
Pleistocene Caves of Eastern Sicily Coast: Exceptional Archives to Reconstruct the History of the Island’s Biota


 ,
, 
Abstract
:1. Introduction
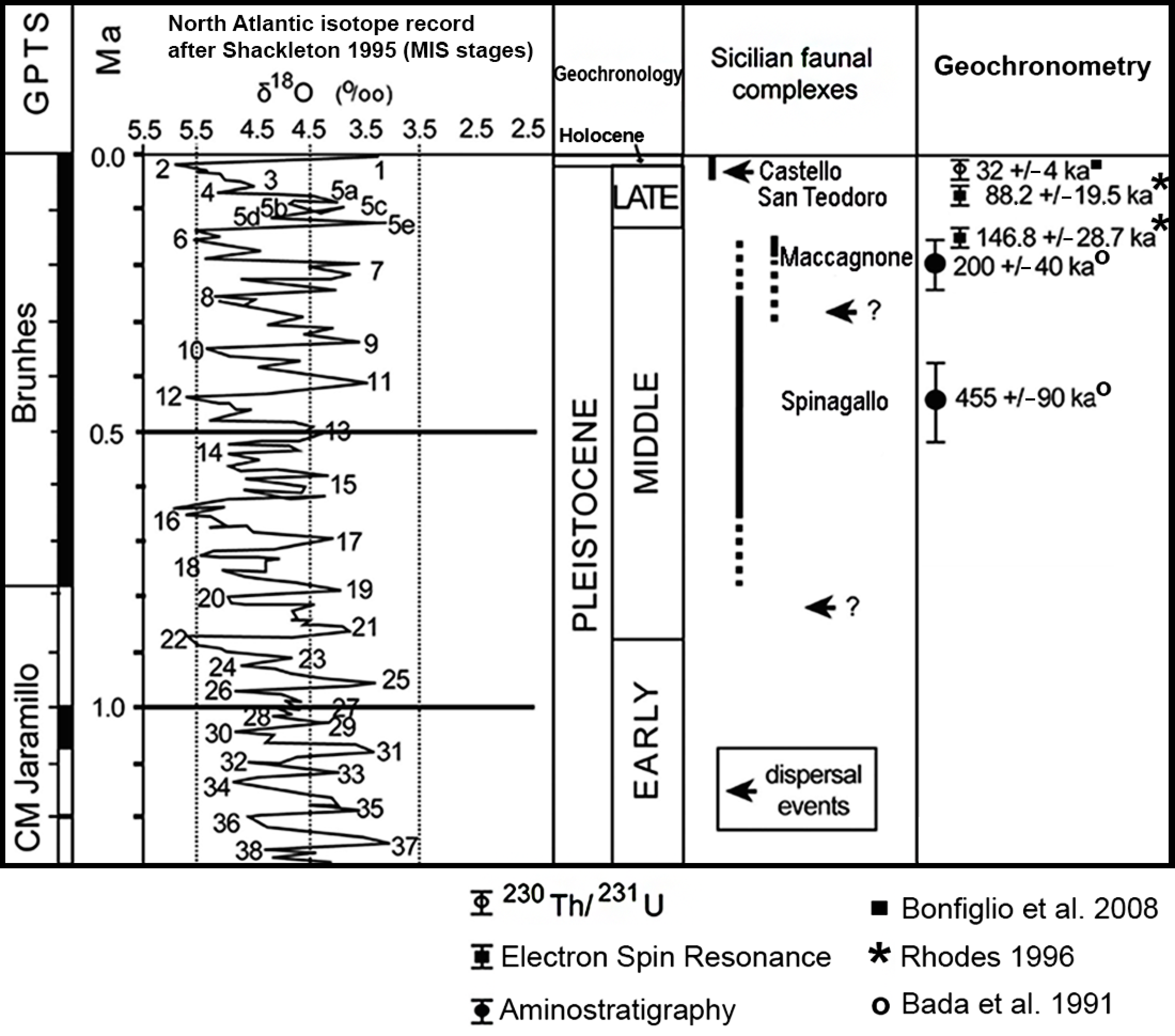
2. Geographical and Geochronological Setting
3. Description of the Caves
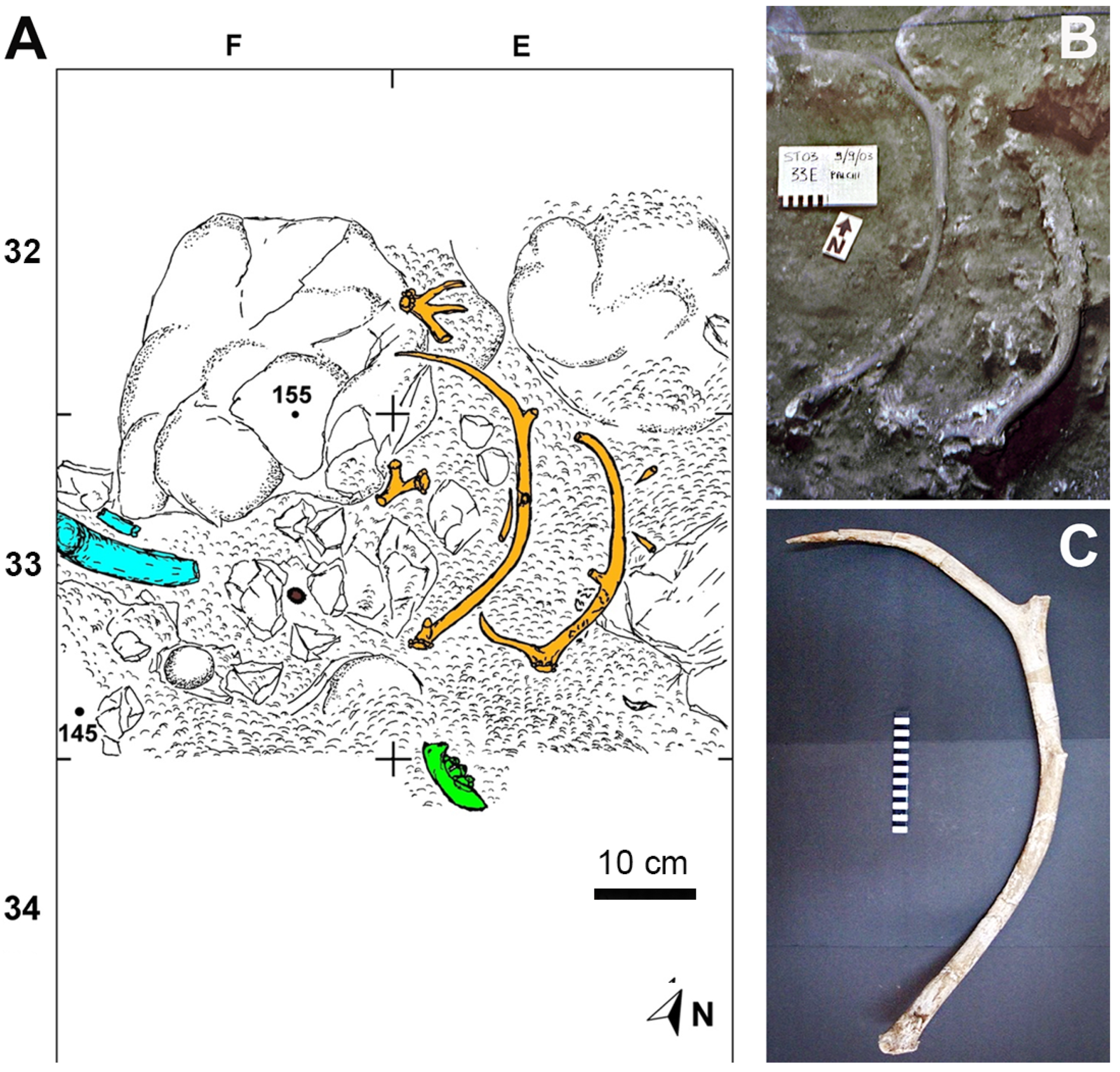
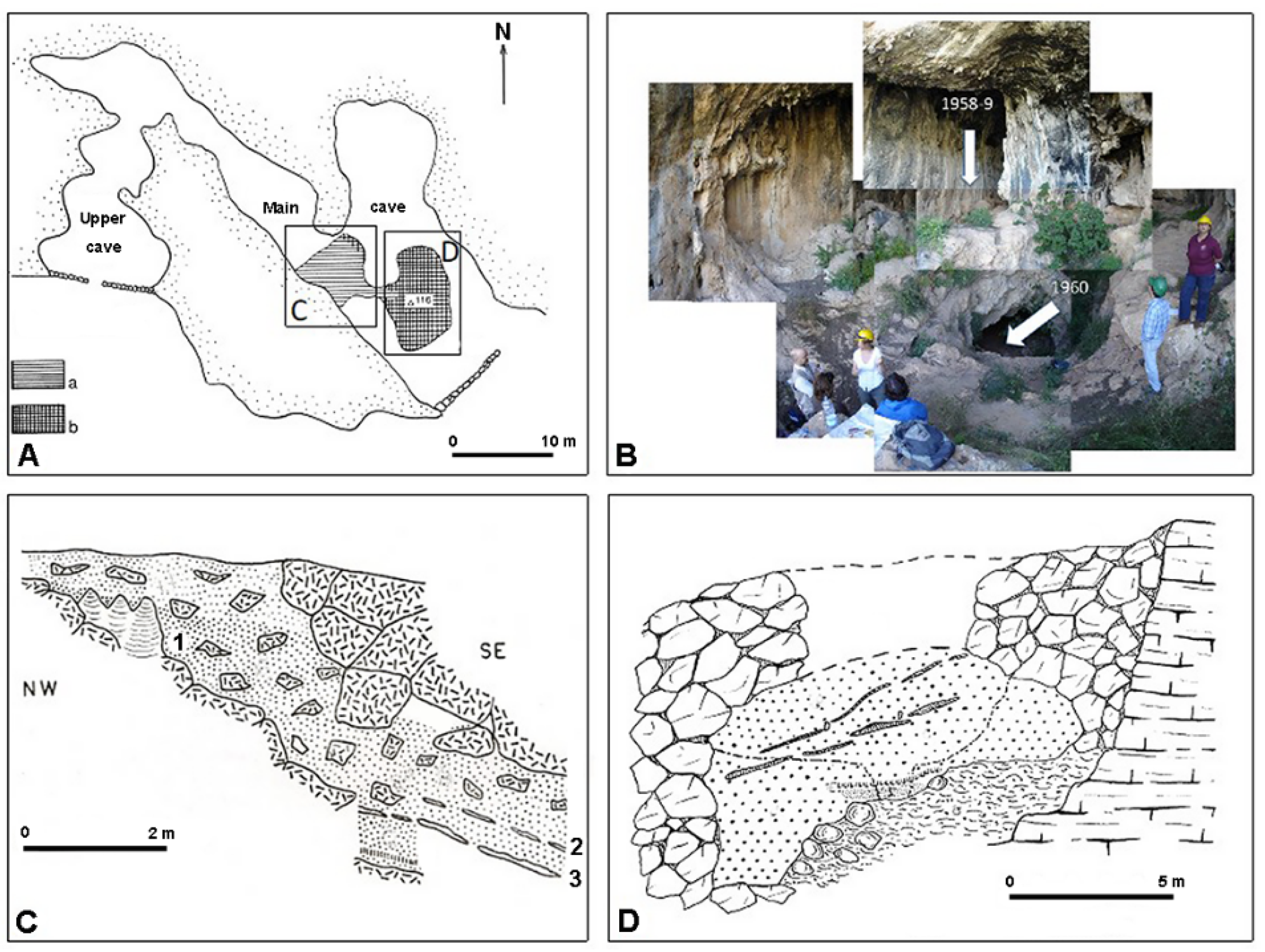
3.1. San Teodoro Cave
3.2. Donnavilla Cave
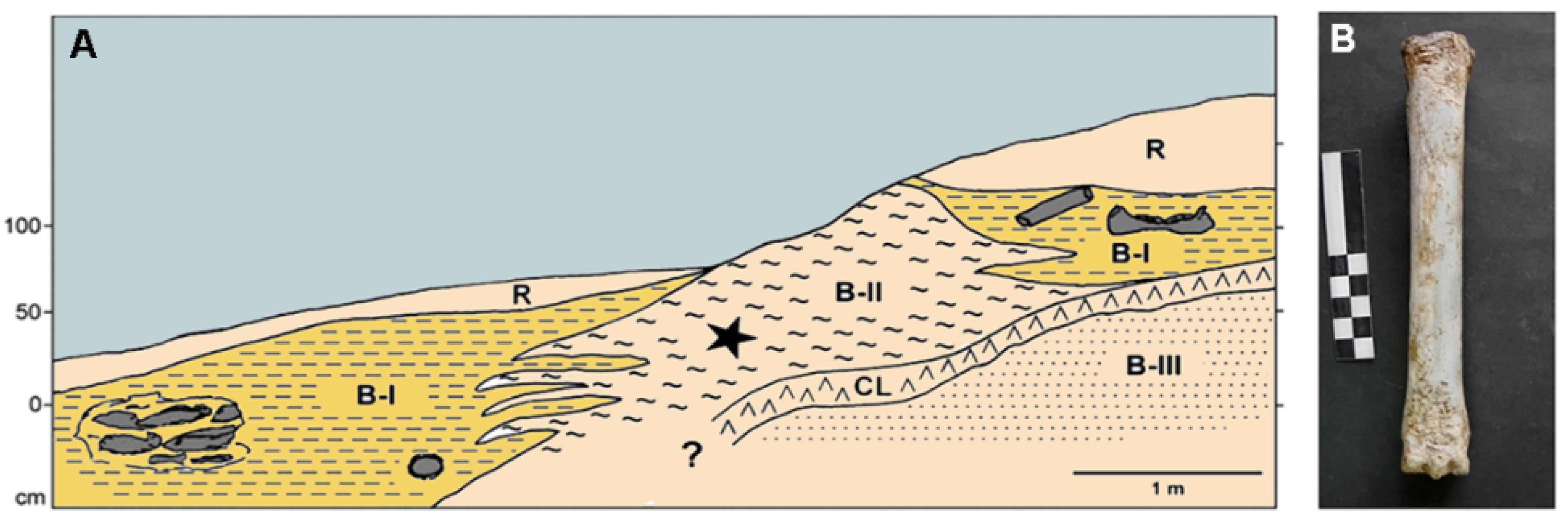
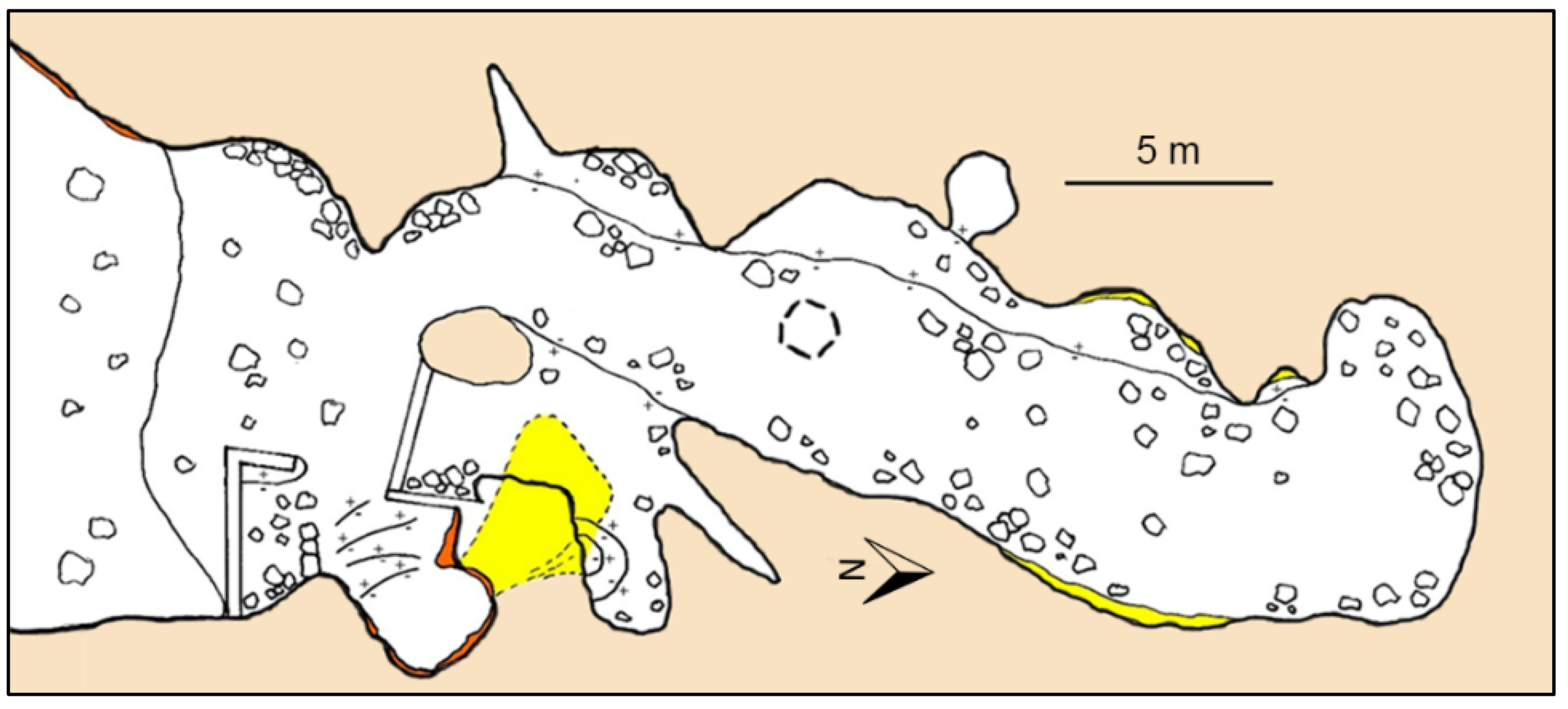
3.3. Fulco Cave
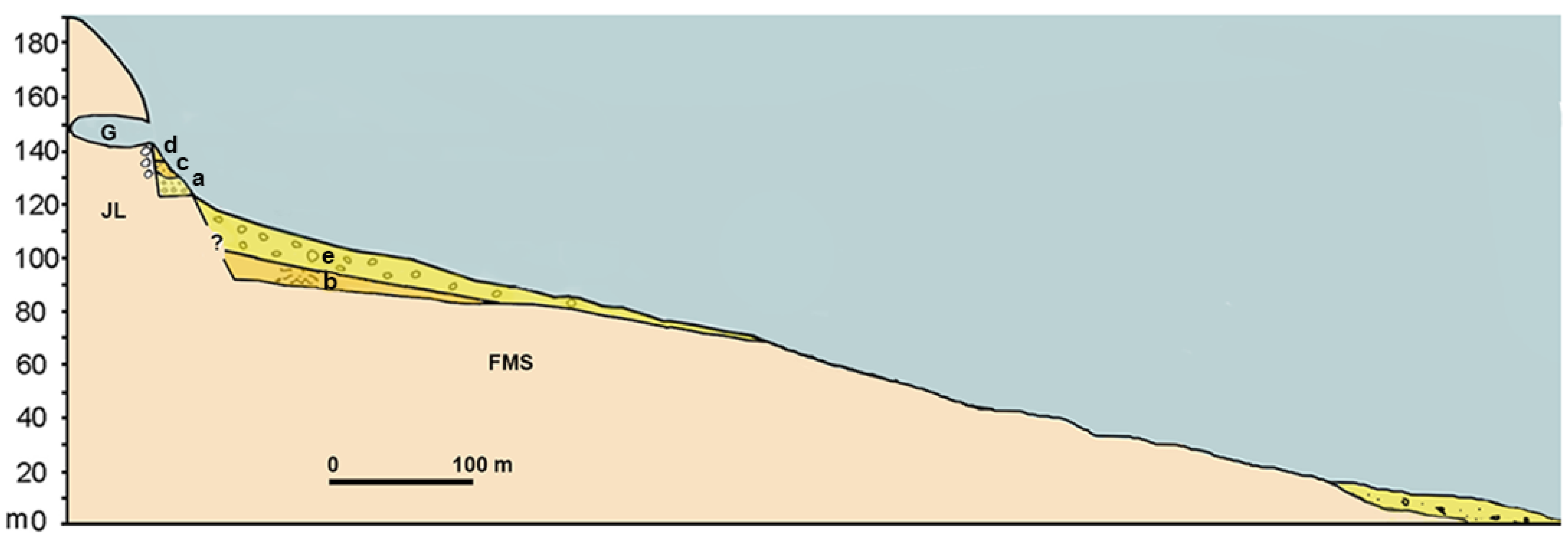
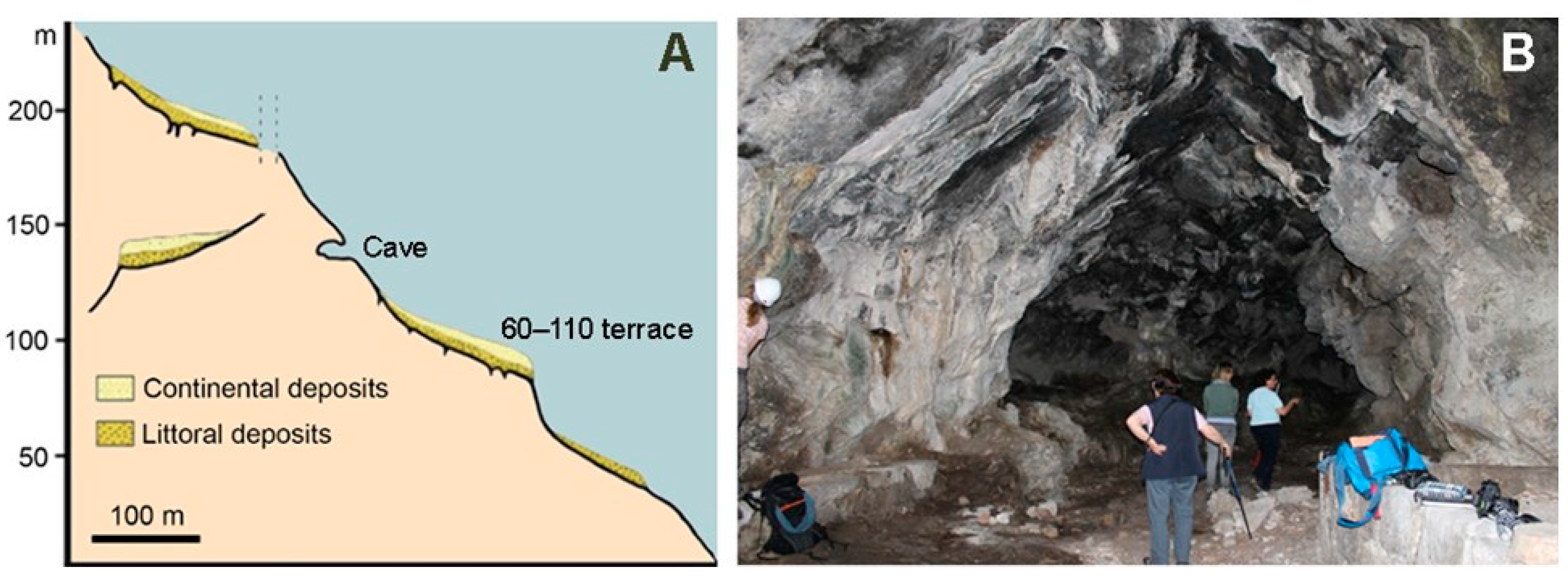
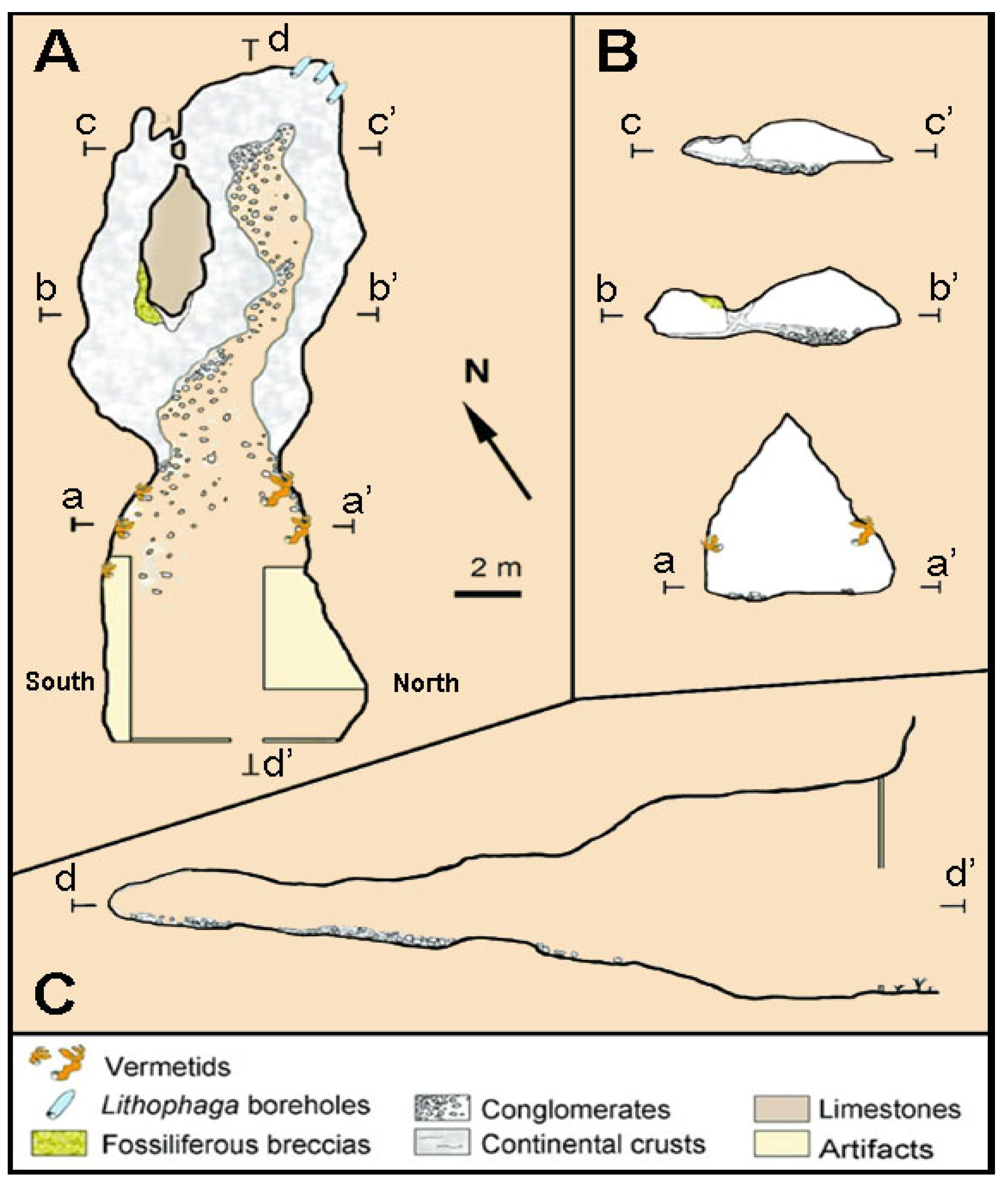
3.4. Taormina Cave
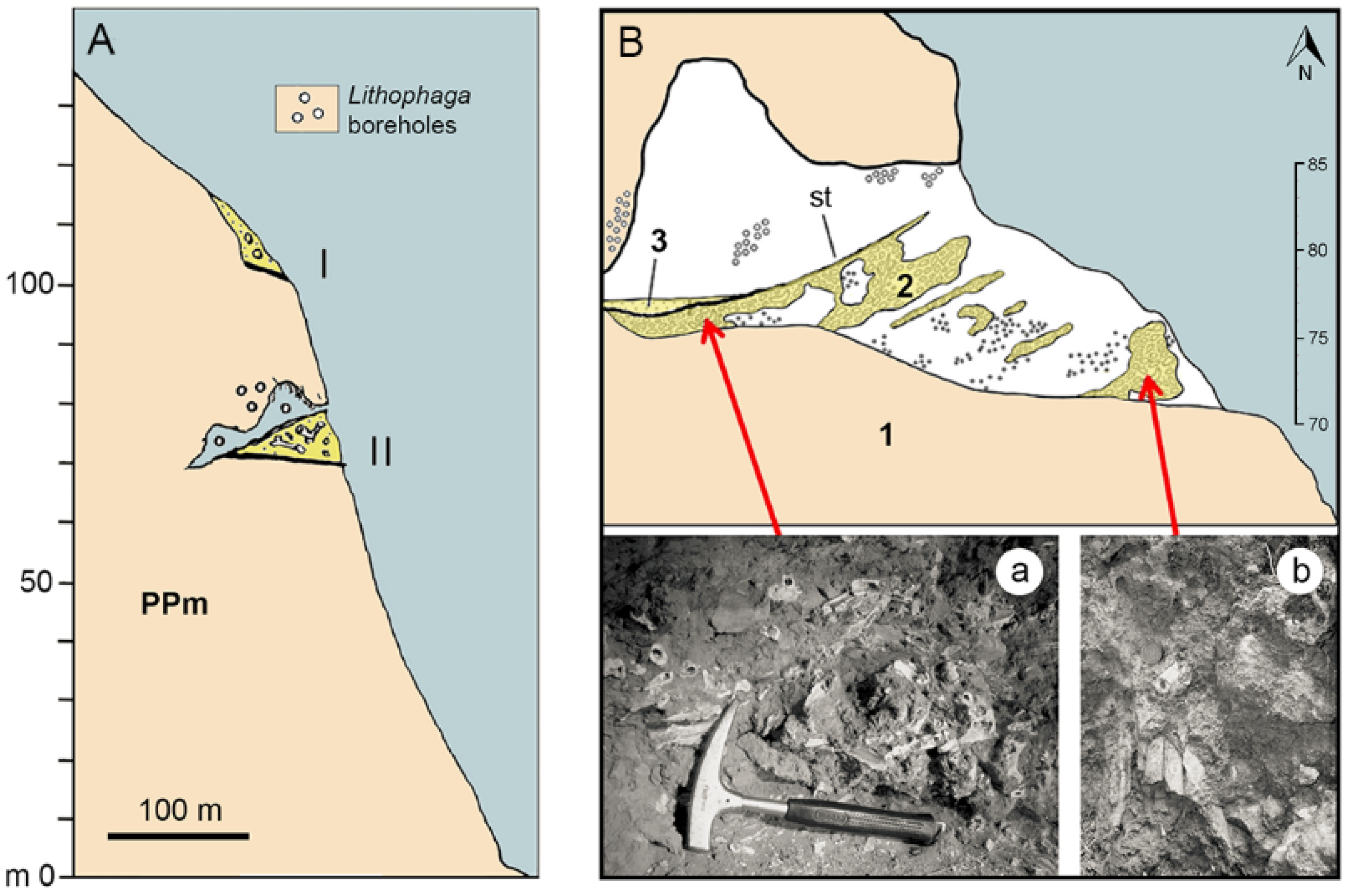
3.5. Tremilia Cave
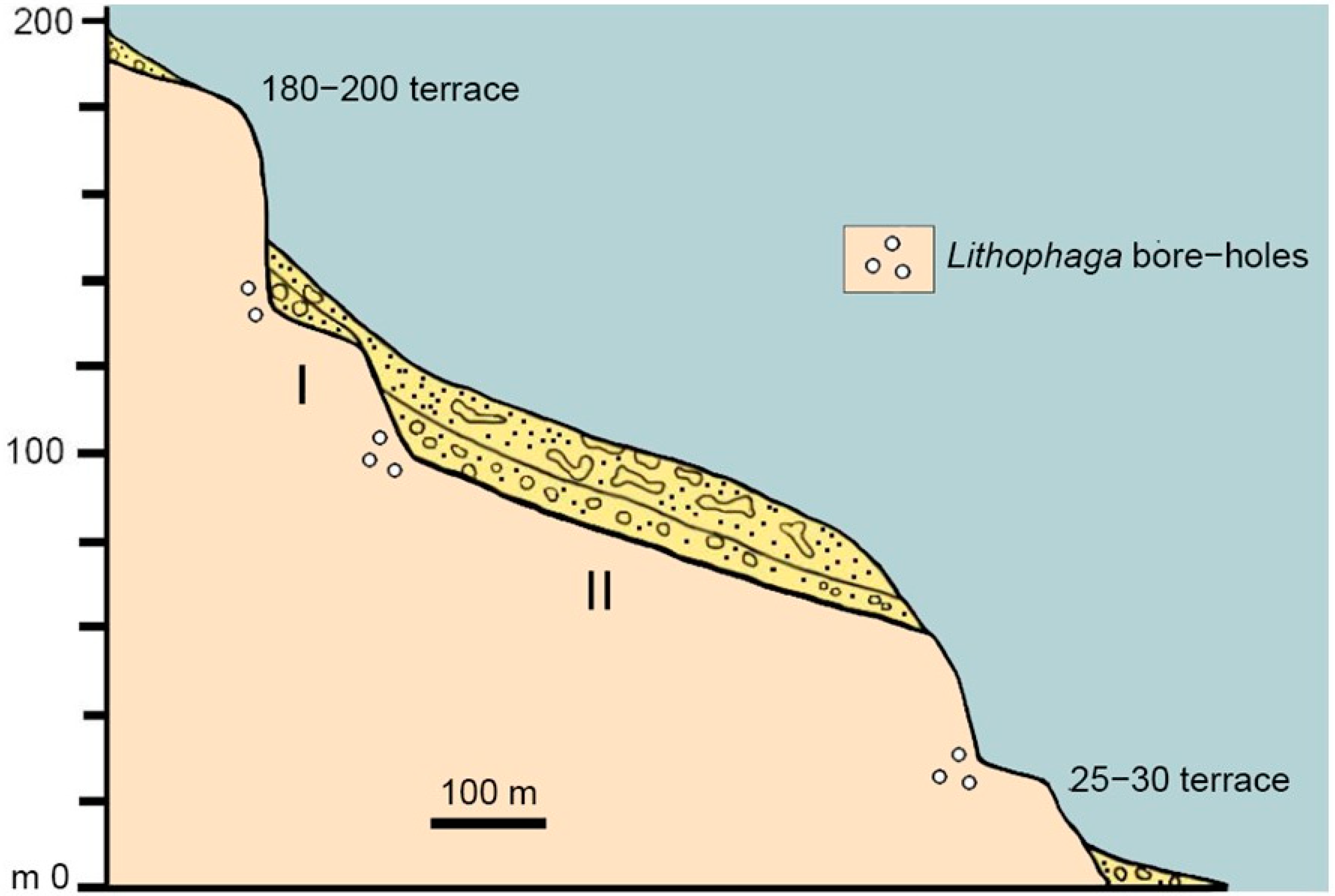
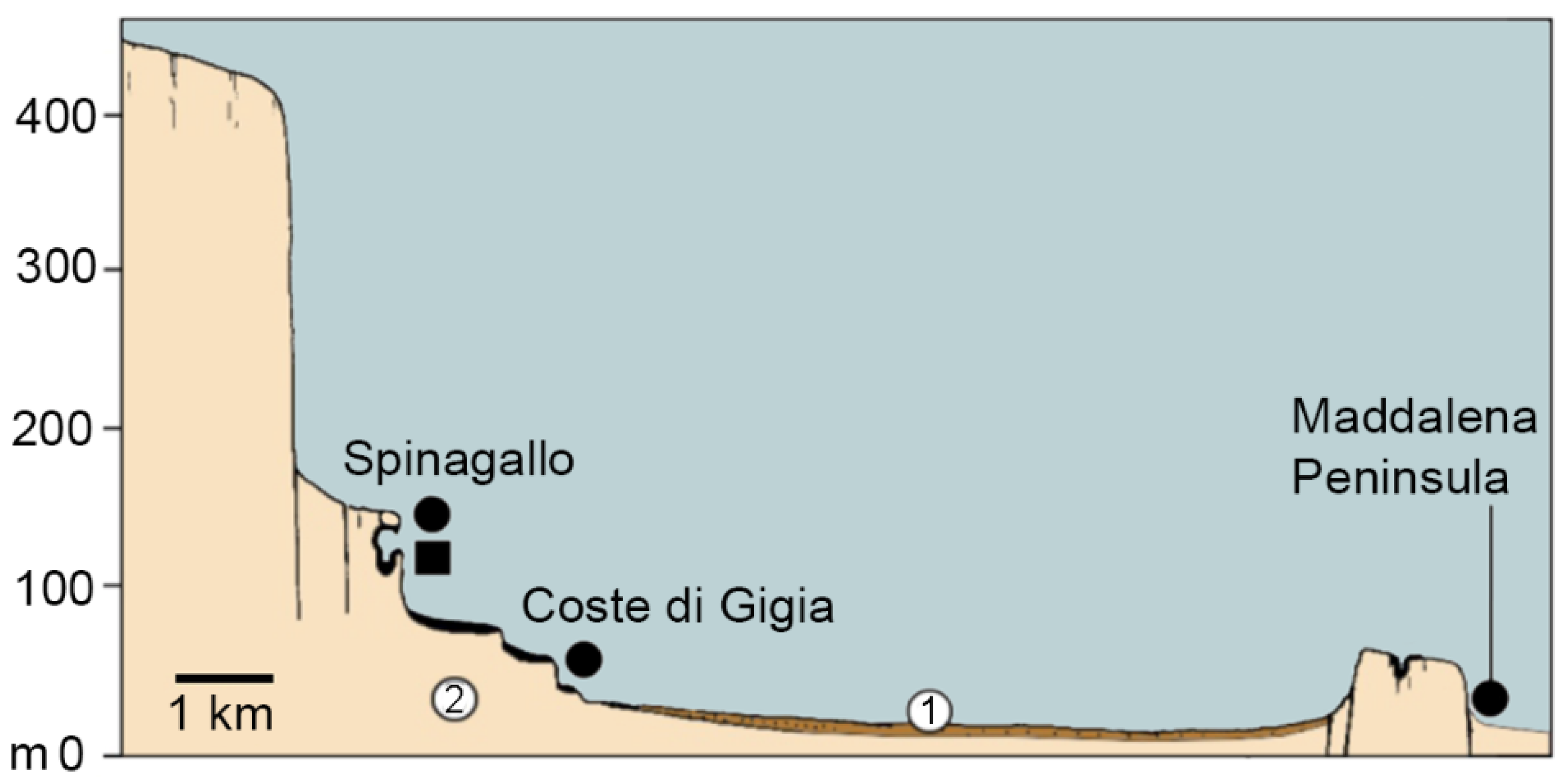
3.6. Spinagallo Cave and Hyblean Plateau Fissure Fills
4. Discussion
4.1. Palaeoenvironmental Meaning
4.2. Documented Time Spans
4.3. Distinctive Features of the Caves
4.4. Palaeoecological Inferences
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antonioli, F.; Kershaw, S.; Rendac, P.; Rust, D.; Belluomini, G.; Cerasoli, M.; Radtke, U.; Silenzi, S. Elevation of the last interglacial highstand in Sicily (Italy): A benchmark of coastal tectonics. Quat. Internat. 2006, 145–146, 3–18. [Google Scholar] [CrossRef]
- Carobene, L. Terrazzi marini, eustatismo e neotettonica. Geogr. Fis. Dinam. Quatern. 1980, 3, 35–41. [Google Scholar]
- Belluomini, G.; Bada, J.L. Isoleucine epimerization ages of the dwarf elephants of Sicily. Geology 1985, 13, 451–452. [Google Scholar] [CrossRef]
- Bada, J.L.; Belluomini, G.; Bonfiglio, L.; Branca, M.; Burgio, E.; Delitala, L. Isoleucine Epimerization ages of Quaternary Mammals of Sicily. Ital. J. Quat. Sci. 1991, 4, 5–11. [Google Scholar]
- Rhodes, E.J. ESR Dating of Tooth Enamel. In Le Ossa dei Giganti. Lo scavo Paleontologico di Contrada Fusco; Basile, B., Chilardi, S., Eds.; Arnaldo Lombardi: Siracusa, Italy, 1996; pp. 39–44. [Google Scholar]
- Bonfiglio, L.; Mangano, G.; Marra, A.C.; Masini, F.; Pavia, M.; Petruso, D. Pleistocene Calabrian and Sicilian bioprovinces. Geobios. Spec. Mem. 2002, 24, 29–39. [Google Scholar] [CrossRef]
- Carbone, S.; Di Geronimo, I.; Grasso, M.; Iozzia, S.; Lentini, F. I terrazzi marini quaternari dell’area iblea (Sicilia sud-orientale). Contributi conclusivi progetto finalizzato. In Geodinamica. Contributi Conclusivi Progetto Finalizzato Geodinamica; C.N.R.: Rome, Italy, 1982; 35p. [Google Scholar]
- Di Grande, A.; Raimondo, W. Linee di costa plio-pleistoceniche e schema litostratigrafico del Quaternario siracusano. Geol. Rom. 1982, 21, 279–309. [Google Scholar]
- Bianca, M. Quaternary normal faulting in southeastern Sicily (Italy): A seismic source for the 1693 large earthquake. Geophys. J. Int. 1998, 139, 370–394. [Google Scholar] [CrossRef] [Green Version]
- Monaco, C.; Barreca, G.; Di Stefano, A.; Ristuccia, G. Quaternary marine terraces and fault activity in the northern sector of the Messina Straits (southern Italy). GNGTS 2014, 1, 85–90. [Google Scholar] [CrossRef]
- Marziano, C.; Chilardi, S. Contribution to knowledge of the Pleistocene mammal-bearing deposits of Siracusa Southeastern Sicily). In Biosphere to Lithosphere, Proceedings of the 9th ICAZ Conference, Durham, UK, 23–28 Auguts 2002; O’Connor, T., Ed.; Oxbow Books: Oxford, UK, 2005; pp. 94–109. [Google Scholar]
- Guzzardi, L. Ricerche archeologiche nel Siracusano. Kokalos 1993, 39–40, 1299–1303. [Google Scholar]
- Bonfiglio, L.; Marra, A.C.; Masini, F. The contribution of Quaternary vertebrates to the paleoenvironmental and paleoclimatic reconstructions in Sicily. Spec. Pubbl. Geol. Soc. Lond. 2000, 181, 169–182. [Google Scholar]
- Marra, A.C. Evolution of Endemic Species, Ecological Interactions and Geographical Changes in an Insular Environment: A Case Study of Quaternary Mammals of Sicily (Italy, EU). Geosciences 2013, 3, 114–139. [Google Scholar] [CrossRef] [Green Version]
- Hugonie, G. Mouvementes tectoniques et variation de la morphogenese au Quaternaire en Sicilie septentrionale. Rev. Geol. Dynam. Geogr. Phys. 1982, 23, 3–14. [Google Scholar]
- Hugonie, G. L’évolution Géomorphologique de la Sicile Septentrionale. Ph.D. Thesis, University Paris Sorbonne, Paris, France, 1979. Chapter 2. pp. 1–949. [Google Scholar]
- Catalano, S.; De Guidi, G. Late Quaternary uplift of northeastern Sicily: Relation with the active normal faulting. J. Geodyn. 2003, 36, 445–467. [Google Scholar] [CrossRef]
- Hearty, P.J.; Bonfiglio, L.; Violanti, D.; Szazo, B.J. Age of late Quaternary marine deposits of Southern Italy determined by aminostratigraphy, faunal correlation and uranium-series dating. Riv. Ital. Paleont. Strat. 1986, 92, 149–164. [Google Scholar]
- Robillard, D. Les depots quaternaires du versant tyrrhénien de la Sicile (secteur d’Aquedolci-Capo d’Orlando) stratigraphie et tectonique. D.E.A; Lille University of Science and Technology: Villeneuve-d’Ascq, France, 1975; p. 143. [Google Scholar]
- Miyauchi, T.; Dai Pra, G.; Sylos Labini, S. Geochronology of Pleistocene marine terraces and regional tectonics in the Tyrrhenian coast of southern Calabria, Italy. Il Quaternario 1994, 7, 17–34. [Google Scholar]
- Di Geronimo, I.; Ghisetti, F.; Grasso, M.; Lentini, F.; Scamarda, G.; Vezzani, L. Dati Preliminari Sulla Neotettonica della Sicilia sud-Orientale. Fogli 273 (Caltagirone), 274 (Siracusa), 275 (Scoglitti), 277 (Noto); Nuovi Contributi alla Carta Neotettonica d’Italia, C.N.R. Pubblicazioni Progetto Finalizzato Geodinamica n. 356; C.N.R.: Rome, Italy, 1980; pp. 747–773. [Google Scholar]
- Di Grande, A.; Scamarda, G. Segnalazione di livelli a Strombus bubonius Lamarck nei dintorni di Augusta (Siracusa). Boll. Acc. Gioenia Sci. Nat. Catania 1973, 11, 157–172. [Google Scholar]
- Bonfiglio, L. Il Tirreniano di Bovetto e Ravagnese presso Reggio Calabria. Quaternaria 1972, 16, 137–148. [Google Scholar]
- Bonfiglio, L.; Violanti, D. Prima segnalazione di Tirreniano ed evoluzione Pleistocenica del Capo Peloro (Sicilia Nord-Orientale). Geogr. Fis. Dinam. Quatern. 1983, 6, 3–15. [Google Scholar]
- Meschis, M.; Roberts, G.P.; Robertson, J.; Mildon, Z.K.; Sahy, D.; Goswami, R.; Sgambato, C.; Faure Walker, J.; Michetti, A.M.; Iezzi, F. Out of Phase Uplift-Rate Changes During the Quaternary Reveal Normal Fault Interaction, Implied by Deformed Marine Palaeoshorelines, in Southern Italy. Available online: http://dx.doi.org/10.2139/ssrn.4016967 (accessed on 25 January 2022).
- Conti, M.A.; Di Geronimo, I.; Esu, D.; Grasso, M. Il Pleistocene in facies limnica di Vittoria (Sicilia meridionale). Geol. Romana 1979, 18, 93–104. [Google Scholar]
- Bonfiglio, L.; Insacco, G. Palaeoenvironmental, paleontologic and stratigraphic significance of vertebrate remains in Pleistocene limnic and alluvial deposits from South Eastern Sicily. Palaeogeogr. Palaeoclimatol. Paleoecol. 1992, 95, 195–208. [Google Scholar] [CrossRef]
- Bonfiglio, L. Faunal and Human Populations. In Early Societies in Sicily; Leighton, R., Ed.; University of London: London, UK, 1996; pp. 21–29. [Google Scholar]
- Antonioli, F.; Lo Presti, V.; Gasparo Morticelli, M.; Bonfiglio, L.; Mannino, A.M.; Palombo, M.R.; Sannino, G.; Ferranti, L.; Furlani, S.; Lambeck, K.; et al. Timing of the emergence of the Europe–Sicily bridge (40–17 cal ka BP) and its implications for the spread of modern humans. Geol. Soc. Lond. Spec. Publ. 2014, 411, 111–144. [Google Scholar] [CrossRef]
- Bonfiglio, L.; Esu, D.; Mangano, G.; Masini, F.; Petruso, D.; Soligo, M.; Tuccimei, P. Late Pleistocene vertebrate-bearing deposits at San Teodoro Cave (Northe-Eastern Sicily): Preliminary data on faunal diversification and chronology. Quat. Int. 2008, 190, 26–37. [Google Scholar] [CrossRef]
- Mangano, G.; Insacco, G.; Bonfiglio, L.; Mazza, P.P. New finds from San Teodoro Cave: An updating of the Middle Pleistocene fossil record from Acquedolci (north-eastern Sicily). Palaeobiodivers. Palaeoenviron. 2020, 100, 1065–1076. [Google Scholar] [CrossRef]
- Anca, F. Note sur deux nouvelles grottes ossifères. Bull. Soc. Géol.Fr. 1860, 17, 684–695. [Google Scholar]
- Graziosi, P.; Maviglia, C. La grotta di S. Teodoro (Messina). Riv. Sci. Preist. 1946, 1, 227–283. [Google Scholar]
- Graziosi, P. Gli uomini paleolitici della grotta di S. Teodoro (Messina). Riv. Sci. Preist. 1947, 2, 123–224. [Google Scholar]
- Sineo, L.; Bigazzi, R.; D’Amore, G.; Tartarelli, G.; Di Patti, C.; Berzero, C.; Caramella Crespi, V. I resti umani della Grotta di S. Teodoro (Messina): Datazione assoluta con il metodo della spettrometria gamma diretta (U/Pa). Antropo 2002, 2, 9–16. [Google Scholar]
- Garilli, V.; Vita, G.; Mulone, A.; Bonfiglio, L.; Sineo, L. From sepulchre to butchery-cooking: Facies analysis, taphonomy and stratigraphy of the Upper Palaeolithic post burial layer from the San Teodoro Cave (NE Sicily) reveal change in the use of the site . J. Archaeol. Sci. Rep. 2020, 30, 102191. [Google Scholar] [CrossRef]
- Bonfiglio, L. Prima campagna di scavo dei depositi a mammiferi pleistocenici dell’area della Grotta di S. Teodoro (Acquedolci, Messina, Sicilia). Geol. Romana 1983, 22, 271–285. [Google Scholar]
- Bonfiglio, L. Middle and Upper Pleistocene Mammal-bearing deposits in southeastern Sicily: New stratigraphical records from Coste di Gigia (Syracuse). Geobios 1992, 14, 189–199. [Google Scholar] [CrossRef]
- Mangano, G.; Bonfiglio, L. New stratigraphic and taphonomic data from the late Pleistocene deposits of the S. Teodoro Cave (North Eastern Sicily, Italy). Ann. Univ. St. Ferrara Museol. Sci. E Nat. 2005, 1, 89–97. [Google Scholar]
- Bonfiglio, L. Nuovi elementi faunistici e stratigrafici del Pleistocene superiore dei Nebrodi (Sicilia nord-orientale). Riv. Ital. Paleontol. Stratigr. 1987, 93, 145–164. [Google Scholar]
- Bonfiglio, L.; Mangano, G.; Marra, A.C.; Masini, F. A new late Pleistocene vertebrate faunal complex from Sicily (S. Teodoro Cave, North Eastern Sicily, Italy). Boll. Soc. Paleont. Ital. 2001, 40, 149–158. [Google Scholar]
- Esu, D.; Mangano, G.; Bonfiglio, L. The molluscan fauna from the Upper Pleistocene vertebrate-bearing deposits of S. Teodoro cave (North-Eastern Sicily). Riv. Ital. Paleont. Strat. 2007, 113, 127–138. [Google Scholar]
- Mangano, G. An exclusively hyena-collected bone assemblage in the Late Pleistocene of Sicily: Taphonomy and stratigraphic context of the large mammal remains from San Teodoro Cave (North-Eastern Sicily, Italy). J. Arch. Sci. 2011, 38, 3584–3595. [Google Scholar] [CrossRef]
- Herridge, V.L. Dwarf Elephants on Mediterranean Islands: A Natural Experiment in Parallel Evolution. Ph.D. Thesis, University College London, London, UK, 2010. [Google Scholar]
- Mangano, G.; Bonfiglio, L. First finding of a partially articulated elephant skeleton from a Late Pleistocene hyena den in Sicily (San Teodoro Cave, North Eastern Sicily, Italy). Quat. Int. 2012, 276–277, 53–60. [Google Scholar] [CrossRef]
- Bonfiglio, L.; Mangano, G.; Pino, P. The contribution of mammal-bearing deposits to timing Late Pleistocene tectonics of Cape Tindari (North Eastern Sicily). Riv. Ital. Paleont. Strat. 2010, 116, 103–118. [Google Scholar]
- Graziosi, P. Gli scavi dell’Istituto Italiano di Paleontologia Umana nella grotta di S. Teodoro (Messina), nota preliminare. Atti Soc. Tosc. Sci. Nat. Mem. 1943, 52, 82–99. [Google Scholar]
- Fabbri, P.F. Nuove determinazioni del sesso e della statura degli individui 1 e 4 del Paleolitico superiore della Grotta di San Teodoro. Riv. Sci. Preist. 1993, 45, 219–232. [Google Scholar]
- Palombo, M.R.; Antonioli, F.; Di Patti, C.; Lo Presti, V.; Scarborough, M. Was the dwarfed Palaeoloxodon from Favignana Island the last endemic Pleistocene elephant from the western Mediterranean islands? Hist. Biol. 2020, 33, 2116–2134. [Google Scholar] [CrossRef]
- Yll, R.; Carriòn, J.S.; Marra, A.C.; Bonfiglio, L. Vegetation reconstruction on the basis of pollen in Late Pleistocene hyena coprolites from San Teodoro Cave (Sicily, Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 237, 32–39. [Google Scholar] [CrossRef]
- Bonfiglio, L.; Riccobono, F. Il deposito a Hippopotamus sp. del Pleistocene superiore presso la grotta di S.Teodoro in Acquedolci (Messina, Sicilia),un progetto di Museo paleontologico in situ. Boll. Soc. Paleont. Ital. 1991, 29, 109–115. [Google Scholar]
- Malatesta, A. Osservazioni sul Pliocene e il Pleistocene della costa settentrionale della Sicilia tra Gualtieri Sicaminò e Gioiosa Marea. Boll. Serv. Geol. Ital. 1958, 79, 291–326. [Google Scholar]
- Gliozzi, E.; Malatesta, A. A megacerine in the Pleistocene of Sicily. Geol. Rom. 1984, 21, 311–389. [Google Scholar]
- Caloi, L. Resti di cervo e di ippopotamo quaternari nella grotta di Capo Tindari. Boll. Serv. Geol. Ital. 1973, 93, 227–249. [Google Scholar]
- Caloi, L.; Palombo, M.R. Hippopotamus pentlandi von Meyer di Capo Tindari (Messina). Geologica Romana 1984, 21, 390–395. [Google Scholar]
- Abbazzi, L.; Bonfiglio, L.; Marra, A.C.; Masini, F. A revision of medium and small sized deer from the Middle and Late Pleistocene of Calabria and Sicily. Boll. Soc. Paleont. Ital. 2001, 40, 115–126. [Google Scholar]
- Fois, E. La successione neogenica di Capo Milazzo (Sicilia nord-orientale). Riv. Ital. Paleont. Strat. 1989, 95, 397–440. [Google Scholar]
- Fois, E. Stratigraphy and palaeogeography of the Capo Milazzo area (NE Sicily, Italy): Clues to evolution of the southern margin of the Tyrrhenian Basin during the Neogene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 78, 87–107. [Google Scholar] [CrossRef]
- Rosso, A.; Sanfilippo, R.; Vertino, A.; Zibrowius, H. Hanging coral gardens of a Tyrrhenian submarine cave from Sicily. Boll. Soc. Paleont. Ital. 2017, 56, 1–12. [Google Scholar]
- Bonfiglio, L. Terrazzi marini e depositi continentali quaternari di Taormina (Sicilia). Quaternaria 1981, 23, 81–102. [Google Scholar]
- Rosso, A.; Sanfilippo, R.; Bonfiglio, L.; Richards, D.; Nita, D. Exceptional Pleistocene vermetid crusts preserved in a cave located 130 m a.s.l. near Taormina (NE Sicily). Boll. Soc. Paleont. Ital. 2018, 57, 133–144. [Google Scholar]
- Rosso, A.; Sanfilippo, R.; Di Geronimo, I.; Bonfiglio, L. Pleistocene occurrence of recently discovered cryptic vermetid species from the Mediterranean. Boll. Soc. Paleont. Ital. 2016, 55, 105–109. [Google Scholar]
- Lister, A.M. Ecological interactions of elephantids in Pleistocene Eurasia: Palaeoloxodon and Mammuthus. In Human Paleoecology in the Levantine Corridor; Goren-Inbar, N., Speth, J.D., Eds.; Oxbow: Oxford, UK, 2004; pp. 53–60. [Google Scholar]
- Ambrosetti, P. The Pleistocene dwarf elephants of Spinagallo (Siracusa, south-eastern Sicily). Geol. Romana 1968, 7, 277–398. [Google Scholar]
- Accordi, B.; Campisi, B.; Colacicchi, R. Scoperta di un giacimento a elefanti nani e ghiro gigante nella grotta Spinagallo (Siracusa). Atti Acc. Gioenia Sci. Nat. Catania 1959, 12, 167–182. [Google Scholar]
- Accordi, B.; Colacicchi, R. Excavation of pigmy elephants cave of Spinagallo (Siracusa). Geol. Romana 1962, 1, 217–230. [Google Scholar]
- Belluomini, G.; Bacchin, P. Datazione di ossa fossili di grotte italiane con il metodo della racemizzazione degli ammino-acidi e criterio di distinzione fra reperti esposti e non esposti al riscaldamento. Geol. Rom. 1980, 19, 171–180. [Google Scholar]
- Bonfiglio, L.; Insacco, G.; Marra, A.C.; Masini, F. Large and small mammals, amphibians, reptiles from a new Late Pleistocene fissure filling deposit of the Hyblean Plateau (South Eastern Sicily). Boll. Soc. Paleont. Ital. 1997, 35, 97–122. [Google Scholar]
- Di Geronimo, I.; Costa, B. Il Pleistocene di Monte dell’Apa (Gela). Riv. Ital. Paleont. Strat. 1978, 84, 1121–1158. [Google Scholar]
- Grasso, M.; Lentini, F. Sedimentary and tectonic evolution of the eastern Hyblean Plateau (Southeastern Sicily) during Late Cretaceous to Quaternary time. Palaeo 1982, 39, 261–280. [Google Scholar] [CrossRef]
- Carbone, S. I depositi pleistocenici del settore nord-orientale ibleo tra Agnone e Melilli (Sicilia SE): Relazione tra facies e lineamenti strutturali. Boll. Soc. Geol. It. 1985, 104, 405–420. [Google Scholar]
- Chilardi, S.; Gilotti, A. Stratigrafia e sedimentologia. In Le Ossa dei Giganti. Lo Scavo Paleontologico di Contrada Fusco; Basile, B., Chilardi, S., Eds.; Arnaldo Lombardi: Siracusa, Italy, 1996; pp. 27–34. [Google Scholar]
- Fabiani, R. Resti di Mammiferi del Terziario e del Quaternario di Ragusa in Sicilia. Rend. R. Acc. Naz. Lincei 1927, 6, 521–524. [Google Scholar]
- Fabiani, R. Aggiunte alla Conoscenza dei Mammiferi Fossili del Ragusano in Sicilia; Istituto Geologico della Regia Università: Palermo, Italy, 1928; 8p. [Google Scholar]
- Maugeri Patanè, G. Su alcune ossa fossili di mammiferi quaternari in Contrada Batteria presso Augusta (Sicilia). Atti Acc. Gioenia Sci. Nat. Catania 1932, 5, 1–12. [Google Scholar]
- Maugeri Patanè, G. Sopra un teschio di lupo fossile di Contrada Tabuna (Ragusa di Sicilia). Atti Acc. Gioenia Sci. Nat. Catania 1936, 6, 1–13. [Google Scholar]
- Bonfiglio, L.; Di Stefano, G.; Insacco, G.; Marra, A.C. New Pleistocene fissure filling deposits from the Hyblean Plateau (South Eastern Sicily). Riv. Ital. Paleont. Strat. 1992, 98, 523–540. [Google Scholar]
- Bonfiglio, L. Correlazioni tra depositi a mammiferi, depositi marini, linee di costa e terrazzi medio e tardo pleistocenici nella Sicilia orientale. Il Quat. 1991, 4, 205–216. [Google Scholar]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Caves | Elevation a.s.l. (m) | Middle Pleistocene | Tyrrhenian | Holocene | Age (ka) | Faunistic Features | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Marine Phase | Continental Phase | Marine Phase | Continental Phase | ||||||||
| Deposits/Encrustations | Lithophaga Borings | P. falconeri FC | Maccagnone FC | San Teodoro FC | Encrustations | Lithophaga Borings | Deposits | ||||
| San Teodoro | 140 | 200–11 | Hominids, hyenas, Equus hydruntinus | ||||||||
| Donnavilla | 73–85 | 200–40 | Lithophaga belt | ||||||||
| Fulco | 51 | 125 | Hanging corals with elongated corallites | ||||||||
| Taormina | 131 | older than 125–upper paleolithic | Vermetids with downward facing tubes | ||||||||
| Tremilia | 86 | 300–200 | Glycymeris bed | ||||||||
| Spinagallo | 120–130 | ?550–200 | Paleoloxodon falconeri | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonfiglio, L.; Rosso, A.; Herridge, V.; Insacco, G.; Reitano, A.; Minniti, G.; Mangano, G.; Sanfilippo, R. Pleistocene Caves of Eastern Sicily Coast: Exceptional Archives to Reconstruct the History of the Island’s Biota. Geosciences 2022, 12, 258. https://doi.org/10.3390/geosciences12070258
Bonfiglio L, Rosso A, Herridge V, Insacco G, Reitano A, Minniti G, Mangano G, Sanfilippo R. Pleistocene Caves of Eastern Sicily Coast: Exceptional Archives to Reconstruct the History of the Island’s Biota. Geosciences. 2022; 12(7):258. https://doi.org/10.3390/geosciences12070258
Chicago/Turabian StyleBonfiglio, Laura, Antonietta Rosso, Victoria Herridge, Gianni Insacco, Agatino Reitano, Gianmarco Minniti, Gabriella Mangano, and Rossana Sanfilippo. 2022. "Pleistocene Caves of Eastern Sicily Coast: Exceptional Archives to Reconstruct the History of the Island’s Biota" Geosciences 12, no. 7: 258. https://doi.org/10.3390/geosciences12070258
APA StyleBonfiglio, L., Rosso, A., Herridge, V., Insacco, G., Reitano, A., Minniti, G., Mangano, G., & Sanfilippo, R. (2022). Pleistocene Caves of Eastern Sicily Coast: Exceptional Archives to Reconstruct the History of the Island’s Biota. Geosciences, 12(7), 258. https://doi.org/10.3390/geosciences12070258