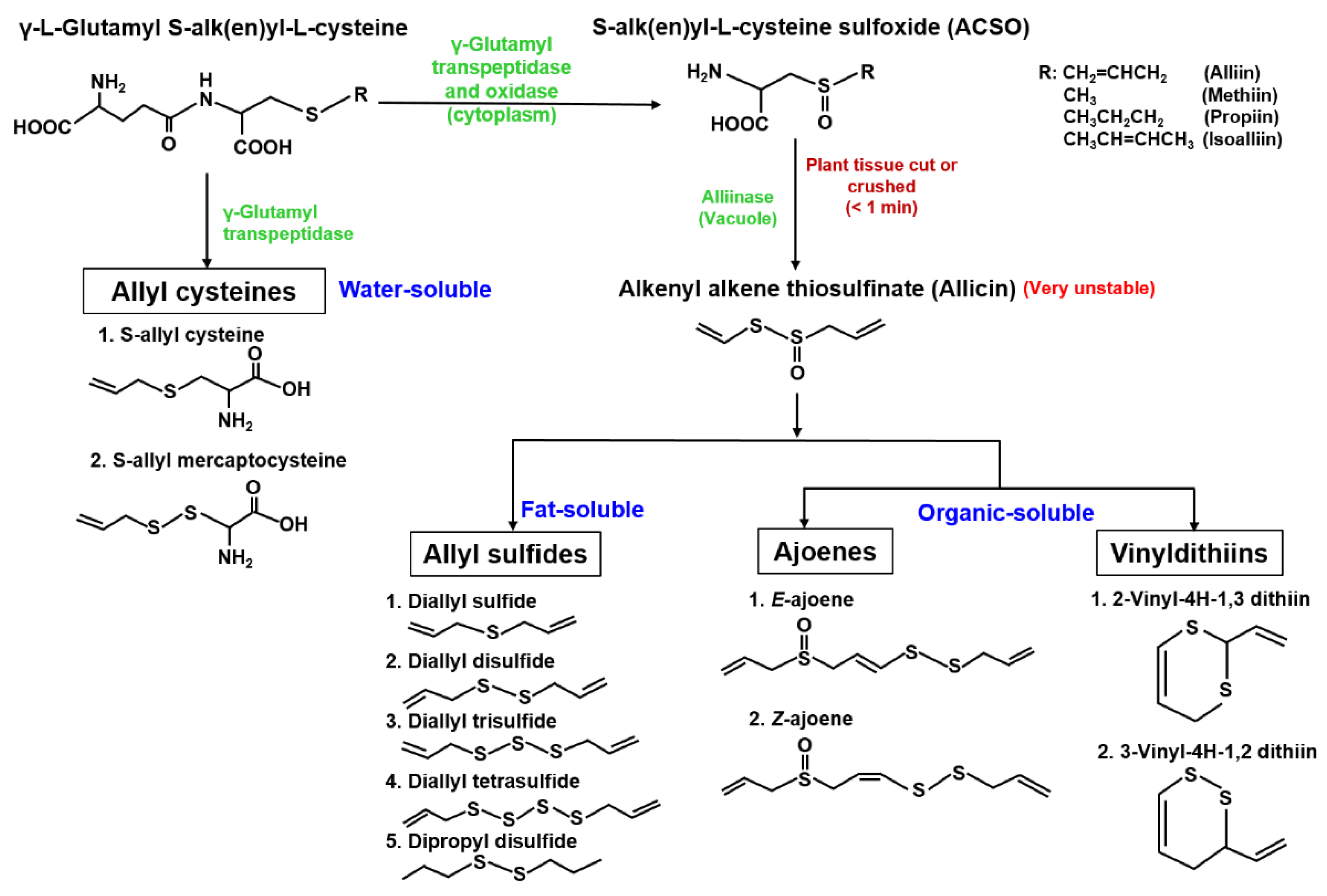
2.1. Organosulfur Compounds
2.2. Polyphenolic Compounds
2.3. Saponins
2.4. Fructans and Fructo-Oligosaccharides
3. Allium spp. as Poultry Feed Additives
3.1. Effects on Growth Performance
3.2. Hypolipidemic and Hypocholesterolemic Effects
3.3. Effects on Infectious Diseases
3.4. Effects on Intestinal Microbiota and Morphology
3.5. Effects on Immune Response
3.6. Effects on Product Quality
4. Factors Determining the Effectiveness of Alliums in Poultry Feed
5. Fortification/Preservation of Allium Bioactivity
5.1. Fermentation
5.2. Microencapsulation/Nanotechnology
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Roth, N.; Hofacre, C.; Zitz, U.; Mathis, G.F.; Moder, K.; Doupovec, B.; Berghouse, R.; Domig, K.J. Prevalence of antibiotic-resistant E. coli in broilers challenged with a multi-resistant E. coli strain and received ampicillin, an organic acid-based feed additive or a synbiotic preparation. Poult. Sci. 2019, 98, 2598–2607. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Das, R.K.; Brar, S.K.; Rouissi, T.; Ramirez, A.A.; Chorfi, Y.; Godbout, S. Alternatives to antibiotics in poultry feed: Molecular perspectives. Crit. Rev. Microbiol. 2018, 44, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- Diarra, M.S.; Malouin, F. Antibiotics in Canadian poultry productions and anticipated alternatives. Front. Microbiol. 2014, 5, 282. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Létourneau-Montminy, M.-P.; Gaucher, M.-L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Côté, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef]
- Kim, W.H.; Lillehoj, H.S. Immunity, immunomodulation, and antibiotic alternatives to maximize the genetic potential of poultry for growth and disease response. Anim. Feed Sci. Technol. 2019, 250, 41–50. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef]
- Cervantes, H.M. Antibiotic-free poultry production: Is it sustainable? J. Appl. Poult. Res. 2015, 24, 91–97. [Google Scholar] [CrossRef]
- Lagha, A.B.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial potential of bacteriocins in poultry and swine production. Vet. Res. 2017, 48, 22. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.; Oh, S.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Habibi, H.; Ghahtan, N. Effects of medicinal plant powder as feed additives on growth performance, carcass characteristics, and immune response of Coturnix japonica against avian influenza and Newcastle disease vaccine virus. Comp. Clin. Pathol. 2019, 28, 1581–1588. [Google Scholar] [CrossRef]
- Sugiharto, S. Role of nutraceuticals in gut health and growth performance of poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111. [Google Scholar] [CrossRef]
- Toghyani, M.; Toghyani, M.; Gheisari, A.; Ghalamkari, G.; Eghbalsaied, S. Evaluation of cinnamon and garlic as antibiotic growth promoter substitutions on performance, immune responses, serum biochemical and haematological parameters in broiler chicks. Livest. Sci. 2011, 138, 167–173. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Mnayer, D.; Tabanelli, G.; Stojanović-Radić, Z.; Sharifi-Rad, M.; Yousaf, Z.; Vallone, L.; Setzer, W.; Iriti, M. Plants of the genus Allium as antibacterial agents: From tradition to pharmacy. Cell. Mol. Biol. 2016, 62, 57–68. [Google Scholar]
- Poojary, M.M.; Putnik, P.; Kovačević, D.B.; Barba, F.J.; Lorenzo, J.M.; Dias, D.A.; Shpigelman, A. Stability and extraction of bioactive sulfur compounds from Allium genus processed by traditional and innovative technologies. J. Food Compos. Anal. 2017, 61, 28–39. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Locatelli, D.A.; Gonzalez, R.E.; Cavagnaro, P.F.; Camargo, A.B. Analytical methods for bioactive sulfur compounds in Allium: An integrated review and future directions. J. Food Compos. Anal. 2017, 61, 4–19. [Google Scholar] [CrossRef]
- Omar, S.; Al-Wabel, N. Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharm. J. 2010, 18, 51–58. [Google Scholar] [CrossRef]
- Chope, G.A.; Cools, K.; Terry, L.A. Alliums (Onion, Garlic, Leek and Shallot). Health-Promot. Prop. Fruits Veg. 2011, 1, 5–28. [Google Scholar]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Kovačević, D.B. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef]
- Kyung, K.H. Antimicrobial properties of allium species. Curr. Opin. Biotechnol. 2012, 23, 142–147. [Google Scholar] [CrossRef]
- Lanzotti, V.; Bonanomi, G.; Scala, F. What makes Allium species effective against pathogenic microbes? Phytochem. Rev. 2013, 12, 751–772. [Google Scholar] [CrossRef]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef] [PubMed]
- Snoussi, M.; Trabelsi, N.; Dehmeni, A.; Benzekri, R.; Bouslama, L.; Hajlaoui, B.; Al-sieni, A.; Papetti, A. Phytochemical analysis, antimicrobial and antioxidant activities of Allium roseum var. odoratissimum (Desf.) Coss extracts. Ind. Crops Prod. 2016, 89, 533–542. [Google Scholar] [CrossRef]
- Li, R.; Wang, Y.-F.; Sun, Q.; Hu, H.-B. Chemical composition and antimicrobial activity of the essential oil from Allium hookeri consumed in Xishuangbanna, southwest China. Nat. Prod. Commun. 2014, 9, 863–864. [Google Scholar]
- Mihaylova, D.; Popova, A. Antimicrobial and antioxidant activity of extracts of Allium ursinum L. J. BioSci. Biotech. 2012, 143–145. [Google Scholar]
- Salem, W.; El-Hamed, D.S.; Sayed, W.; Elamary, R. Alterations in virulence and antibiotic resistant genes of multidrug-resistant Salmonella serovars isolated from poultry: The bactericidal efficacy of Allium sativum. Microb. Pathog. 2017, 108, 91–100. [Google Scholar] [CrossRef]
- Velkers, F.; Dieho, K.; Pecher, F.; Vernooij, J.; Van Eck, J.; Landman, W. Efficacy of allicin from garlic against Ascaridia galli infection in chickens. Poult. Sci. 2011, 90, 364–368. [Google Scholar] [CrossRef]
- Anthony, J.-P.; Fyfe, L.; Smith, H. Plant active components—A resource for antiparasitic agents? Trends Parasitol. 2005, 21, 462–468. [Google Scholar] [CrossRef]
- Tsao, S.-M.; Yin, M.-C. In-Vitro antimicrobial activity of four diallyl sulphides occurring naturally in garlic and Chinese leek oils. J. Med. Microbiol. 2001, 50, 646–649. [Google Scholar] [CrossRef]
- Galdón, B.R.; Rodríguez, E.M.R.; Romero, C.D. Flavonoids in onion cultivars (Allium cepa L.). J. Food Sci. 2008, 73, C599–C605. [Google Scholar] [CrossRef] [PubMed]
- Maisashvili, M.; Gvazava, L.; Kuchukhidze, J. Flavonoids and coumarins from Allium rotundum. Chem. Nat. Compd. 2009, 45, 87–88. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Wu, H.; Dushenkov, S.; Ho, C.-T.; Sang, S. Novel acetylated flavonoid glycosides from the leaves of Allium ursinum. Food Chem. 2009, 115, 592–595. [Google Scholar] [CrossRef]
- Leighton, T.; Ginther, C.; Fluss, L.; Harter, W.K.; Cansado, J.; Notario, V. Molecular characterization of quercetin and quercetin glycosides in Allium vegetables: Their effects on malignant cell transformation. Proc. ACS Symp. Ser. 1992, 507, 220. [Google Scholar]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [Green Version]
- Parvu, M.; Toiu, A.; Vlase, L.; Parvu, E.A. Determination of some polyphenolic compounds from Allium species by HPLC-UV-MS. Nat. Prod. Res. 2010, 24, 1318–1324. [Google Scholar] [CrossRef]
- Li, M.; Zhu, X.; Tian, J.; Liu, M.; Wang, G. Dietary flavonoids from Allium mongolicum Regel promotes growth, improves immune, antioxidant status, immune-related signaling molecules and disease resistance in juvenile northern snakehead fish (Channa argus). Aquaculture 2019, 501, 473–481. [Google Scholar] [CrossRef]
- Qi, S.; Wang, T.; Chen, R.; Wang, C.; Ao, C. Effects of flavonoids from Allium mongolicum Regel on growth performance and growth-related hormones in meat sheep. Anim. Nutr. 2017, 3, 33–38. [Google Scholar]
- Sobolewska, D.; Michalska, K.; Podolak, I.; Grabowska, K. Steroidal saponins from the genus Allium. Phytochem. Rev. 2016, 15, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostafa, A.; Sudisha, J.; El-Sayed, M.; Ito, S.-I.; Ikeda, T.; Yamauchi, N.; Shigyo, M. Aginoside saponin, a potent antifungal compound, and secondary metabolite analyses from Allium nigrum L. Phytochem. Lett. 2013, 6, 274–280. [Google Scholar] [CrossRef]
- Lanzotti, V.; Scala, F.; Bonanomi, G. Compounds from Allium species with cytotoxic and antimicrobial activity. Phytochem. Rev. 2014, 13, 769–791. [Google Scholar] [CrossRef]
- Ernst, M.K.; Chatterton, N.J.; Harrison, P.A.; Matitschka, G. Characterization of fructan oligomers from species of the genus Allium L. J. Plant Physiol. 1998, 153, 53–60. [Google Scholar] [CrossRef]
- Chandrashekar, P.M.; Prashanth, K.V.H.; Venkatesh, Y.P. Isolation, structural elucidation and immunomodulatory activity of fructans from aged garlic extract. Phytochemistry 2011, 72, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-B.; Miyake, S.; Umetsu, R.; Hayashi, K.; Chijimatsu, T.; Hayashi, T. Anti-influenza A virus effects of fructan from Welsh onion (Allium fistulosum L.). Food Chem. 2012, 134, 2164–2168. [Google Scholar] [CrossRef]
- Li, M.; Yan, Y.X.; Yu, Q.T.; Deng, Y.; Wu, D.T.; Wang, Y.; Ge, Y.Z.; Li, S.P.; Zhao, J. Comparison of immunomodulatory effects of fresh garlic and black garlic polysaccharides on RAW 264.7 macrophages. J. Food Sci. 2017, 82, 765–771. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, X.; Zeng, Y.; Wu, X.; Peng, X. Study on prebiotic effectiveness of neutral garlic fructan in vitro. Food Sci. Hum. Wellness 2013, 2, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Lee, S.; Gadde, U.D.; Oh, S.; Lee, S.; Lillehoj, H.S. Dietary Allium hookeri reduces inflammatory response and increases expression of intestinal tight junction proteins in LPS-induced young broiler chicken. Res. Vet. Sci. 2017, 112, 149–155. [Google Scholar] [CrossRef]
- Malafaia, C.R.; da Silva, B.P.; Tinoco, L.W.; Parente, J.P. Structural characterization and gastroprotective property of a novel glucofructan from Allium ampeloprasum var. porrum. Carbohydr. Res. 2015, 402, 44–49. [Google Scholar] [CrossRef]
- Kumar, V.P.; Prashanth, K.H.; Venkatesh, Y. Structural analyses and immunomodulatory properties of fructo-oligosaccharides from onion (Allium cepa). Carbohydr. Polym. 2015, 117, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Farahani, M.; Goodarzi, M.; Nanekarani, S. The effects of aqueous extract of onion on performance and some blood biochemical parameters of the Cobb and Ross broilers. Int. J. Adv. Biol. Biomed. Res. 2015, 3, 370–377. [Google Scholar]
- Ur Rahman, S.; Khan, S.; Chand, N.; Sadique, U.; Khan, R.U. In vivo effects of Allium cepa L. on the selected gut microflora and intestinal histomorphology in broiler. Acta Histochem. 2017, 119, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.; Landy, N.; Nanekarani, S. Effect of onion (Allium cepa L.) as an antibiotic growth promoter substitution on performance, immune responses and serum biochemical parameters in broiler chicks. Health 2013, 5, 1210. [Google Scholar] [CrossRef] [Green Version]
- Brzóska, F.; Śliwiński, B.; Michalik-Rutkowska, O.; Śliwa, J. The effect of garlic (Allium sativum L.) on growth performance, mortality rate, meat and blood parameters in broilers. Ann. Anim. Sci. 2015, 15, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Sheoran, N.; Kumar, R.; Kumar, A.; Batra, K.; Sihag, S.; Maan, S.; Maan, N. Nutrigenomic evaluation of garlic (Allium sativum) and holy basil (Ocimum sanctum) leaf powder supplementation on growth performance and immune characteristics in broilers. Vet. World 2017, 10, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Kirubakaran, A.; Moorthy, M.; Chitra, R.; Prabakar, G. Influence of combinations of fenugreek, garlic, and black pepper powder on production traits of the broilers. Vet. World 2016, 9, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Varmaghany, S.; Torshizi, M.A.K.; Rahimi, S.; Lotfollahian, H.; Hassanzadeh, M. The effects of increasing levels of dietary garlic bulb on growth performance, systolic blood pressure, hematology, and ascites syndrome in broiler chickens. Poult. Sci. 2015, 94, 1812–1820. [Google Scholar] [CrossRef]
- Pourali, M.; Mirghelenj, S.A.; Kermanshahi, H. Effects of garlic powder on productive performance and immune response of broiler chickens challenged with Newcastle Disease Virus. Glob. Vet. 2010, 4, 616–621. [Google Scholar]
- Issa, K.J.; Omar, J.A. Effect of garlic powder on performance and lipid profile of broilers. Open J. Anim. Sci. 2012, 2, 18526. [Google Scholar] [CrossRef] [Green Version]
- Al-Homidan, A. Efficacy of using different sources and levels of Allium sativum and Zingiber officinale on broiler chicks performance. Saudi J. Biol. Sci. 2005, 12, 96–102. [Google Scholar]
- Aji, S.B.; Ignatius, K.; Ado, A.; Nuhu, J.B.; Abdulkarim, A.; Aliyu, U.; Gambo, M.B.; Ibrahim, M.A.; Abubakar, H.; Bukar, M.M. Effects of feeding onion (Allium cepa) and garlic (Allium sativum) on some performance characteristics of broiler chickens. Res. J. Poult. Sci. 2011, 4, 22–27. [Google Scholar]
- Choi, I.; Park, W.; Kim, Y. Effects of dietary garlic powder and α-tocopherol supplementation on performance, serum cholesterol levels, and meat quality of chicken. Poult. Sci. 2010, 89, 1724–1731. [Google Scholar] [CrossRef] [PubMed]
- Onibi, G.E.; Adebisi, O.E.; Fajemisin, A.N. Response of broiler chickens in terms of performance and meat quality to garlic (Allium sativum) supplementation. Afr. J. Agric. Res. 2009, 4, 511–517. [Google Scholar]
- Adebiyi, F.G.; Ologhobo, A.D.; Adejumo, I.O. Raw Allium sativum as performance enhancer and hypocholesterolemic agent in laying hens. Asian J. Anim. Vet. Adv. 2018, 13, 210–217. [Google Scholar]
- Ao, X.; Yoo, J.; Zhou, T.; Wang, J.; Meng, Q.; Yan, L.; Cho, J.; Kim, I. Effects of fermented garlic powder supplementation on growth performance, blood profiles and breast meat quality in broilers. Livest. Sci. 2011, 141, 85–89. [Google Scholar] [CrossRef]
- Olobatoke, R.; Mulugeta, S. Effect of dietary garlic powder on layer performance, fecal bacterial load, and egg quality. Poult. Sci. 2011, 90, 665–670. [Google Scholar] [CrossRef]
- Yalcin, S.; Onbasilar, I.; Sehu, A.; Yalcin, S. The effects of dietary garlic powder on the performance, egg traits and blood serum cholesterol of laying quails. Asian Australas. J. Anim. Sci. 2007, 20, 944. [Google Scholar] [CrossRef]
- Ao, X.; Yoo, J.; Lee, J.; Jang, H.; Wang, J.; Zhou, T.; Kim, I. Effects of fermented garlic powder on production performance, egg quality, blood profiles and fatty acids composition of egg yolk in laying hens. Asian-Australas. J. Anim. Sci. 2010, 23, 786–791. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, S.-H.; Lee, S.-J.; Gadde, U.D.; Oh, S.-T.; Han, H.; Lillehoj, H.S. Effects of dietary Allium hookeri root on growth performance and antioxidant activity in young broiler chickens. Res. Vet. Sci. 2018, 118, 345–350. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, S.-H.; Gadde, U.D.; Oh, S.-T.; Lee, S.-J.; Lillehoj, H.S. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Bou, R.; Codony, R.; Tres, A.; Decker, E.A.; Guardiola, F. Dietary strategies to improve nutritional value, oxidative stability, and sensory properties of poultry products. Crit. Rev. Food Sci. Nutr. 2009, 49, 800–822. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Lillehoj, E.P.; Bravo, D. Improved resistance to Eimeria acervulina infection in chickens due to dietary supplementation with garlic metabolites. Br. J. Nutr. 2013, 109, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Peinado, M.J.; Ruiz, R.; Echávarri, A.; Rubio, L.A. Garlic derivative propyl propane thiosulfonate is effective against broiler enteropathogens in vivo. Poult. Sci. 2012, 91, 2148–2157. [Google Scholar] [CrossRef] [PubMed]
- Peinado, M.; Ruiz, R.; Echávarri, A.; Aranda-Olmedo, I.; Rubio, L. Garlic derivative PTS-O modulates intestinal microbiota composition and improves digestibility in growing broiler chickens. Anim. Feed Sci. Technol. 2013, 181, 87–92. [Google Scholar] [CrossRef]
- Lim, K.; You, S.; An, B.; Kang, C. Effects of dietary garlic powder and copper on cholesterol content and quality characteristics of chicken eggs. Asian-Australas. J. Anim. Sci. 2006, 19, 582–586. [Google Scholar] [CrossRef]
- Prasad, R.; Rose, M.; Virmani, M.; Garg, S.; Puri, J. Effect of garlic (Allium sativum) supplementation on haematological parameters in chicken (Gallus domesticus). Indian J. Anim. Res. 2009, 43, ARCC1589. [Google Scholar]
- An, B.; Kim, J.; Oh, S.; Kang, C.; Cho, S.; Kim, S. Effects of onion extracts on growth performance, carcass characteristics and blood profiles of white mini broilers. Asian-Australas. J. Anim. Sci. 2015, 28, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Aditya, S.; Ahammed, M.; Jang, S.H.; Ohh, S.J. Effects of dietary onion (Allium cepa) extract supplementation on performance, apparent total tract retention of nutrients, blood profile and meat quality of broiler chicks. Asian-Australas. J. Anim. Sci. 2017, 30, 229–235. [Google Scholar] [CrossRef]
- Karangiya, V.; Savsani, H.; Patil, S.S.; Garg, D.; Murthy, K.; Ribadiya, N.; Vekariya, S. Effect of dietary supplementation of garlic, ginger and their combination on feed intake, growth performance and economics in commercial broilers. Vet. World 2016, 9, 245–250. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Smith, T. Effects of dietary garlic on cholesterol metabolism in laying hens. Poult. Sci. 2002, 81, 1856–1862. [Google Scholar] [CrossRef]
- Yalcın, S.; Onbaşılar, E.E.; Reisli, Z.; Yalcın, S. Effect of garlic powder on the performance, egg traits and blood parameters of laying hens. J. Sci. Food Agric. 2006, 86, 1336–1339. [Google Scholar] [CrossRef]
- Canogullari, S.; Baylan, M.; Erdogan, Z.; Duzguner, V.; Kucukgul, A. The effects of dietary garlic powder on performance, egg yolk and serum cholesterol concentrations in laying quails. Czech J. Anim. Sci. 2010, 55, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Omer, H.A.; Ahmed, S.M.; Abdel-Magid, S.S.; El-Mallah, G.M.; Bakr, A.A.; Fattah, M.M.A. Nutritional impact of inclusion of garlic (Allium sativum) and/or onion (Allium cepa L.) powder in laying hens’ diets on their performance, egg quality, and some blood constituents. Bull. Natl. Res. Cent. 2019, 43, 23. [Google Scholar] [CrossRef]
- Mahmoud, K.Z.; Gharaibeh, S.M.; Zakaria, H.A.; Qatramiz, A.M. Garlic (Allium sativum) supplementation: Influence on egg production, quality, and yolk cholesterol level in layer hens. Asian-Australas. J. Anim. Sci. 2010, 23, 1503–1509. [Google Scholar] [CrossRef]
- Horton, G.; Fennell, M.; Prasad, B. Effect of dietary garlic (Allium sativum) on performance, carcass composition and blood chemistry changes in broiler chickens. Can. J. Anim. Sci. 1991, 71, 939–942. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Abuirmeileh, N.; Din, Z.Z.; Elson, C.E.; Burger, W.C. Inhibition of cholesterol and fatty acid biosynthesis in liver enzymes and chicken hepatocytes by polar fractions of garlic. Lipids 1983, 18, 343–348. [Google Scholar] [CrossRef]
- Freeman, F.; Kodera, Y. Garlic chemistry: Stability of S-(2-propenyl)-2-propene-1-sulfinothioate (allicin) in blood, solvents, and simulated physiological fluids. J. Agric. Food Chem. 1995, 43, 2332–2338. [Google Scholar] [CrossRef]
- Elmowalid, G.A.; El-Hamid, M.I.A.; El-Wahab, A.M.A.; Atta, M.; El-Naser, G.A.; Attia, A.M. Garlic and ginger extracts modulated broiler chicks innate immune responses and enhanced multidrug resistant Escherichia coli O78 clearance. Comp. Immunol. Microbiol. Infect. Dis. 2019, 66, 101334. [Google Scholar] [CrossRef]
- Jimoh, A.; Ibitoye, E.; Dabai, Y.; Garba, S. In vivo antimicrobial potentials of garlic against Clostridium perfringens and its promotant effects on performance of broiler chickens. Pak. J. Biol. Sci. 2013, 16, 1978–1984. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Chand, N.; Khan, R.U.; Naz, S.; Gul, S. Anticoccidial effect of garlic (Allium sativum) and ginger (Zingiber officinale) against experimentally induced coccidiosis in broiler chickens. J. App. Anim. Res. 2019, 47, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Kavindra, S.; Shalini, N. Studies on the anthelmintic activity of Allium sativum (garlic) oil on common poultry worms Ascaridia galli and Heterakis gallinae. J. Parasitol. Appl. Anim. Biol. 2000, 9, 47–52. [Google Scholar]
- Shojai, T.M.; Langeroudi, A.G.; Karimi, V.; Barin, A.; Sadri, N. The effect of Allium sativum (Garlic) extract on infectious bronchitis virus in specific pathogen free embryonic egg. Avicenna J. Phytomed. 2016, 6, 458. [Google Scholar]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Ruiz, R.; Peinado, M.; Aranda-Olmedo, I.; Abecia, L.; Suárez-Pereira, E.; Mellet, C.O.; Fernandez, J.G.; Rubio, L. Effects of feed additives on ileal mucosa–associated microbiota composition of broiler chickens. J. Anim. Sci. 2015, 93, 3410–3420. [Google Scholar] [CrossRef]
- Goodarzi, M.; Nanekarani, S.; Landy, N. Effect of dietary supplementation with onion (Allium cepa L.) on performance, carcass traits and intestinal microflora composition in broiler chickens. Asian Pac. J. Trop. Dis. 2014, 4, S297–S301. [Google Scholar] [CrossRef]
- Shargh, M.S.; Dastar, B.; Zerehdaran, S.; Khomeiri, M.; Moradi, A. Effects of using plant extracts and a probiotic on performance, intestinal morphology, and microflora population in broilers. J. Appl. Poult. Res. 2012, 21, 201–208. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Kim, G.-B.; Seo, Y.; Kim, C.; Paik, I. Effect of dietary prebiotic supplementation on the performance, intestinal microflora, and immune response of broilers. Poult. Sci. 2011, 90, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Adibmoradi, M.; Navidshad, B.; Seifdavati, J.; Royan, M. Effect of dietary garlic meal on histological structure of small intestine in broiler chickens. J. Poult. Sci. 2006, 43, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, S.; Rehman, A.; Yousaf, M.; Akhtar, P.; Abbas, G.; Hayat, K.; Mahmood, A.; Shahzad, M.K. Comparative efficacy of different herbal plant’s leaf extract on haematology, intestinal histomorphology and nutrient digestibility in broilers. Adv. Zool. Bot. 2015, 3, 11–16. [Google Scholar]
- Torok, V.A.; Hughes, R.J.; Mikkelsen, L.L.; Perez-Maldonado, R.; Balding, K.; MacAlpine, R.; Percy, N.J.; Ophel-Keller, K. Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials. Appl. Environ. Microbiol. 2011, 77, 5868–5878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravindran, V.; Tancharoenrat, P.; Zaefarian, F.; Ravindran, G. Fats in poultry nutrition: Digestive physiology and factors influencing their utilisation. Anim. Feed Sci. Technol. 2016, 213, 1–21. [Google Scholar] [CrossRef]
- Thomas, F.; Hehemann, J.-H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [Green Version]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef]
- Rubio, L.; Peinado, M.; Ruiz, R.; Suárez-Pereira, E.; Mellet, C.O.; Fernandez, J.G. Correlations between changes in intestinal microbiota composition and performance parameters in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2015, 99, 418–423. [Google Scholar] [CrossRef]
- Dieumou, F.; Teguia, A.; Kuiate, J.; Tamokou, J.; Fonge, N.; Dongmo, M. Effects of ginger (Zingiber officinale) and garlic (Allium sativum) essential oils on growth performance and gut microbial population of broiler chickens. Livest. Res. Rural Dev. 2009, 21, 23–32. [Google Scholar]
- Kırkpınar, F.; Ünlü, H.B.; Özdemir, G. Effects of oregano and garlic essential oils on performance, carcase, organ and blood characteristics and intestinal microflora of broilers. Livest. Sci. 2011, 137, 219–225. [Google Scholar] [CrossRef]
- Hanieh, H.; Narabara, K.; Piao, M.; Gerile, C.; Abe, A.; Kondo, Y. Modulatory effects of two levels of dietary Alliums on immune response and certain immunological variables, following immunization, in White Leghorn chickens. Anim. Sci. J. 2010, 81, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, H.; Narabara, K.; Tanaka, Y.; Gu, Z.; Abe, A.; Kondo, Y. Immunomodulatory effects of Alliums and Ipomoea batata extracts on lymphocytes and macrophages functions in White Leghorn chickens: In vitro study. Anim. Sci. J. 2012, 83, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Jafari, R.; Ghorbanpoor, M.; Diarjan, S.H. Effect of dietary garlic on serum antibody titer against Newcastle disease vaccine in broiler chicks. J. Biol. Sci. 2008, 8, 1258–1260. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yue, H.; Wu, S.; Zhang, H.; Qi, G. Nutritional modulation of health, egg quality and environmental pollution of the layers. Anim. Nutr. 2017, 3, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Damaziak, K.; Riedel, J.; Gozdowski, D.; Niemiec, J.; Siennicka, A.; Róg, D. Productive performance and egg quality of laying hens fed diets supplemented with garlic and onion extracts. J. Appl. Poult. Res. 2017, 26, 337–349. [Google Scholar] [CrossRef]
- González-Morales, S.; Pérez-Labrada, F.; García-Enciso, E.L.; Leija-Martínez, P.; Medrano-Macías, J.; Dávila-Rangel, I.E.; Juárez-Maldonado, A.; Rivas-Martínez, E.N.; Benavides-Mendoza, A. Selenium and sulfur to produce Allium functional crops. Molecules 2017, 22, 558. [Google Scholar] [CrossRef] [Green Version]
- Chung, L.Y. The antioxidant properties of garlic compounds: Allyl cysteine, alliin, allicin, and allyl disulfide. J. Med. Food 2006, 9, 205–213. [Google Scholar] [CrossRef]
- Wakebe, M. Organic Selenium and Egg Freshness. Feed for Meat Chickens and Feed for Laying Hens. Japanese Patent 10-23864, 27 January 1998. [Google Scholar]
- Motozono, Y.; Li, S.; Ishihara, N.; Arai, N.; Nakahara, R.; Kochi, Y.; Tanaka, S.; Kami, A. Poultry Egg with Improved Flavor and Method of Producing. U.S. Patent 5,882,701, 16 March 1999. [Google Scholar]
- Birrenkott, G.; Brockenfelt, G.; Owens, M.; Halpin, E. Yolk and blood cholesterol levels and organoleptic assessment of eggs from hens fed a garlic-supplemented diet. Poult. Sci. 2000, 79, 75. [Google Scholar]
- Abdullah, A.Y.; Al-Beitawi, N.A.; Rjoup, M.M.S.; Qudsieh, R.I.; Ishmais, M.A.A. Growth performance, carcass and meat quality characteristics of different commercial crosses of broiler strains of chicken. J. Poult. Sci. 2010, 47, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Chowdhury, M.; Huo, Y.; Gong, J. Phytogenic compounds as alternatives to in-feed antibiotics: Potentials and challenges in application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, H.; Watanabe, K.; Suma, K.; Origuchi, K.; Matsufuji, H.; Seki, T.; Ariga, T. Antibacterial potential of garlic-derived allicin and its cancellation by sulfhydryl compounds. Biosci. Biotechnol. Biochem. 2009, 73, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Luo, X.; Wang, Y.; Feng, Y. Evaluation of zinc oxide nanoparticles on lettuce (Lactuca sativa L.) growth and soil bacterial community. Environ. Sci. Pollut. Res. 2018, 25, 6026–6035. [Google Scholar] [CrossRef] [PubMed]
- Hafeez, A.; Männer, K.; Schieder, C.; Zentek, J. Effect of supplementation of phytogenic feed additives (powdered vs. encapsulated) on performance and nutrient digestibility in broiler chickens. Poult. Sci. 2015, 95, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Niu, K.-M.; Lee, J.-H.; Cho, S.; Han, S.-G.; Kim, S.-K. Characteristics of Chinese chives (Allium tuberosum) fermented by Leuconostoc mesenteroides. Appl. Biol. Chem. 2016, 59, 349–357. [Google Scholar] [CrossRef]
- Pan, M.; Wu, Q.; Tao, X.; Wan, C.; Shah, N.P.; Wei, H. Fermentation of Allium chinense bulbs with Lactobacillus plantarum ZDY 2013 shows enhanced biofunctionalities, and nutritional and chemical properties. J. Food Sci. 2015, 80, M2272–M2278. [Google Scholar] [CrossRef]
- Yang, J.; Ji, Y.; Park, H.; Lee, J.; Park, S.; Yeo, S.; Shin, H.; Holzapfel, W.H. Selection of functional lactic acid bacteria as starter cultures for the fermentation of Korean leek (Allium tuberosum Rottler ex Sprengel.). Int. J. Food Microbiol. 2014, 191, 164–171. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.-L.; Lee, S.; Lee, S.-Y.; Ko, S.; Yoo, M. UPLC/ESI-MS/MS analysis of compositional changes for organosulfur compounds in garlic (Allium sativum L.) during fermentation. Food Chem. 2016, 211, 555–559. [Google Scholar] [CrossRef]
- Peres, C.M.; Alves, M.; Hernandez-Mendoza, A.; Moreira, L.; Silva, S.; Bronze, M.R.; Vilas-Boas, L.; Peres, C.; Malcata, F.X. Novel isolates of lactobacilli from fermented Portuguese olive as potential probiotics. LWT-Food Sci. Technol. 2014, 59, 234–246. [Google Scholar] [CrossRef] [Green Version]
- Bernaert, N.; De Paepe, D.; Bouten, C.; De Clercq, H.; Stewart, D.; Van Bockstaele, E.; De Loose, M.; Van Droogenbroeck, B. Antioxidant capacity, total phenolic and ascorbate content as a function of the genetic diversity of leek (Allium ampeloprasum var. porrum). Food Chem. 2012, 134, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Lee, S.; Kim, I. Effect of dietary Korean aged garlic extract by Leukonostoc citreum SK2556 on production, hematological status, meat quality, relative organ weight, targeted Escherichia coli colony and excreta gas emission in broilers. Anim. Feed Sci. Technol. 2014, 198, 333–340. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, S.J.; Yu, H.J.; Lee, J.H.; Cho, H.Y. Fermentation with Lactobacillus enhances the preventive effect of garlic extract on high fat diet-induced hepatic steatosis in mice. J. Funct. Foods 2017, 30, 125–133. [Google Scholar] [CrossRef]
- Sato, E.; Kohno, M.; Hamano, H.; Niwano, Y. Increased anti-oxidative potency of garlic by spontaneous short-term fermentation. Plant Foods Hum. Nutr. 2006, 61, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.-J.; Kim, S.-I.; Park, S.-Y.; Bang, H.-Y.; Jeong, J.H.; So, J.-H.; Rhee, I.-K.; Song, K.-S. Fermentation enhances the in vitro antioxidative effect of onion (Allium cepa) via an increase in quercetin content. Food Chem. Toxicol. 2012, 50, 2042–2048. [Google Scholar] [CrossRef]
- Millet, A.; Lamy, E.; Jonas, D.; Stintzing, F.; Mersch-Sundermann, V.; Merfort, I. Fermentation enhances the biological activity of Allium cepa bulb extracts. J. Agric. Food Chem. 2012, 60, 2148–2156. [Google Scholar] [CrossRef]
- Chung, D.-M.; Chung, Y.-C.; Maeng, P.J.; Chun, H.-K. Regioselective deglycosylation of onion quercetin glucosides by Saccharomyces cerevisiae. Biotechnol. Lett. 2011, 33, 783–786. [Google Scholar] [CrossRef]
- Cheng, L.; Luo, J.; Li, P.; Yu, H.; Huang, J.; Luo, L. Microbial diversity and flavor formation in onion fermentation. Food Funct. 2014, 5, 2338–2347. [Google Scholar] [CrossRef]
- Bisakowski, B.; Atwal, A.S.; Gardner, N.; Champagne, C.P. Effect of lactic acid fermentation of onions (Allium cepa) on the composition of flavonol glucosides. Int. J. Food Sci. Technol. 2007, 42, 783–789. [Google Scholar] [CrossRef]
- Wouters, D.; Bernaert, N.; Anno, N.; Van Droogenbroeck, B.; De Loose, M.; Van Bockstaele, E.; De Vuyst, L. Application and validation of autochthonous lactic acid bacteria starter cultures for controlled leek fermentations and their influence on the antioxidant properties of leek. Int. J. Food Microbiol. 2013, 165, 121–133. [Google Scholar] [CrossRef]
- Bernaert, N.; Wouters, D.; De Vuyst, L.; De Paepe, D.; De Clercq, H.; Van Bockstaele, E.; De Loose, M.; Van Droogenbroeck, B. Antioxidant changes of leek (Allium ampeloprasum var. porrum) during spontaneous fermentation of the white shaft and green leaves. J. Sci. Food Agric. 2013, 93, 2146–2153. [Google Scholar]
- Milea, Ș.-A.; Aprodu, I.; Vasile, A.M.; Barbu, V.; Râpeanu, G.; Bahrim, G.E.; Stănciuc, N. Widen the functionality of flavonoids from yellow onion skins through extraction and microencapsulation in whey proteins hydrolysates and different polymers. J. Food Eng. 2019, 251, 29–35. [Google Scholar] [CrossRef]
- Akdeniz, B.; Sumnu, G.; Sahin, S. The effects of maltodextrin and gum arabic on encapsulation of onion skin phenolic compounds. Chem. Eng. Trans. 2017, 57, 1891–1896. [Google Scholar]
- Piletti, R.; Zanetti, M.; Jung, G.; de Mello, J.M.M.; Dalcanton, F.; Soares, C.; Riella, H.G.; Fiori, M.A. Microencapsulation of garlic oil by β-cyclodextrin as a thermal protection method for antibacterial action. Mater. Sci. Eng. C 2019, 94, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Veršič, R.J. The Economics of Microencapsulation in the Food Industry. In Microencapsulation in the Food Industry: A Practical Implementation Guide; Elsevier: Cambridge, MA, USA, 2014; pp. 409–417. [Google Scholar]
- Gangadoo, S.; Stanley, D.; Hughes, R.J.; Moore, R.J.; Chapman, J. Nanoparticles in feed: Progress and prospects in poultry research. Trends Food Sci. Technol. 2016, 58, 115–126. [Google Scholar] [CrossRef]
- Bhavsar, S. Application of Nanotechnology for Phyto Constituents. Arch. Nanomed. Open Access J. 2018, 1, 6–7. [Google Scholar] [CrossRef]
- Sundari, Z.; Yuwanta, T.; Martien, R. Effect of nanocapsule level on broiler performance and fat deposition. Int. J. Poult. Sci. 2014, 13, 31–35. [Google Scholar]
- Jini, D.; Sharmila, S. Green synthesis of silver nanoparticles from Allium cepa and its in vitro antidiabetic activity. Mater. Today Proc. 2019, in press. [Google Scholar] [CrossRef]


{kind=link}
| Animals | Allium spp. & treatments | Formulation | Main active components | Effects | Ref. |
|---|---|---|---|---|---|
| Male Shaver Starbo broilers (7-d-old) | 0.5 or 5 g A. sativum/kg feed; 8 weeks | Raw and boiled Powder | ND | ↑ BWG; ↑ Oxidative stability of meat during storage | [64] |
| 104 Mixed sex broilers (4-week-old) | A. cepa (0, 0.025, 0.05, or 0.1 g) and A. sativum (0, 0.025, 0.05, or 0.1 g)/kg feed; 3 weeks | Powder | ND | ↑ BWG; ↑ Feed and water intake; ↑ FCR | [62] |
| 320 Ross-308 broilers (1-d-old) | 1.5, 2.0, or 2.5 g A. cepa/kg feed; 6 weeks | Powder | ND | ↑ Body weight; ↑ Feed intake; ↑ Lactobacillus and Streptococcus; ↓ E. coli; ↑ Intestinal histomorphology | [53] |
| 60 Ross broilers (1-d-old) (Challenged by live Eimeria acervulina) | 0.01 g Garlicon40® (A. sativum)/kg feed; 20 days | Powder | 67% Propyl thiosulfinate and 33% propyl thiosulfinate oxide | ↑ BWG; ↓ Fecal oocyst shedding; ↑ E. acervulina profilin antibody responses | [73] |
| 144 Male Cobb broilers (1-d-old) | 0.045 or 0.09 g Proallium-SO-DMC® (A. sativum)/kg feed; 3 weeks | Powder | 11.3% Propyl propane thiosulfonate | ↑ Body weight; ↓ Feed intake; ↓ Feed/gain ratio; ↓ Enteropathogens; ↑ Ileal histological structure | [74] |
| 144 Male Cobb broilers (1-d-old) | 0.045 or 0.09 g Proallium-SO-DMC® (A. sativum)/kg feed; 3 weeks | Powder | 11.3% Propyl propane thiosulfonate | ↑ Nutrients digestibility; Modulate intestinal microbiota | [75] |
| 300 Male Arbor Acres broilers (1-d-old) | 10–50 g A. sativum/kg feed; 5 weeks | Powder | ND | ↓ Total serum cholesterol levels; ↓ LDL-C; ↑ HDL-C; ↑ Meat quality (↑ Color stability; ↓ TBARS; ↑ Total unsaturated fatty acid:total saturated fatty acid ratios) | [63] |
| 150 Hy-Line Brown layers (50-week-old) | 10–50 g A. sativum/kg feed; 5 weeks | Powder | ND | ↑ Haugh Unit; ↓ Total serum cholesterol; ↓ Total egg yolk cholesterol | [76] |
| 24 Mixed sex broilers (1-d-old) | 15 or 30 g A. sativum/kg feed; 8 weeks | Powder | ND | Regulate lipid metabolism (↓ Plasma cholesterol; ↓ triglycerides; ↓ VLDL-C; ↓ LDL-C; ↑ HDL-C) | [77] |
| 150 Male Ross-708 broilers (1-d-old) (Challenged by LPS | 10 g or 50 g A. hookeri/kg feed; 8 days | Powdered fermented roots | ND | ↑ Body weight; ↓ Inflammatory response (↑ Expression of intestinal tight junction proteins and mucin; ↓ Serum α-1-AGP; ↓ Pro-inflammatory cytokines) | [49] |
| 125 Male Ross-708 broilers (1-d-old) | 10 g A. hookeri/kg feed; 3 weeks | Powdered roots | ND | ↑ Body weight; ↑ antioxidant activity (↑Gene expression of heme oxygenase, aflatoxin B1 aldehyde reductase, SOD 1, and CAT; ↑Serum levels of SOD, CAT, and MDA) | [70] |
| 300 male Ross-708 broiler chicks (1-d-old) (Challenged by NE) | 10 g or 30 g A. hookeri/kg feed; 20 days | Powdered roots | ND | ↓ Loss of BWG; ↓ Lesion; ↓ Fecal oocyst shedding; ↑ Innate immunity (↓ Pro-inflammatory cytokines; ↑ Expression of tight gut junction proteins and mucin) | [71] |
| 500 Arbor Acres broiler chicks (1-d-old) | 1–4 g A. sativum/kg feed; 5 weeks | Fermented powder | ND | ↑ WBC, lymphocyte and IgG; ↓ Total cholesterol; ↓ Triglyceride; ↓ LDL-C; ↓ Cortisol; ↑ Meat quality (↓ TBARS and pH) | [66] |
| 600 White mini broiler chicks (1-d-old) | 3 or 5 mL A. cepa/kg feed; 5 weeks | Fermented liquid | ND | ↑ Final body weight; ↑BWG; ↓ Serum cholesterol; ↓ Triacylglycerol | [78] |
| 400 Male Ross-308 broiler chicks (3-d-old) | 5, 7.5, or 10 g A. cepa/kg feed; 4 weeks | Liquid extract | ND | ↑ BWG; ↑ Feed intake; ↑ ATTR; ↑ Serum IgG; ↓ TBARS | [79] |
| 240 Cobb-400 broiler chicks (1 day old) | 10 g A. sativum/kg feed; 6 weeks | Powder | ND | ↑ BWG; ↑ Final weight; ↑ Villi length and width; ↑ Cryptal depth | [80] |
| 288 commercial broiler chicks (1-d-old) | 5 or 10 g A. sativum+1 or 2 g Piper nigram/kg feed; 6 weeks | Powder | ND | ↑ BWG; ↑ Final weight; ↑ FCR | [57] |
| 36 layers of six different strains (Hisex Brown, Isa Brown, Lohmann, Starcross, Babcock, and Starcross-579 strains) (28-week-old) | 20, 40, 60, 80, or 100 g A. sativum/kg feed; 6 weeks. | Paste (sun-dried) | ND | ↑ Egg production; ↑ Egg yolk weight; ↓ Egg yolk cholesterol; ↓ Serum cholesterol | [81] |
| 162 SHSY-type brown layers (21-week-old) | 5 or 10 g A. sativum/kg feed; 22 weeks | Powder | ND | ↑ Egg weight; ↓ Egg yolk cholesterol; ↓ Serum triglyceride and cholesterol | [82] |
| 300 Quails (9-week-old) | 5 or 10 g A. sativum/kg feed; 21 weeks | Powder | ND | ↑ Egg weight; ↓ Egg yolk cholesterol; ↓ Serum cholesterol | [68] |
| 72 Dekalb white layers (30-week-old) | 30 or 50 mg A. sativum/kg feed; 7 weeks | Powder | ND | ↑ Albumen height; ↑ Haugh units; ↑ Egg and albumen weight | [67] |
| 120 laying Japanese quails (10-week-old) | 10, 20, or 40 g A. sativum/kg feed; 12 weeks | Powder | ND | ↓ Egg yolk cholesterol; ↓ Serum cholesterol | [83] |
| 240 Isa brown laying hens (41-week-old) | 10, 20, or 40 g A. sativum/kg feed; 5 weeks | Fermented powder | ND | ↑ Yolk height and color; ↑ Haugh Unit; ↓ Serum total cholesterol; ↑ Yolk PUFA:SFA ratio | [69] |
| 108 laying hens (30-week-old) | 5 or 10 g A. sativum and 10 g A. cepa/kg feed; 4 weeks | Powder | ND | ↑ FCR; ↑ Egg production; ↑ Egg weight; ↓ Total cholesterol; ↓ Creatinine | [84] |
| 180 Isa Brown hens (18-week-old) | 10, 20, 30, 40, or 50 g A. sativum/kg feed; 20 weeks | Paste (Raw) | ND | ↑ HDL-C; ↓ Total cholesterol; ↓ Yolk cholesterol; ↓ LDL-C | [65] |
| 640 Mixed sex Ross 308 broiler chickens (body weight 45 ± 7 g) | 1, 1.5, or 2.25 mL/kg feed; 6 weeks | Liquid | ND | ↓ Mortality; ↓ FCR; ↑ EPEF; ↑ Dressing percentage; ↑ Weight of breast muscles; ↑ Liver weight; ↑ Protein and crude ash content of breast meat; ↑ Total protein content of serum | [55] |
| Items | Characteristics | Effects |
|---|---|---|
| Thiosulfinates | Volatile | Varied final concentration in feed |
| Pungent smell | Feed palatability | |
| Reacts with SH groups of other feed constituents | Loss of antimicrobial activity | |
| Thermal instability | Difficult in feed processing | |
| Poor water solubility | ||
| Formulation | Variation in chemical constituents | Variation in biological response |
| Dosage | Pungent smell | Feed palatability |
| Other factors | Subjects (broilers, layers, quails, etc.), age, health status, feed type and quality, environmental conditions, and duration of study | Inconsistent results |
| Plants | Microorganisms | Fermentation Conditions | Compositional Changes | Study | Biological Activities | Ref. |
|---|---|---|---|---|---|---|
| A. sativum | Saccharomyces cerevisiae; Lactobacillus plantarum; Mimulus pilosus | S. cerevisiae and L. plantarum at 25 and 37 °C, respectively, for 48 h and M. pilosus at 25 °C for 7 days | ↑ S-allyl-l-cysteine and cycloalliin; ↓ γ-Glutamyl peptide | - | ND | [130] |
| A. sativum | L. plantarum | 37 °C for 24 h | Alliin content ↓; ↑ Cycloalliin content; ↑ S-allyl cysteine content | In vivo (mice model) | Lipid metabolism and antioxidant | [134] |
| A. sativum | Spontaneous | 40 days at 60–70 °C and 85–95% relative humidity | ↑ Polyphenol content | In vitro | Antioxidant | [135] |
| A. cepa | Aspergillus kawachii crude enzyme extract | 30 °C for 24 h | ↑ Quercetin and quercetin-3-glucoside; ↓ quercetin-3,4′-diglucoside and -4′-glucoside | In vitro | Antioxidant and neuroprotection | [136] |
| A. cepa | Spontaneous | 37 °C for 3.5 days | Flavonoid profile changed | In vitro | Antibacterial, antigenotoxic, and antiproliferative | [137] |
| A. cepa | Aspergillus oryzae, Bacillus subtilis, L. plantarum, and S. cerevisiae | 30 °C for 3 days | ↑ Isoquercitrin | In vitro | [138] | |
| A. cepa | Spontaneous | 96–108 h with or without 1% (w/v) salt | ↑ Lactic acid and acetic acid; ↓ amino acids; ↑ esters, alcohols, and aldehydes | In vitro | Flavor | [139] |
| A. cepa | L. plantarum | 19 °C for 48 h | ↑ Quercetin diglucoside | In vitro | Antioxidant | [140] |
| A. ampeloprasum var. porrum | L. plantarum, Leuconostoc mesenteroides, and Lactobacillus sakei | 18 °C for 3 weeks | ND | In vitro | Sensory properties | [141] |
| A. ampeloprasum var. porrum | Spontaneous | 18 °C for 21 days | ↑ Polyphenol; ↓ methiin and isoalliin | In vitro | Antioxidant | [142] |
| A. tuberosum | L. mesenteroides | 30 °C for 3 days | ↑ Polyphenol | In vitro | Antibacterial and antioxidant | [127] |
| A. chinense | L. plantarum | 37 °C for 7 days | ↑ Polyphenol content; ↑ Free amino acid content; ↓ Sulfur containing compounds | In vitro | Antioxidant and flavor | [128] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kothari, D.; Lee, W.-D.; Niu, K.-M.; Kim, S.-K. The Genus Allium as Poultry Feed Additive: A Review. Animals 2019, 9, 1032. https://doi.org/10.3390/ani9121032
Kothari D, Lee W-D, Niu K-M, Kim S-K. The Genus Allium as Poultry Feed Additive: A Review. Animals. 2019; 9(12):1032. https://doi.org/10.3390/ani9121032
Chicago/Turabian StyleKothari, Damini, Woo-Do Lee, Kai-Min Niu, and Soo-Ki Kim. 2019. "The Genus Allium as Poultry Feed Additive: A Review" Animals 9, no. 12: 1032. https://doi.org/10.3390/ani9121032
APA StyleKothari, D., Lee, W. -D., Niu, K. -M., & Kim, S. -K. (2019). The Genus Allium as Poultry Feed Additive: A Review. Animals, 9(12), 1032. https://doi.org/10.3390/ani9121032