Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
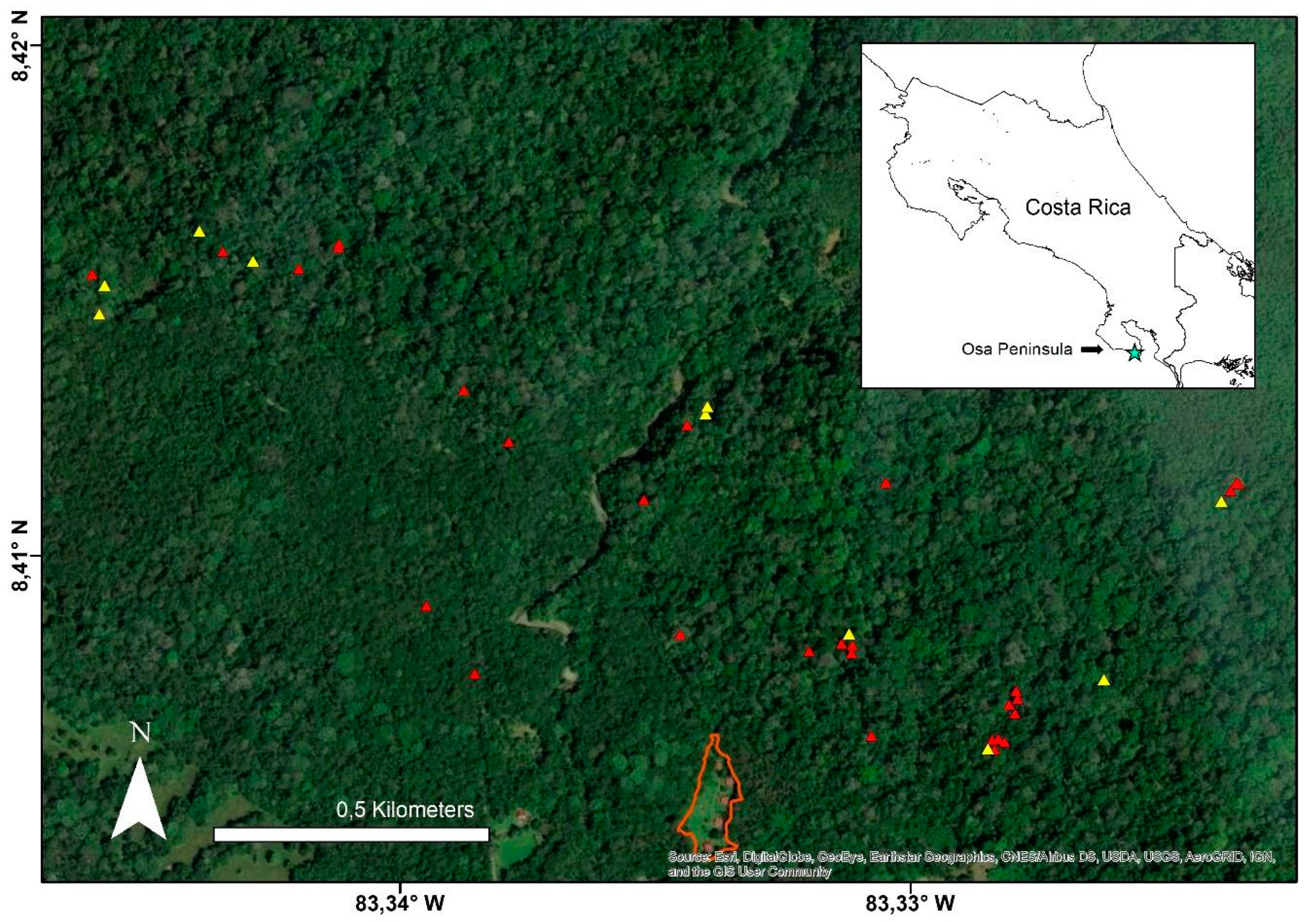
2.1. Study Site
2.2. Sleeping Site Identification
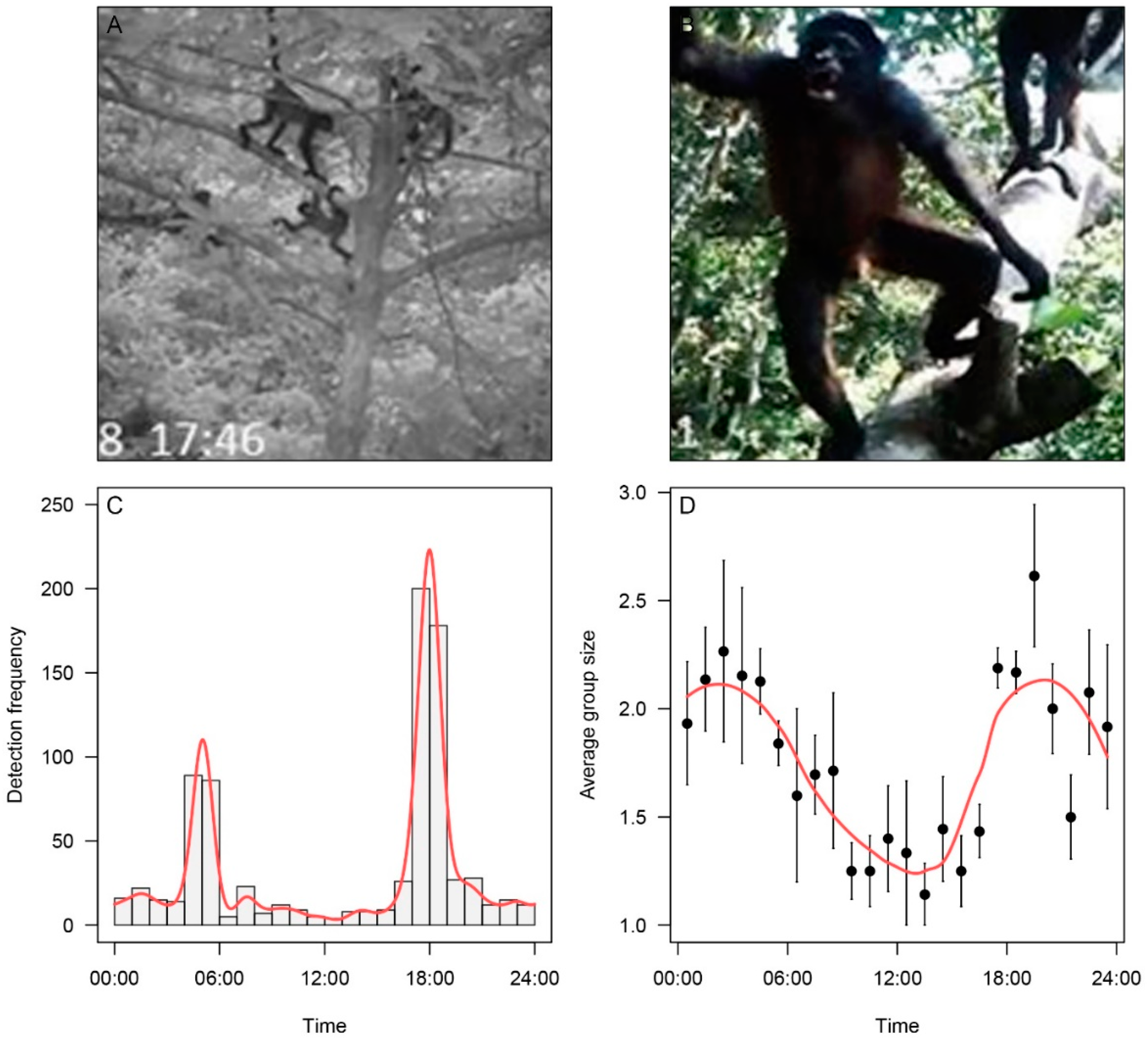
2.3. Characterizing Sleeping Site Usage with Arboreal Camera Traps
2.4. Characterizing Latrine Use by Terrestrial Vertebrates
2.5. Characterizing Latrine Use by Dung Beetles
2.6. Characterizing the Effects of Latrines on Seedling Density and Soil Quality
2.7. Data Analysis
3. Results
3.1. Use of Sleeping Sites
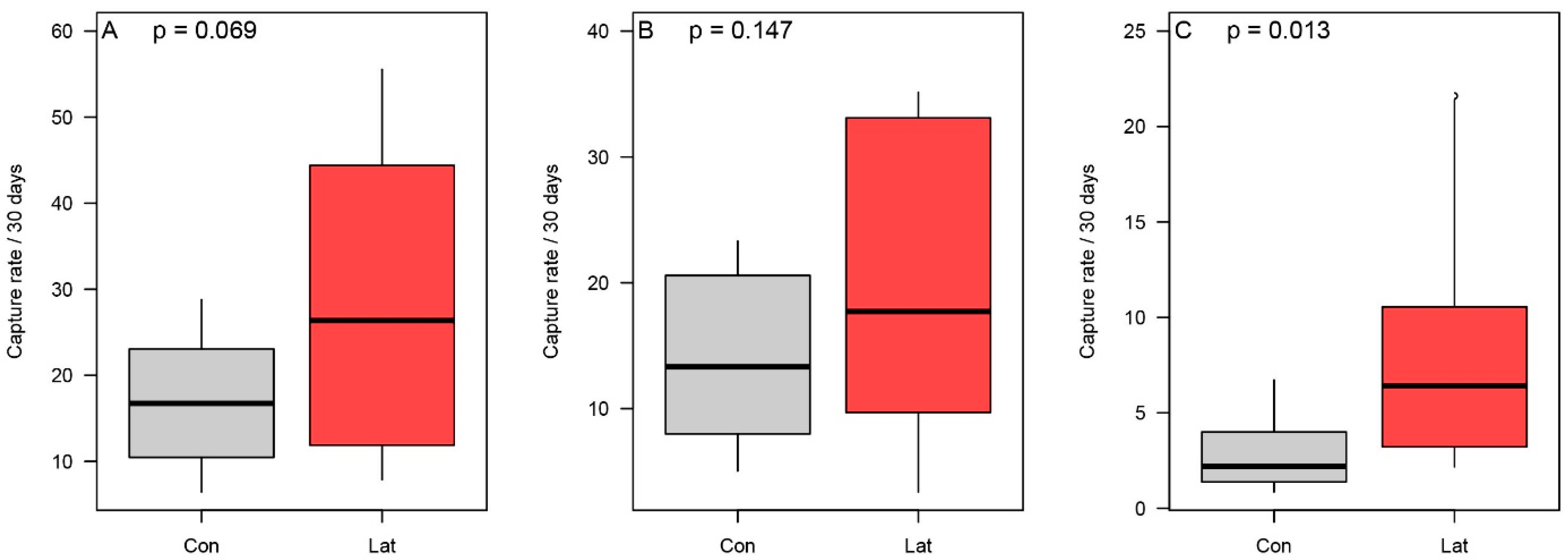
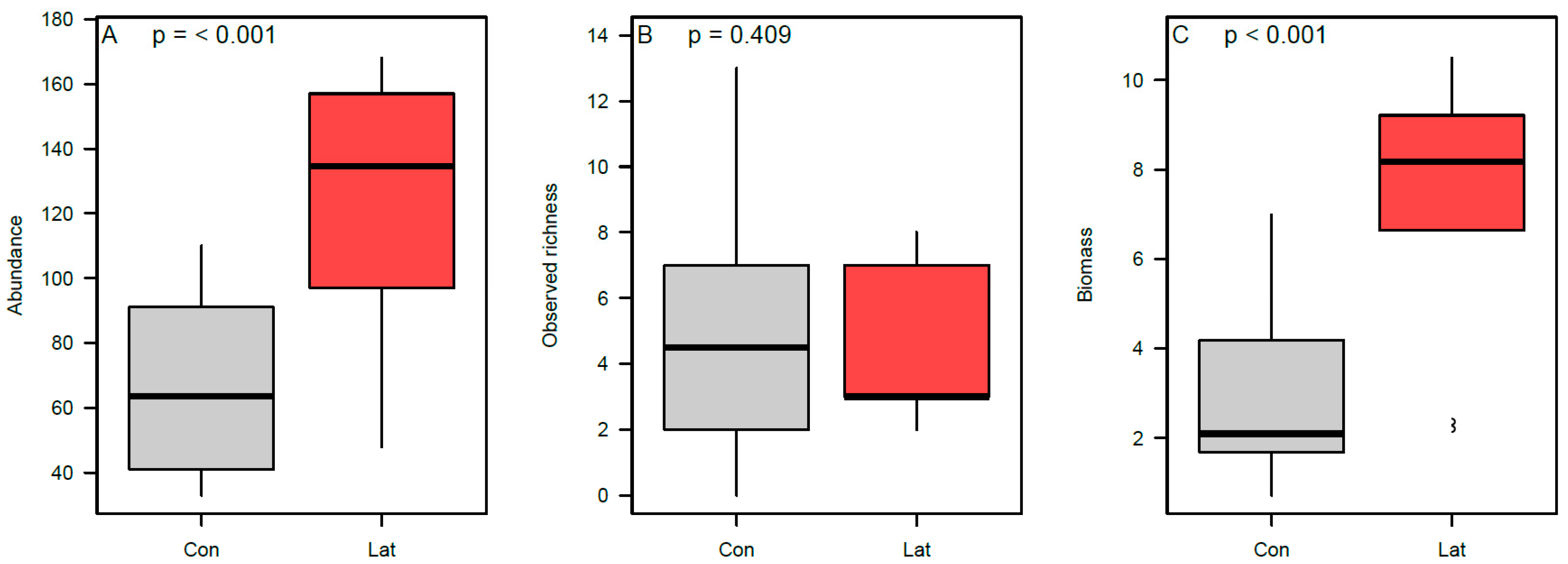
3.2. Terrestrial Vertebrate Activity Around Latrines
3.3. Dung Beetle Activity Within Latrine Sites
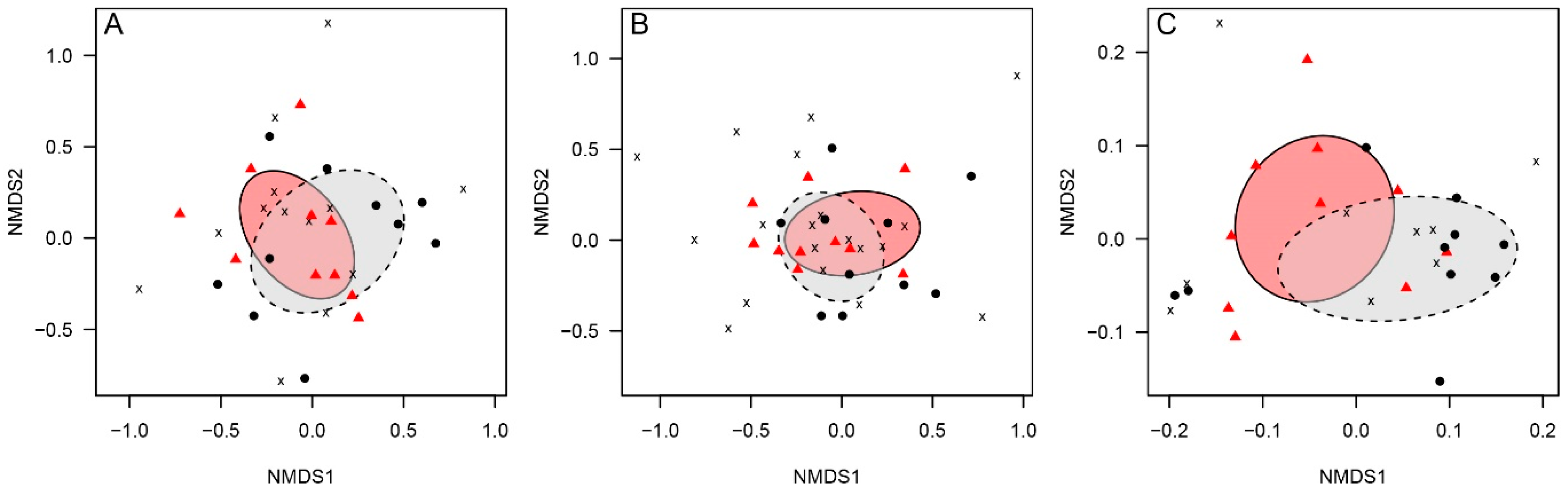
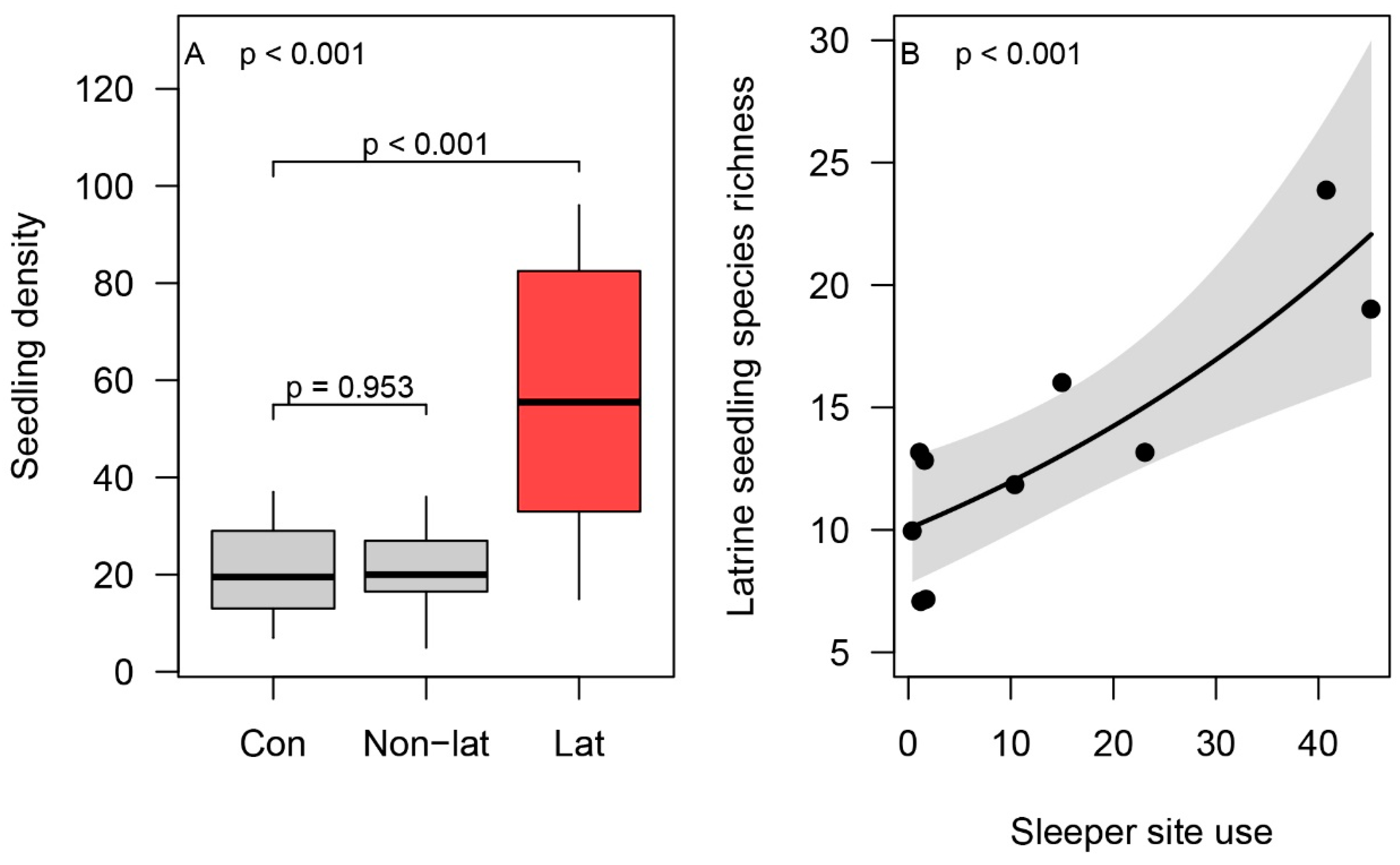
3.4. Latrine Floristic Composition and Soil Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peres, C.A.; Emilio, T.; Schietti, J.; Desmoulière, S.J.M.; Levi, T.; Thaise, E.; Schietti, J.; Desmoulieres, S.J.M.; Levi, T. Dispersal limitation induces long-term biomass collapse in overhunted Amazonian forests. Proc. Natl. Acad. Sci. USA 2016, 113, 892–897. [Google Scholar] [CrossRef]
- Brodie, J.F. How Monkeys Sequester Carbon. Trends Ecol. Evol. 2016, 31, 414–416. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Bello, C.; Galetti, M.; Pizo, M.A.; Magnago, L.F.S.; Rocha, M.F.; Lima, R.A.F.; Peres, C.A.; Ovaskainen, O.; Jordano, P. Defaunation affects carbon storage in tropical forests. Sci. Adv. 2015, 1, e1501105. [Google Scholar] [CrossRef]
- Link, A.; Di Fiore, A. Seed dispersal by spider monkeys and its importance in the maintenance of neotropical rain-forest diversity. J. Trop. Ecol. 2006, 22, 235–246. [Google Scholar] [CrossRef]
- González-Zamora, A.; Arroyo-Rodríguez, V.; Escobar, F.; Oyama, K.; Aureli, F.; Stoner, K.E. Sleeping-tree fidelity of the spider monkey shapes community-level seed-rain patterns in continuous and fragmented rain forests. J. Trop. Ecol. 2015, 31, 305–313. [Google Scholar] [CrossRef]
- Buxton, A.P. Further Observations of the Night-Resting Habits of Monkeys in a Small Area on the Edge of the Semliki Forest, Uganda. J. Anim. Ecol. 1951, 20, 31–32. [Google Scholar] [CrossRef]
- Chapman, C.A. Spider monkey sleeping sites: Use and availability. Am. J. Primatol. 1989, 18, 53–60. [Google Scholar] [CrossRef]
- Dew, J.L. Spider Monkeys as Seed Dispersers. In Spider Monkeys: Behavior, Ecology and Evolution of the Genus Ateles; Campbell, C.J., Ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 9780521867504. [Google Scholar]
- Hartwell, K.S.; Notman, H.; Bonenfant, C.; Pavelka, M.S.M. Assessing the Occurrence of Sexual Segregation in Spider Monkeys (Ateles geoffroyi yucatanensis), Its Mechanisms and Function. Int. J. Primatol. 2014, 35, 425–444. [Google Scholar] [CrossRef]
- González-Zamora, A.; Arroyo-Rodríguez, V.; Oyama, K.; Sork, V.; Chapman, C.A.; Stoner, K.E. Sleeping Sites and Latrines of Spider Monkeys in Continuous and Fragmented Rainforests: Implications for Seed Dispersal and Forest Regeneration. PLoS ONE 2012, 7, e46852. [Google Scholar] [CrossRef]
- Chapman, C.A.; Chapman, L.J.; McLaughlin, R.L. Multiple central place foraging by spider monkeys: Travel consequences of using many sleeping sites. Oecologia 1989, 79, 506–511. [Google Scholar] [CrossRef]
- Russo, S.E.; Augspurger, C.K. Aggregated seed dispersal by spider monkeys limits recruitment to clumped patterns in Virola calophylla. Ecol. Lett. 2004, 7, 1058–1067. [Google Scholar] [CrossRef]
- Whitworth, A.; Beirne, C.; Huarcaya, R.P.; Whittaker, L.; Rojas, S.J.S.; Tobler, M.W.; Macleod, R. Human disturbance impacts on rainforest mammals are most notable in the canopy, especially for larger-bodied species. Divers. Distrib. 2019, 25, 1166–1178. [Google Scholar] [CrossRef]
- Gregory, T.; Carrasco-Rueda, F.; Alonso, A.; Kolowski, J.; Deichmann, J.L. Natural canopy bridges effectively mitigate tropical forest fragmentation for arboreal mammals. Nature 2017, 7, 3892. [Google Scholar] [CrossRef]
- Bowler, M.T.; Tobler, M.W.; Endress, B.A.; Gilmore, M.P.; Anderson, M.J. Estimating mammalian species richness and occupancy in tropical forest canopies with arboreal camera traps. Remote Sens. Ecol. Conserv. 2017, 3, 146–157. [Google Scholar] [CrossRef]
- Whitworth, A.; Braunholtz, L.D.; Huarcaya, R.P.; Macleod, R.; Beirne, C. Out on a limb: Arboreal camera traps as an emerging methodology for inventorying elusive rainforest mammals. Trop. Conserv. Sci. 2016, 9, 675–698. [Google Scholar] [CrossRef]
- Oliveira-Santos, L.G.R.; Tortato, M.A.; Graipel, M.E. Activity pattern of Atlantic Forest small arboreal mammals as revealed by camera traps. J. Trop. Ecol. 2008, 24, 563–567. [Google Scholar] [CrossRef]
- Tan, C.L.; Yang, Y.; Niu, K. Into the night: Camera traps reveal nocturnal activity in a presumptive diurnal primate, Rhinopithecus brelichi. Primates 2013, 54, 1–6. [Google Scholar] [CrossRef]
- Fragoso, J.M.V.; Kirsten, M.S.; Correa, J.A. Long-Distance Seed Dispersal by Tapirs Increases Seed Survival and Aggregates Tropical Trees. Ecology 2003, 84, 1998–2006. [Google Scholar] [CrossRef]
- Bravo, S.P. The Impact of Seed Dispersal by Black and Gold Howler Monkeys on Forest Regeneration. Ecol. Res. 2012, 27, 311–321. [Google Scholar] [CrossRef]
- Camargo-Sanabria, A.A.; Mendoza, E. Interactions between terrestrial mammals and the fruits of two neotropical rainforest tree species. Acta Oecol. 2016, 73, 45–52. [Google Scholar] [CrossRef]
- Bogoni, J.A.; da Silva, P.G.; Peres, C.A. Co-declining mammal-dung beetle faunas throughout the Atlantic Forest biome of South America. Ecography 2019, 42, 1803–1818. [Google Scholar] [CrossRef]
- Culot, L.; Huynen, M.C.; Heymann, E.W. Primates and Dung Beetles: Two Dispersers Are Better than One in Secondary Forest. Int. J. Primatol. 2018, 39, 397–414. [Google Scholar] [CrossRef]
- Lawson, C.R.; Mann, D.J.; Lewis, O.T. Dung Beetles Reduce Clustering of Tropical Tree Seedlings. Biotropica 2012, 44, 271–275. [Google Scholar] [CrossRef]
- Lambert, J.E.; Chapman, C.A. The Fate of Primate-dispersed Seeds: Deposition Pattern, Dispersal Distance and Implications for Conservation. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.-M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CABI Publishing: Wallingford, UK, 2005; Volume 42, p. 137. ISBN 0851998062. [Google Scholar]
- Andresen, E.; Levey, D.J. Effects of dung and seed size on secondary dispersal, seed predation, and seedling establishment of rain forest trees. Oecologia 2004, 139, 45–54. [Google Scholar] [CrossRef]
- Urrea-Galeano, L.A.; Andresen, E.; Coates, R.; Mora Ardila, F.; Ibarra-Manríquez, G. Dung beetle activity affects rain forest seed bank dynamics and seedling establishment. Biotropica 2019, 51, 186–195. [Google Scholar] [CrossRef]
- Griffiths, H.M.; Bardgett, R.D.; Louzada, J.; Barlow, J. The value of trophic interactions for ecosystem function: Dung beetle communities influence seed burial and seedling recruitment in tropical forests. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161634. [Google Scholar] [CrossRef]
- Holdridge, L.R. Life zone ecology. In Life Zone Ecology; Tropical Science Center: San Jose, CA, USA, 1967. [Google Scholar]
- Weghorst, J.A. High population density of black-handed spider monkeys (Ateles geoffroyi) in Costa Rican lowland wet forest. Primates 2007, 48, 108–116. [Google Scholar] [CrossRef]
- Herwitz, S.R. Regeneration of Selected Tropical Tree Species in Corcovado National Park, Costa Rica; University of California Press: California, CA, USA, 1981; Volume 24, ISBN 9780520096318. [Google Scholar]
- Boza, M.A. Costa Rica National Parks; Editorial Heliconia: San José, CA, USA, 1988. [Google Scholar]
- Hartshorn, G.S. Wildlands conservation in Central America. In Tropical rain forest: ecology and management; Sutton, S.L., Whitmore, T.C., Chadwick, A.C., Eds.; Blackwell: Oxford, UK, 1983; pp. 423–443. [Google Scholar]
- Whitworth, A.; Beirne, C.; Flatt, E.; Huarcaya, R.P.; Diaz, J.C.C.; Forsyth, A.; Molnár, P.K.; Soto, J.S.V. Secondary forest is utilized by Great Curassows (Crax rubra) and Great Tinamous (Tinamus major) in the absence of hunting. Condor 2018, 120, 852–862. [Google Scholar] [CrossRef]
- Meek, P.D.; Ballard, G.; Claridge, A.; Kays, R.; Moseby, K.; O’Brien, T.; O’Connell, A.; Sanderson, J.; Swann, D.E.; Tobler, M.; et al. Recommended guiding principles for reporting on camera trapping research. Biodivers. Conserv. 2014, 23, 2321–2343. [Google Scholar] [CrossRef]
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- Pillco Huarcaya, R.; Beirne, C.; Serrano Rojas, S.J.; Whitworth, A. Camera trapping reveals a diverse and unique high-elevation mammal community under threat. Oryx 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.H.; Forsyth, A. Trap spacing and transect design for dung beetle biodiversity studies. Biotropica 2005, 37, 322–325. [Google Scholar] [CrossRef]
- Marsh, C.J.; Louzada, J.; Beiroz, W.; Ewers, R.M. Optimising Bait for Pitfall Trapping of Amazonian Dung Beetles (Coleoptera: Scarabaeinae). PLoS ONE 2013, 8, e73147. [Google Scholar] [CrossRef] [Green Version]
- Howden, H.F.; Nealis, V.G. Effects of clearing in a tropical rain forest on the composition of the coprophagous scarab beetle fauna (Coleoptera). Biotropica 1975, 7, 77–83. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.I.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef]
- R. Core. Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying levels of animal activity using camera trap data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, A.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; Hara, R.B.O.; Simpson, G.L.; Solymos, P.; et al. The Vegan Package. Community Ecol. Packag. 2007, 10, 631–637. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Bates, D. Linear mixed model implementation in lme4. Manuscr. Univ. Wis. 2007, 15, 1–32. [Google Scholar]
- Galvis, N.; Link, A.; Di Fiore, A. A novel use of camera traps to study demography and life history in wild animals: A case study of spider monkeys (Ateles belzebuth). Int. J. Primatol. 2014, 35, 908–918. [Google Scholar] [CrossRef]
- Scherbaum, C.; Estrada, A. Selectivity in feeding preferences and ranging patterns in spider monkeys Ateles geoffroyi yucatanensis of northeastern Yucatan peninsula, Mexico. Curr. Zool. 2013, 59, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Velázquez-Vázquez, G.; Reyna-Hurtado, R.; Arroyo-Rodríguez, V.; Calmé, S.; Léger-Dalcourt, M.; Navarrete, D.A. Sleeping Sites of Spider Monkeys (Ateles geoffroyi) in Logged and Unlogged Tropical Forests. Int. J. Primatol. 2015, 36, 1154–1171. [Google Scholar] [CrossRef]
- González-Zamora, A.; Arroyo-Rodríguez, V.; Escobar, F.; Rös, M.; Oyama, K.; Ibarra-Manríquez, G.; Stoner, K.E.; Chapman, C.A. Contagious deposition of seeds in spider monkeys’ sleeping trees limits effective seed dispersal in fragmented landscapes. PLoS ONE 2014, 9, e89346. [Google Scholar] [CrossRef] [Green Version]
- Busia, L.; Schaffner, C.M.; Aureli, F. Relationship quality affects fission decisions in wild spider monkeys (Ateles geoffroyi). Ethology 2017, 123, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Levi, T.; Shepard, G.H.; Ohl-Schacherer, J.; Peres, C.A.; Yu, D.W. Modelling the long-term sustainability of indigenous hunting in Manu National Park, Peru: Landscape-scale management implications for Amazonia. J. Appl. Ecol. 2009, 46, 804–814. [Google Scholar] [CrossRef]
- González-Zamora, A.; Arroyo-Rodríguez, V.; Chaves, Ó.M.; Sánchez-López, S.; Stoner, K.E.; Riba-Hernández, P. Diet of spider monkeys (Ateles geoffroyi) in mesoamerica: Current knowledge and future directions. Am. J. Primatol. 2009, 71, 8–20. [Google Scholar] [CrossRef]
- Pouvelle, S.; Jouard, S.; Feer, F.; Tully, T.; Ponge, J.F. The latrine effect: Impact of howler monkeys on the distribution of small seeds in a tropical rain-forest soil. J. Trop. Ecol. 2009, 25, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Aquino, R.; Encarnación, F. Characteristics and use of sleeping sites in Aotus (Cebidae: Primates) in the Amazon lowlands of Peru. Am. J. Primatol. 1986, 11, 319–331. [Google Scholar] [CrossRef]
- Fan, P.F.; Jiang, X.L. Sleeping sites, sleeping trees, and sleep-related behaviors of black crested gibbons (Nomascus concolor jingdongensis) at Mt. Wuliang, Central Yunnan, China. Am. J. Primatol. Off. J. Am. Soc. Primatol. 2008, 70, 153–160. [Google Scholar] [CrossRef]
- José-Domínguez, J.M.; Asensio, N.; García, C.J.G.; Huynen, M.C.; Savini, T. Exploring the multiple functions of sleeping sites in northern pigtailed macaques (Macaca leonina). Int. J. Primatol. 2015, 36, 948–966. [Google Scholar] [CrossRef]
- Dröscher, I.; Kappeler, P.M. Maintenance of familiarity and social bonding via communal latrine use in a solitary primate (Lepilemur leucopus). Behav. Ecol. Sociobiol. 2014, 68, 2043–2058. [Google Scholar] [CrossRef] [Green Version]
- Irwin, M.T.; Samonds, K.E.; Raharison, J.L.; Wright, P.C. Lemur latrines: Observations of latrine behavior in wild primates and possible ecological significance. J. Mammal. 2004, 85, 420–427. [Google Scholar] [CrossRef]
- Wallace, R.B. Diurnal Activity Budgets of black spider monkeys, Ateles chamek, in a Southern Amazonian Tropical Forest. Neotrop. Primates 2001, 9, 101–107. [Google Scholar]
- Ahumada, J.A. Grooming Behavior of Spider Monkeys (Ateles). Int. J. Primatol. 1992, 13, 33–49. [Google Scholar] [CrossRef]
- Munõz-Delgado, J.; Sánchez-Ferrer, J.C.; Pérez-Galicia, S.; Canales-Espinosa, D.; Erkert, H.G. Effects of housing conditions and season on the activity rhythm of spider monkeys (Ateles geoffroyi) kept under natural conditions within their distributional range in Central Mexico. Chronobiol. Int. 2014, 31, 983–995. [Google Scholar] [CrossRef]
- Muñoz-Delgado, J.; Corsi-Cabrera, M.; Canales-Espinosa, D.; Santillán-Doherty, A.M.; Erkert, H.G. Astronomical and meteorological parameters and rest-activity rhythm in the Spider monkey Ateles geoffroyi. Physiol. Behav. 2004, 83, 107–117. [Google Scholar] [CrossRef]
- Anderson, J.R. Sleep, sleeping sites, and sleep-related activities: Awakening to their significance. Am. J. Primatol. 1998, 46, 63–75. [Google Scholar] [CrossRef]
- Nichols, E.; Spector, S.; Louzada, J.; Larsen, T.; Amezquita, S.; Favila, M.E. Ecological functions and ecosystem services provided by Scarabaeinae dung beetles. Biol. Conserv. 2008, 141, 1461–1474. [Google Scholar] [CrossRef]
- Feeley, K. The role of clumped defecation in the spatial distribution of soil nutrients and the availability of nutrients for plant uptake. J. Trop. Ecol. 2005, 21, 99–102. [Google Scholar] [CrossRef]
- Dos Santos Neves, N.; Feer, F.; Salmon, S.; CHateil, C.; Ponge, J.F. The impact of red howler monkey latrines on the distribution of main nutrients and on topsoil profiles in a tropical rain forest. Austral Ecol. 2010, 35, 549–559. [Google Scholar] [CrossRef]
- Watmough, S.A.; Eimers, M.C.; Dillon, P.J. Manganese cycling in central Ontario forests: Response to soil acidification. Appl. Geochem. 2007, 22, 1241–1247. [Google Scholar] [CrossRef]
- Altrichter, M.; Carrillo, E.; Saenz, J.; Fuller Todd, K. White-lipped peccary (Tayassu pecari, Artiodactyla: Tayassuidae) diet and fruit availability in a Costa Rican rain forest. Rev. Biol. Trop. 2001, 49, 1183–1192. [Google Scholar]
- Spaan, D.; Burke, C.; McAree, O.; Aureli, F.; Rangel-Rivera, C.E.; Hutschenreiter, A.; Longmore, S.N.; McWhirter, P.R.; Wich, S.A. Thermal Infrared Imaging from Drones Offers a Major Advance for Spider Monkey Surveys. Drones 2019, 3, 34. [Google Scholar] [CrossRef] [Green Version]
- Chanthorn, W.; Hartig, F.; Brockelman, W.Y.; Srisang, W.; Nathalang, A.; Santon, J. Defaunation of large-bodied frugivores reduces carbon storage in a tropical forest of Southeast Asia. Sci. Rep. 2019, 9, 10015. [Google Scholar] [CrossRef]
- Wallace, R.B. Factors influencing spider monkey habitat use and ranging patterns. In Spider Monkeys; Cambridge University Press: Cambridge, UK, 2010; pp. 138–154. [Google Scholar]
- Chapman, C.A.; Bonnell, T.R.; Gogarten, J.F.; Lambert, J.E.; Omeja, P.A.; Twinomugisha, D.; Wasserman, M.D.; Rothman, J.M. Are Primates Ecosystem Engineers? Int. J. Primatol. 2013, 34, 1–14. [Google Scholar] [CrossRef]
- Lawton, J.H.; Jones, C.G. Linking Species and Ecosystems: Organisms as Ecosystem Engineers. In Link Species & Ecosystems; Springer: Boston, MA, USA, 1995; pp. 141–150. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitworth, A.; Whittaker, L.; Pillco Huarcaya, R.; Flatt, E.; Morales, M.L.; Connor, D.; Priego, M.G.; Forsyth, A.; Beirne, C. Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity. Animals 2019, 9, 1052. https://doi.org/10.3390/ani9121052
Whitworth A, Whittaker L, Pillco Huarcaya R, Flatt E, Morales ML, Connor D, Priego MG, Forsyth A, Beirne C. Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity. Animals. 2019; 9(12):1052. https://doi.org/10.3390/ani9121052
Chicago/Turabian StyleWhitworth, Andrew, Lawrence Whittaker, Ruthmery Pillco Huarcaya, Eleanor Flatt, Marvin Lopez Morales, Danielle Connor, Marina Garrido Priego, Adrian Forsyth, and Chris Beirne. 2019. "Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity" Animals 9, no. 12: 1052. https://doi.org/10.3390/ani9121052
APA StyleWhitworth, A., Whittaker, L., Pillco Huarcaya, R., Flatt, E., Morales, M. L., Connor, D., Priego, M. G., Forsyth, A., & Beirne, C. (2019). Spider Monkeys Rule the Roost: Ateline Sleeping Sites Influence Rainforest Heterogeneity. Animals, 9(12), 1052. https://doi.org/10.3390/ani9121052