Phenological Model Intercomparison for Estimating Grapevine Budbreak Date (Vitis vinifera L.) in Europe

 ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
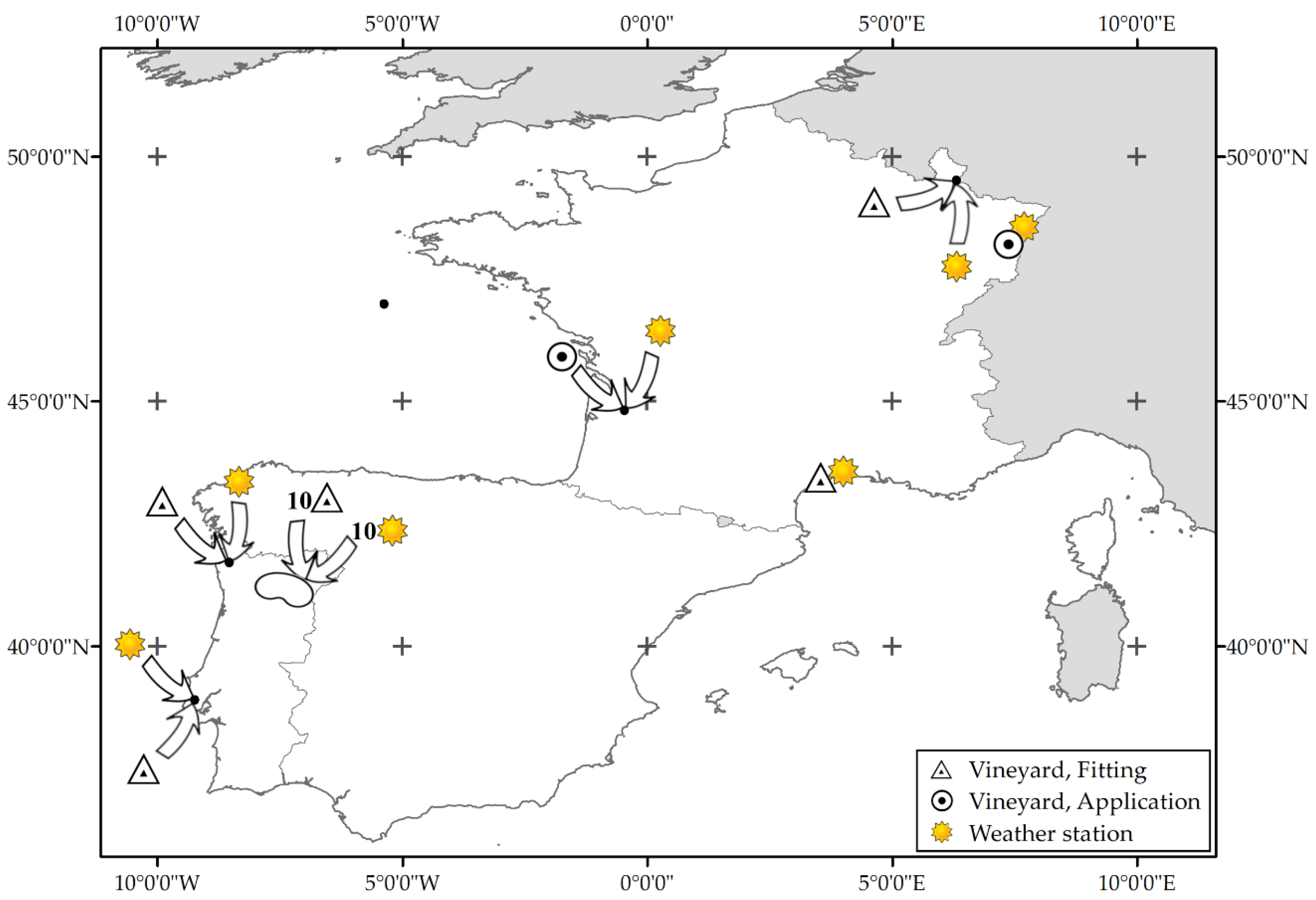
2.1. Observations
2.2. Phenological Models
2.2.1. GDD
2.2.2. WANG
2.2.3. UNIFORC
2.2.4. BRIN
2.2.5. UNICHILL
2.2.6. UNIFIED
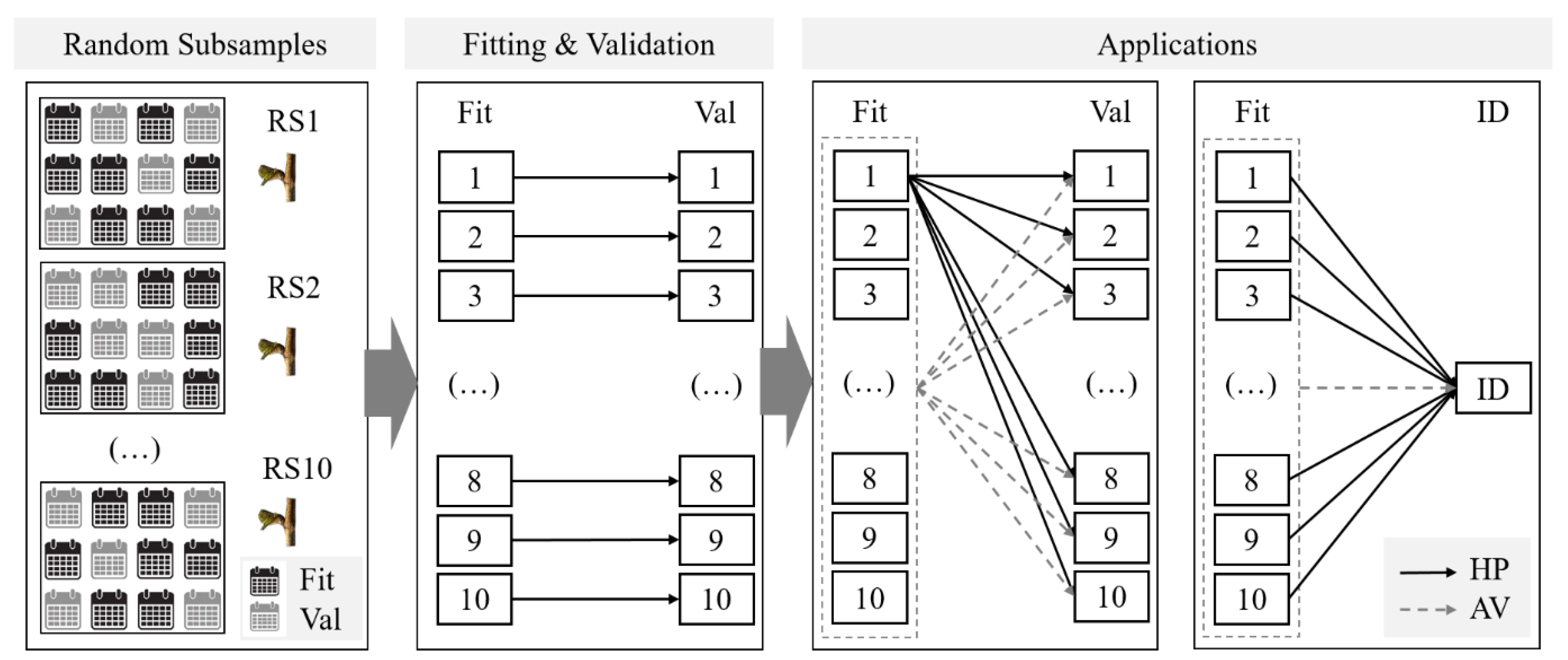
2.3. Model Fitting and Validation
2.3.1. Outliers Removal
2.3.2. Parameters’ Range and Starting Date
2.3.3. Model Fitting, Validation and Application
2.4. Statistical Analysis
3. Results
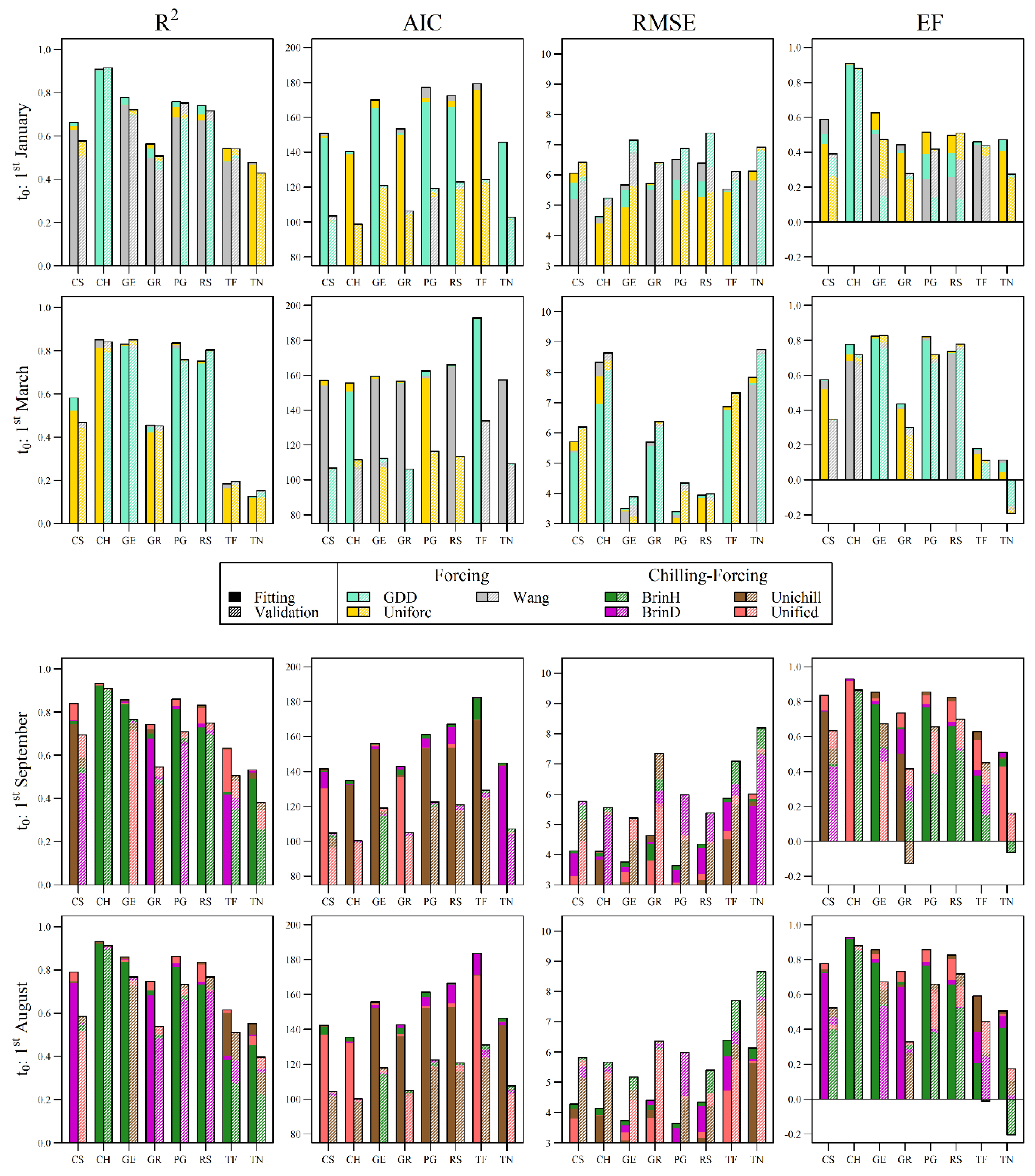
3.1. Overall Model Behavior in Fitting and Validation
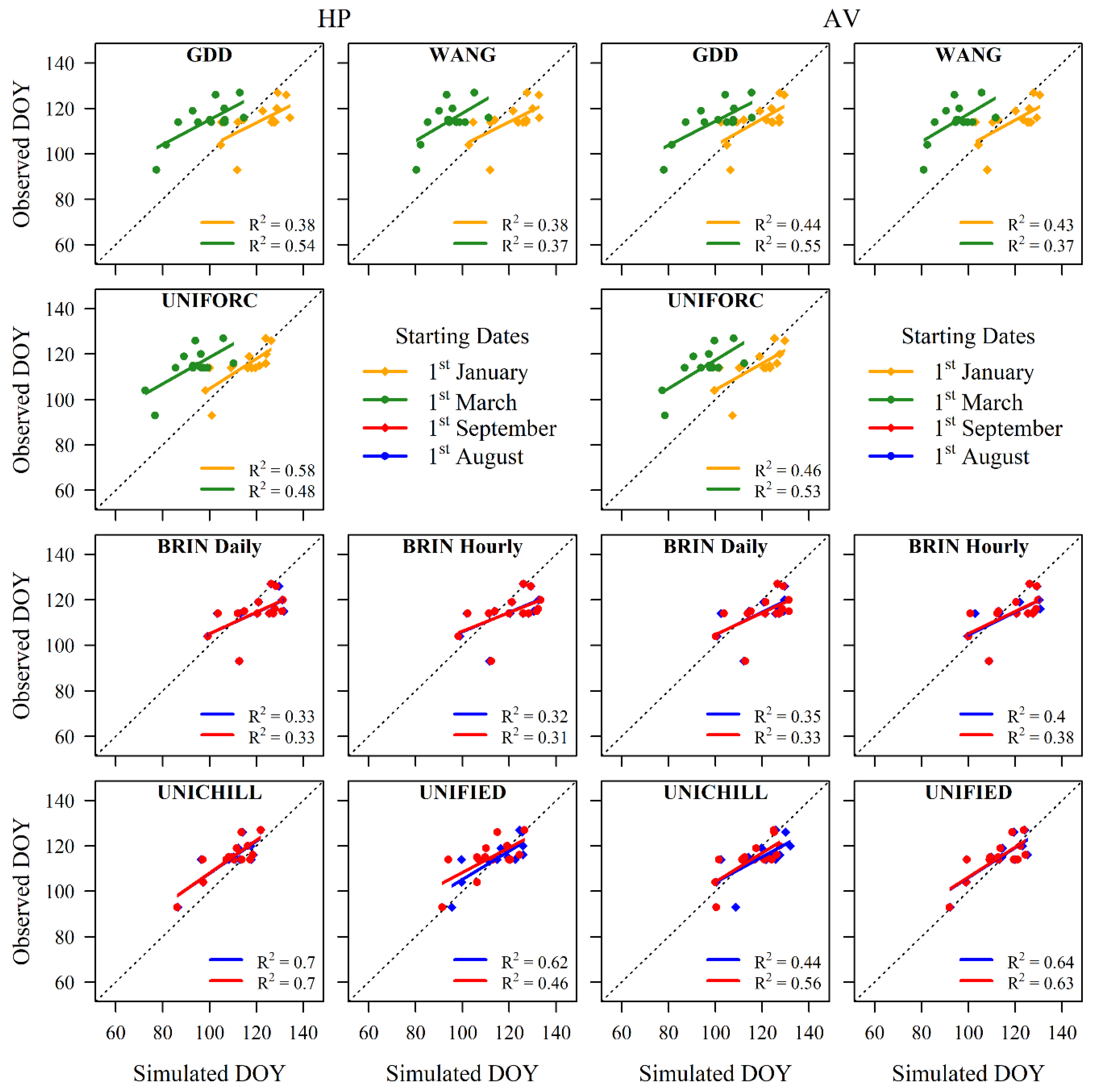
3.2. HP versus AV
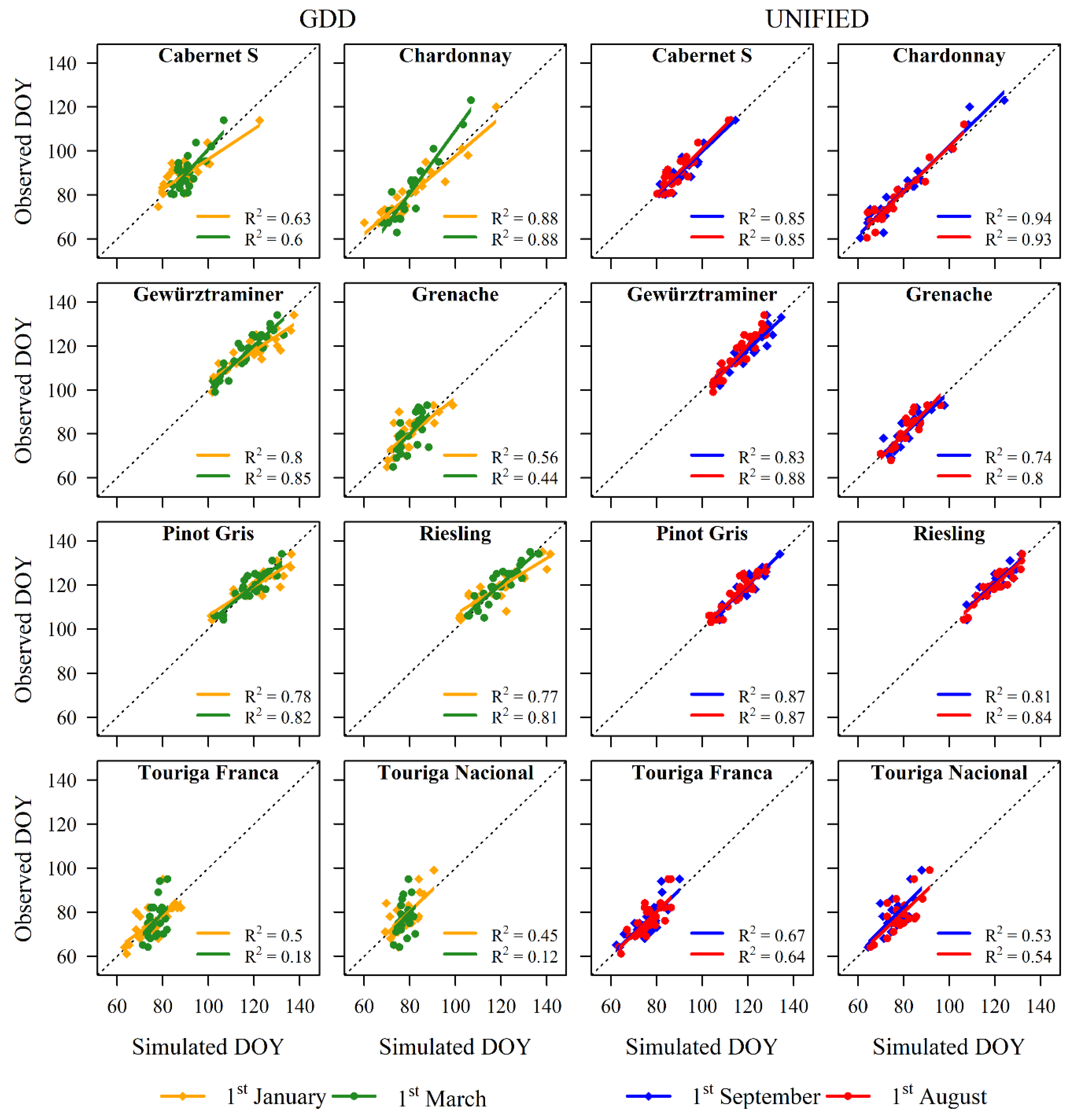
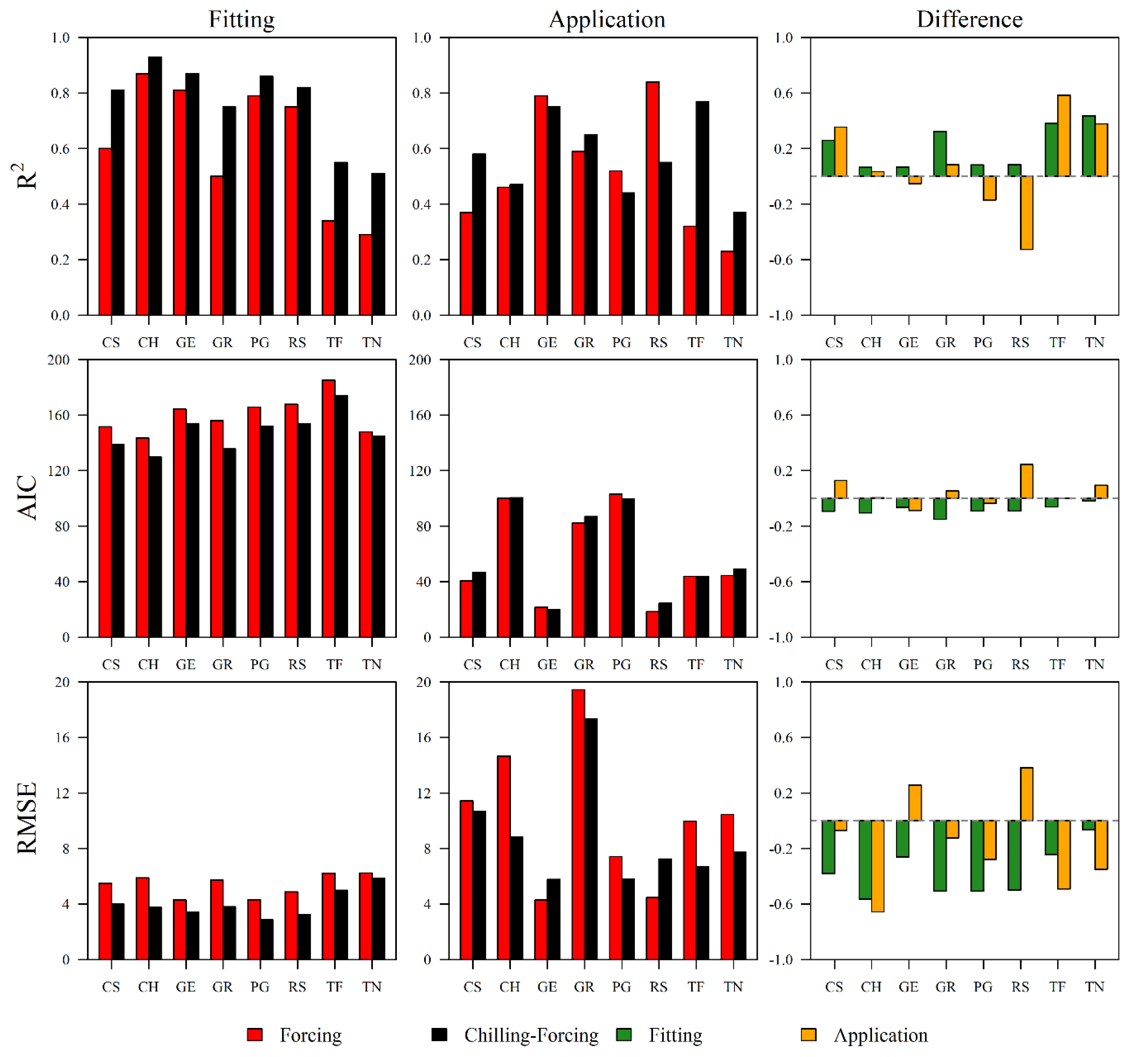
3.3. Model Applications on Other Independent Datasets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mullins, M.G.; Bouquet, A.; Williams, L.E. Biology of the Grapevine; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Van Leeuwen, C.; Seguin, G. The concept of terroir in viticulture. J. Wine Res. 2006, 17, 1–10. [Google Scholar] [CrossRef]
- Jones, G.V. Winegrape phenology. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Nederlands, 2013; pp. 563–584. [Google Scholar]
- Jones, G.V.; Davis, R.E. Climate influences on grapevine phenology, grape composition, and wine production and quality for Bordeaux, France. Am. J. Enol. Vitic. 2000, 51, 249–261. [Google Scholar]
- Menzel, A. A 500 year pheno-climatological view on the 2003 heatwave in Europe assessed by grape harvest dates. Meteorol. Zeitschrift 2005, 14, 75–77. [Google Scholar]
- Tomasi, D.; Jones, G.V.; Giust, M.; Lovat, L.; Gaiotti, F. Grapevine Phenology and Climate Change: Relationships and Trends in the Veneto Region of Italy for 1964-2009. Am. J. Enol. Vitic. 2011, 62, 329–339. [Google Scholar]
- Van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Pieri, P.; Parker, A.; de Rességuier, L.; Ollat, N. An Update on the Impact of Climate Change in Viticulture and Potential Adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Ramos, M.C.; Jones, G.V.; Martínez-Casasnovas, J.A. Structure and trends in climate parameters affecting winegrape production in northeast Spain. Clim. Res. 2008, 38, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Moriondo, M.; Bindi, M.; Fagarazzi, C.; Ferrise, R.; Trombi, G. Framework for high-resolution climate change impact assessment on grapevines at a regional scale. Reg. Environ. Chang. 2011, 11, 553–567. [Google Scholar]
- Gaál, M.; Moriondo, M.; Bindi, M. Modelling the impact of climate change on the Hungarian wine regions using Random Forest. Appl. Ecol. Environ. Res. 2012, 10, 121–140. [Google Scholar]
- Moriondo, M.; Jones, G.V.; Bois, B.; Dibari, C.; Ferrise, R.; Trombi, G.; Bindi, M. Projected shifts of wine regions in response to climate change. Clim. Change 2013, 119, 825–839. [Google Scholar] [CrossRef]
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Chang. 2018, 8, 29–37. [Google Scholar] [CrossRef]
- Fraga, H.; Pinto, J.G.; Santos, J.A. Climate change projections for chilling and heat forcing conditions in European vineyards and olive orchards: A multi-model assessment. Clim. Chang. 2019, 152, 179–193. [Google Scholar] [CrossRef]
- Malheiro, A.C.; Campos, R.; Fraga, H.; Eiras-Dias, J.; Silvestre, J.; Santos, J.A. Winegrape phenology and temperature relationships in the Lisbon wine region, Portugal. J. Int. Sci. Vigne Vin 2013, 47, 287–299. [Google Scholar]
- Costa, R.; Fraga, H.; Fonseca, A.; De Cortázar-Atauri, I.G.; Val, M.C.; Carlos, C.; Reis, S.; Santos, J.A. Grapevine phenology of cv. Touriga Franca and Touriga Nacional in the Douro wine region: Modelling and climate change projections. Agronomy 2019, 9, 1–20. [Google Scholar]
- Molitor, D.; Junk, J. Climate change is implicating a two-fold impact on air temperature increase in the ripening period under the conditions of the Luxembourgish grapegrowing region. OENO One 2019, 53, 409–422. [Google Scholar] [CrossRef]
- Shulman, Y.; Nir, G.; Fanberstein, L.; Lavee, S. The effect of cyanamide on the release from dormancy of grapevine buds. Sci. Hortic. (Amsterdam) 1983, 19, 97–104. [Google Scholar]
- Wicks, A.S.; Johnson, J.O.; Bracho, E.; Jensen, F.L.; Neja, R.A.; Lider, L.A.; Weaver, R.J. Induction of early and more uniform budbreak in Vitis vinifera L. cvs. Perlette, Thompson Seedless, and Flame Seedless. In Proceedings of the Bud Dormancy in Grapevine: Potential and Practical Uses of Hydrogen Cyanamide on Grapevine, Fresno, CA, USA, 20 August 1984; Weaver, R.J., Ed.; University of California: Fresno, CA, USA, 1984; pp. 48–58. [Google Scholar]
- Dokoozlian, N.K. Chilling temperature and duration interact on the budbreak of “Perlette” grapevine cuttings. HortScience 1999, 34, 1054–1056. [Google Scholar] [CrossRef] [Green Version]
- Sarvas, R. Investigations on the annual cycle of development of forest trees. Autumn dormancy and winter dormancy. Commun. Inst. For. Fenn. 1974, 84, 1–101. [Google Scholar]
- Pouget, R. Considérations Générales sur le rythme végétatif et la dormance des bourgeons de la Vigne. Vitis 1972, 217, 198–217. [Google Scholar]
- Pouget, R. Nouvelle conception du seuil de croissance chez la vigne. Vitis 1968, 7, 201–205. [Google Scholar]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R.; Walker, D.I. A model for estimating the completion of rest for “Redhaven” and “Elberta” peach trees. Hortic. Sci. 1974, 9, 331–332. [Google Scholar]
- Chuine, I. A unified model for budburst of trees. J. Theor. Biol. 2000, 207, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Cesaraccio, C.; Spano, D.; Snyder, R.L.; Duce, P. Chilling and forcing model to predict bud-burst of crop and forest species. Agric. For. Meteorol. 2004, 126, 1–13. [Google Scholar] [CrossRef]
- Garcia de Cortazar-Atauri, I.; Brisson, N.; Seguin, B.; Gaudillere, J.; Baculat, E. Simulation of budbreak date for vine. The bring model. Some application in climate change study. In Proceedings of the XIV International GESCO Viticulture Congress, Geisenheim, Germany, 23–27 August 2005; pp. 485–490. [Google Scholar]
- Nendel, C. Grapevine bud break prediction for cool winter climates. Int. J. Biometeorol. 2010, 54, 231–241. [Google Scholar] [CrossRef]
- Caffarra, A.; Eccel, E. Projecting the impacts of climate change on the phenology of grapevine in a mountain area. Aust. J. Grape Wine Res. 2011, 17, 52–61. [Google Scholar] [CrossRef]
- García de Cortázar-Atauri, I.; Brisson, N.; Gaudillere, J.P. Performance of several models for predicting budburst date of grapevine (Vitis vinifera L.). Int. J. Biometeorol. 2009, 53, 317–326. [Google Scholar] [CrossRef]
- Fila, G.; Gardiman, M.; Belvini, P.; Meggio, F.; Pitacco, A. A comparison of different modelling solutions for studying grapevine phenology under present and future climate scenarios. Agric. For. Meteorol. 2014, 195–196, 192–205. [Google Scholar] [CrossRef]
- Leolini, L.; Moriondo, M.; Fila, G.; Costafreda-Aumedes, S.; Ferrise, R.; Bindi, M. Late spring frost impacts on future grapevine distribution in Europe. Field Crops Res. 2018, 222, 197–208. [Google Scholar] [CrossRef]
- Moriondo, M.; Ferrise, R.; Trombi, G.; Brilli, L.; Dibari, C.; Bindi, M. Environmental Modelling & Software Modelling olive trees and grapevines in a changing climate *. Environ. Model. Softw. 2015, 72, 387–401. [Google Scholar]
- Molitor, D.; Junk, J.; Evers, D.; Hoffmann, L.; Beyer, M. A high-resolution cumulative degree day-based model to simulate phenological development of grapevine. Am. J. Enol. Vitic. 2014, 65, 72–80. [Google Scholar] [CrossRef]
- Menne, M.J.; Durre, I.; Vose, R.S.; Gleason, B.E.; Houston, T.G. An overview of the global historical climatology network-daily database. J. Atmos. Ocean. Technol. 2012, 29, 897–910. [Google Scholar] [CrossRef]
- Destrac-Irvine, A.; van Leeuwen, C. VitAdapt: An experimental program to study the behavior of a wide range of Vitis vinifera varieties in a context of climate change in the Bordeaux vineyards. In Proceedings of the Sustainable Grape and Wine Production in the Context of Climate Change. ClimWine 2016 International Symposium, Bordeaux, France, 10–13 April 2016. [Google Scholar]
- Garcia de Cortazar-Atauri, I.; Audergon, J.M.; Delzon, S.; Anger, C.; Duchêne, E.; Bonhomme, M.; Legave, J.M.; Chuine, I.; Raynal, H.; Davi, H.; et al. PERPHECLIM ACCAF Project-perennial fruit crops and forest phenology evolution facing climatic changes. In Proceedings of the XVI International Sumposium on Apricot Breeding and Culture and XV Chinese National Symposium on Plum and Apricot, Shenyang, China, 29 June–3 July 2015. [Google Scholar]
- Jones, G.V. Climate and Terroir: Impacts of Climate Variability and Change on Wine. In Fine Wine and Terroir-the Geoscience Perspective; Meinert, L.D., Ed.; Geological Association of Canada: St. John’s, NL, Canada, 2006; p. 247. [Google Scholar]
- Eichhorn, V.K.W.; Lorenz, D.H. Phänologische Entwicklungsstadien der Rebe [Phenological development stages of the grapevine]. Nachr. Dtsch. Pflanzenschutzd. 1977, 29, 119–120. [Google Scholar]
- Amerine, M.A.; Winkler, A.J. Composition and Quality of Musts and Wines of California Grapes. Hilgardia 1944, 15, 493–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.; Engel, T. Simulation of phenological development of wheat crops. Agric. Syst. 1998, 58, 1–24. [Google Scholar] [CrossRef]
- Bidabe, B. Contrôle de l’époque de floraison du pommier par une nouvelle conception de l’action de températures. C. R. Séances L’Acad. D’Agric. Fr. 1963, 49, 934–945. [Google Scholar]
- Bidabe, B. L’action Des Températures Sur L’évolution Des Bourgeons de L’entrée en Dormance à la Floraison; Congrès Pomologique: Paris, France, 1965. [Google Scholar]
- Tukey, J.W. Box-and-whisker plots. In Exploratory Data Analysis; Addison-Wesley: Reading, MA, USA, 1977; pp. 39–43. ISBN 978-0201076165. [Google Scholar]
- Shellie, K.C. Viticultural performance of red and white wine grape cultivars in southwestern Idaho. Horttechnology 2007, 17, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.K.; Garcia de Cortazar-Atauri, I.; Van Leeuwen, C.; Chuine, I. General phenological model to characterise the timing of flowering and veraison of Vitis vinifera L. Aust. J. Grape Wine Res. 2011, 17, 206–216. [Google Scholar] [CrossRef]
- Fila, G. Mathematical Models for the Analysis of the Spatio-Temporal Variability of Vine Phenology [Modelli Matematici per L’analisi Della Variabilità Spazio-Temporale Della Fenologia Della Vite]. Ph.D. Thesis, University of Padua, Padua, Italy, 2012. [Google Scholar]
- Streck, N.A.; Weiss, A.; Xue, Q.; Stephen Baenziger, P. Improving predictions of developmental stages in winter wheat: A modified Wang and Engel model. Agric. For. Meteorol. 2003, 115, 139–150. [Google Scholar] [CrossRef]
- Chuine, I.; Garcia de Cortazar-Atauri, I.; Kramer, K.; Hänninen, H. Plant development models. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Nederlands, 2013; pp. 275–293. ISBN 9789400769250. [Google Scholar]
- Hastings, W.K. Monte carlo sampling methods using Markov chains and their applications. Biometrika 1970, 57, 97–109. [Google Scholar] [CrossRef]
- Kirkpatrick, S.; Gelatt, C.D.; Vecchi, M.P. Optimization by simulated annealing. Science 1983, 220, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Metropolis, N.; Rosenbluth, A.W.; Rosenbluth, M.N.; Teller, A.H.; Teller, E. Equation of state calculations by fast computing machines. J. Chem. Phys. 1953, 21, 1087–1093. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Version 3.5.1; R Foundation for Statisrical Computing: Vienna, Austria, 2018; ISBN 3900051070. [Google Scholar]
- Wright, S. Correlation and causation. J. Agric. Res. 1921, 20, 557–585. [Google Scholar]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Proceedings of the Second International Symposium on Information Theory, Tsahkadsor, Armenia, 2–8 September 1973; Petrov, B.N., Caski, F., Eds.; Akademiai Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Kenney, J.F. Root mean square. In Mathematics of Statistics; Chapman & Hall, LTD.: Princeton, NJ, USA, 1962; pp. 59–60. [Google Scholar]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models part I—A discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar]
- Cola, G.; Failla, O.; Maghradze, D.; Megrelidze, L.; Mariani, L. Grapevine phenology and climate change in Georgia. Int. J. Biometeorol. 2017, 61, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Fila, G.; Di, B.; Gardiman, M.; Storchi, P.; Tomasi, D.; Silvestroni, O.; Pitacco, A. Calibration and validation of grapevine budburst models using growth-room experiments as data source. Agric. For. Meteorol. 2012, 160, 69–79. [Google Scholar] [CrossRef]
- Duchêne, E.; Huard, F.; Dumas, V.; Schneider, C.; Merdinoglu, D. The challenge of adapting grapevine varieties to climate change. Clim. Res. 2010, 41, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Reis Pereira, M.; Ribeiro, H.; Abreu, I.; Eiras-Dias, J.; Mota, T.; Cunha, M. Predicting the flowering date of Portuguese grapevine varieties using temperature-based phenological models: A multi-site approach. J. Agric. Sci. 2018, 156, 865–876. [Google Scholar] [CrossRef]
- Moriondo, M.; Orlandini, S.; Nuntiis, P.D.; Mandrioli, P. Effect of agrometeorological parameters on the phenology of pollen emission and production of olive trees (Olea europea L. Aerobiologia 2001, 17, 225–232. [Google Scholar]
- Chuine, I.; Bonhomme, M.; Legave, J.M.; García de Cortázar-Atauri, I.; Charrier, G.; Lacointe, A.; Améglio, T. Can phenological models predict tree phenology accurately in the future? The unrevealed hurdle of endodormancy break. Glob. Chang. Biol. 2016, 22, 3444–3460. [Google Scholar]
- Martínez-Lüscher, J.; Kizildeniz, T.; Vučetić, V.; Dai, Z.; Luedeling, E.; van Leeuwen, C.; Gomès, E.; Pascual, I.; Irigoyen, J.J.; Morales, F.; et al. Sensitivity of Grapevine Phenology to Water Availability, Temperature and CO2 Concentration. Front. Environ. Sci. 2016, 4, 48. [Google Scholar] [CrossRef]
- Fraga, H.; García de Cortázar Atauri, I.; Malheiro, A.C.; Santos, J.A. Modelling climate change impacts on viticultural yield, phenology and stress conditions in Europe. Glob. Chang. Biol. 2016, 22, 3774–3788. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [PubMed] [Green Version]
- Fraga, H.; Santos, J.A.; Malheiro, A.C.; Oliveira, A.A.; Moutinho-Pereira, J.; Jones, G.V. Climatic suitability of Portuguese grapevine varieties and climate change adaptation. Int. J. Climatol. 2016, 36, 1–12. [Google Scholar]
- Camargo-Alvarez, H.; Salazar-Gutiérrez, M.; Keller, M.; Hoogenboom, G. Modeling the effect of temperature on bud dormancy of grapevines. Agric. For. Meteorol. 2020, 280, 107782. [Google Scholar] [CrossRef]
- Fennell, A.; Mathiason, K.; Luby, J. Genetic segregation for indicators of photoperiod control of dormancy induction in vitis species. Acta Hortic. 2005, 689, 533–540. [Google Scholar] [CrossRef]
- Rubio, S.; Dantas, D.; Bressan-Smith, R.; Pérez, F.J. Relationship Between Endodormancy and Cold Hardiness in Grapevine Buds. J. Plant Growth Regul. 2016, 35, 266–275. [Google Scholar] [CrossRef]
- Leolini, L.; Moriondo, M.; Romboli, Y.; Gardiman, M.; Costafreda-Aumedes, S.; Costafreda-Aumedes, S.; Bindi, M.; Granchi, L.; Brilli, L. Modelling sugar and acid content in Sangiovese grapes under future climates: An Italian case study. Clim. Res. 2019, 78, 211–224. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety × Site | Weather Data | Budbreak Data | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Procedure | Variety | Country | Weather Station | Period | Vineyard | Period | Cases | ||
| Lat | Long | Years | Lat | Long | Years | Number | |||
| Fitting & Validation | Cabernet Sauvignon | France | 43.58 | 3.96 | 1950–2017 | 43.33 | 3.56 | 1951–2012 | 39 |
| Luxembourg | 49.50 | 6.35 | 2016–2018 | 49.50 | 6.35 | 2017–2018 | 2 | ||
| Chardonnay | France | 43.58 | 3.96 | 1950–2017 | 43.33 | 3.56 | 1951–2012 | 33 | |
| Luxembourg | 49.50 | 6.35 | 2010–2018 | 49.50 | 6.35 | 2011–2018 | 8 | ||
| Gewürztraminer | Luxembourg | 49.50 | 6.35 | 1970–2018 | 49.50 | 6.35 | 1972–2018 | 50 | |
| Grenache | France | 43.58 | 3.96 | 1950–2017 | 43.33 | 3.56 | 1951–2012 | 41 | |
| Pinot Gris | Luxembourg | 49.50 | 6.35 | 1970–2018 | 49.50 | 6.35 | 1971–2018 | 51 | |
| Riesling | Luxembourg | 49.50 | 6.35 | 1970–2018 | 49.50 | 6.35 | 1971–2018 | 50 | |
| Touriga Franca | Portugal | 39.04 | −9.18 | 1994–2014 | 39.04 | −9.18 | 1995–2014 | 20 | |
| 41.81 | −8.41 | 2004–2009 | 41.81 | −8.41 | 2005–2009 | 5 | |||
| 41.25 | −7.11 | 2013–2018 | 41.25 | −7.11 | 2014–2018 | 5 | |||
| 41.19 | −7.54 | 2013–2018 | 41.19 | −7.54 | 2014–2018 | 5 | |||
| 41.04 | −7.04 | 2013–2017 | 41.04 | −7.04 | 2014–2017 | 4 | |||
| 41.17 | −7.56 | 2013–2017 | 41.17 | −7.56 | 2014–2017 | 4 | |||
| 41.15 | −7.62 | 2013–2018 | 41.15 | −7.62 | 2014–2018 | 5 | |||
| 41.17 | −7.55 | 2014–2018 | 41.17 | −7.55 | 2015–2018 | 4 | |||
| Touriga Nacional | Luxembourg | 49.50 | 6.35 | 2016–2018 | 49.50 | 6.35 | 2017–2018 | 2 | |
| Portugal | 39.04 | −9.18 | 1989–2014 | 39.04 | −9.18 | 1990–2014 | 19 | ||
| 41.81 | −8.41 | 2004–2009 | 41.81 | −8.41 | 2005–2009 | 5 | |||
| 41.21 | −7.43 | 2013–2018 | 41.21 | −7.43 | 2014–2018 | 5 | |||
| 41.24 | −7.76 | 2014–2017 | 41.24 | −7.76 | 2015–2017 | 3 | |||
| 41.22 | −7.54 | 2013–2018 | 41.22 | −7.54 | 2014–2018 | 5 | |||
| 41.15 | −7.76 | 2015–2018 | 41.15 | −7.76 | 2016–2018 | 3 | |||
| Application | Cabernet Sauvignon | France | 44.79 | −0.58 | 2012–2019 | 44.79 | −0.58 | 2013–2019 | 7 |
| Chardonnay | France | 48.55 | 7.64 | 1950–2017 | 48.22 | 7.35 | 1976–1990 | 15 | |
| Gewürztraminer | France | 48.55 | 7.64 | 1950–2017 | 48.22 | 7.35 | 2006–2009 | 4 | |
| Grenache | France | 48.55 | 7.64 | 1950–2017 | 48.22 | 7.35 | 1976–1990 | 13 | |
| Pinot Gris | France | 48.55 | 7.64 | 1950–2017 | 48.22 | 7.35 | 1976–1990 | 15 | |
| Riesling | France | 48.55 | 7.64 | 1950–2017 | 48.22 | 7.35 | 2006–2009 | 4 | |
| Touriga Franca | France | 44.79 | −0.58 | 2012–2019 | 44.79 | −0.58 | 2013–2019 | 7 | |
| Touriga Nacional | France | 44.79 | −0.58 | 2012–2019 | 44.79 | −0.58 | 2013–2019 | 7 | |
| GDD | WANG | UNIFORC/UNICHILL/UNIFIED | BRIN h/BRIN d | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety | t0 | Fcrit | Tb | Fcrit | Topt | Tmin | Tmax | Ccrit | Fcrit | a | b | c | d | e | w | z | Ccrit | Fcrit | Q10c | Tl | Th |
| Cabernet Sauvignon | 1 −121 | 763.47 | 0 | 67.32 | 18.15 | 0 | 45 | - | 16.39 | - | - | - | −0.21 | 16.47 | - | - | 190.14 184.86 | 6636.2 295.3 | 1.54 1.54 | 5.07 4.45 | 25 |
| 142.36 | 26.54 | 0.15 | −23.61 | −20.41 | −0.19 | 11.80 | - | - | |||||||||||||
| 151.14 | 20.62 | 1.31 | −31.06 | −1.45 | −0.20 | 12.44 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 286.89 | 1.18 | 19.49 | 23.96 | 0 | 45 | - | 7.74 | - | - | - | −0.21 | 16.45 | - | - | 217.53 206.55 | 6597.1 272.1 | 1.52 1.59 | 5.07 4.84 | 25 | |
| 178.96 | 23.19 | 0.16 | −15.33 | −24.24 | −0.25 | 11.83 | - | - | |||||||||||||
| 175.24 | 20.40 | 0.91 | −26.60 | −1.63 | −0.14 | 14.69 | −0.00025 | 21.05 | |||||||||||||
| Chardonnay | 1 −121 | 432.23 | 2.34 | 14.63 | 30.42 | 0 | 45 | - | 8.58 | - | - | - | −0.21 | 18.46 | - | - | 148.48 148.34 | 5194.0 238.3 | 1.54 1.54 | 5.86 4.99 | 25 |
| 132.02 | 17.55 | 0.72 | −29.03 | −17.19 | −0.24 | 12.56 | - | - | |||||||||||||
| 123.96 | 19.71 | 0.85 | −32.05 | −2.20 | −0.28 | 11.79 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 49.18 | 7.21 | 4.00 | 31.26 | 0 | 45 | - | 1.33 | - | - | - | −0.30 | 18.36 | - | - | 173.38 172.90 | 5276.9 211.9 | 1.54 1.54 | 5.81 5.44 | 25 | |
| 165.85 | 19.05 | 0.46 | −19.82 | −13.84 | −0.24 | 11.93 | - | - | |||||||||||||
| 172.71 | 19.63 | 1.19 | −34.66 | 0.22 | −0.25 | 10.46 | −0.00025 | 21.05 | |||||||||||||
| Grenache | 1 −121 | 645.68 | 0 | 47.27 | 21.91 | 0 | 45 | - | 13.40 | - | - | - | −0.21 | 16.48 | - | - | 177.23 158.40 | 3769.3 312.9 | 1.67 1.55 | 6.63 4.02 | 25 |
| 156.67 | 8.05 | 0.61 | −20.19 | −11.39 | −0.19 | 16.94 | - | - | |||||||||||||
| 157.54 | 20.94 | 0.55 | −30.18 | −10.31 | −0.26 | 9.98 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 85.11 | 5.99 | 5.17 | 30.40 | 0 | 45 | - | 4.17 | - | - | - | −0.21 | 16.49 | - | - | 215.25 210.27 | 5029.4 319.2 | 1.53 1.54 | 5.27 2.80 | 25 | |
| 186.21 | 4.73 | 0.79 | −26.99 | −7.35 | −0.18 | 21.33 | - | - | |||||||||||||
| 186.89 | 20.41 | 1.04 | −30.11 | −1.81 | −0.28 | 9.72 | −0.00025 | 21.05 | |||||||||||||
| Gewürrtztraminer | 1 −121 | 137.17 | 7.10 | 12.11 | 31.26 | 0 | 45 | - | 9.37 | - | - | - | −0.21 | 18.50 | - | - | 283.90 280.72 | 3961.1 181.3 | 1.53 1.54 | 6.19 5.39 | 25 |
| 186.01 | 17.05 | 0.17 | −32.51 | −12.74 | −0.35 | 10.79 | - | - | |||||||||||||
| 172.62 | 20.42 | 1.18 | −34.32 | −10.15 | −0.23 | 11.48 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 255.89 | 3.61 | 9.17 | 31.26 | 0 | 45 | - | 8.43 | - | - | - | −0.23 | 16.45 | - | - | 312.24 311.34 | 4159.1 182.5 | 1.53 1.54 | 5.98 5.36 | 25 | |
| 161.74 | 12.51 | 1.62 | −34.01 | −6.44 | −0.33 | 12.71 | - | - | |||||||||||||
| 214.68 | 20.29 | 0.50 | −31.89 | −10.95 | −0.28 | 10.28 | −0.00025 | 21.05 | |||||||||||||
| Pinot Gris | 1 −121 | 122.16 | 7.39 | 12.15 | 31.26 | 0 | 45 | - | 9.63 | - | - | - | −0.21 | 18.49 | - | - | 285.01 284.32 | 4633.1 225.9 | 1.53 1.55 | 5.22 4.11 | 25 |
| 183.87 | 14.70 | 0.56 | −23.01 | −19.53 | −0.25 | 12.67 | - | - | |||||||||||||
| 183.25 | 20.45 | 0.95 | −30.94 | −10.68 | −0.28 | 10.26 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 250.48 | 3.91 | 9.36 | 31.23 | 0 | 45 | - | 9.44 | - | - | - | −0.21 | 16.54 | - | - | 314.40 314.75 | 6374.7 218.9 | 1.53 1.54 | 3.52 4.29 | 25 | |
| 174.41 | 12.60 | 1.13 | −25.39 | −8.01 | −0.33 | 12.02 | - | - | |||||||||||||
| 211.57 | 20.26 | 0.36 | −11.66 | −7.14 | −0.28 | 10.31 | −0.00025 | 21.05 | |||||||||||||
| Riesling | 1 −121 | 156.14 | 6.74 | 12.60 | 31.26 | 0 | 45 | - | 9.90 | - | - | - | −0.21 | 18.46 | - | - | 267.03 280.95 | 4980.8 208.2 | 1.53 1.56 | 5.44 4.93 | 25 |
| 150.99 | 5.33 | 1.39 | −27.15 | −5.31 | −0.18 | 21.68 | - | - | |||||||||||||
| 173.34 | 20.47 | 1.09 | −28.42 | −10.79 | −0.21 | 11.23 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 279.34 | 3.46 | 11.80 | 30.29 | 0 | 45 | - | 10.14 | - | - | - | −0.22 | 16.45 | - | - | 297.05 313.12 | 5022.2 221.8 | 1.53 1.53 | 5.41 4.68 | 25 | |
| 168.39 | 10.63 | 1.08 | −23.72 | −7.63 | −0.29 | 13.89 | - | - | |||||||||||||
| 163.61 | 20.58 | 1.00 | −19.62 | −5.65 | −0.26 | 10.30 | −0.00025 | 21.05 | |||||||||||||
| Touriga Franca | 1 −121 | 743.93 | 0 | 36.53 | 26.70 | 0 | 45 | - | 16.61 | - | - | - | −0.21 | 16.45 | - | - | 158.30 144.69 | 7933.9 493.9 | 1.53 1.52 | 4.80 2.89 | 25 |
| 142.03 | 24.35 | 0.83 | −21.47 | −1.62 | −0.32 | 10.79 | - | - | |||||||||||||
| 141.22 | 20.61 | 0.42 | −14.45 | −10.02 | −0.27 | 12.24 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 149.08 | 2.86 | 7.61 | 28.57 | 0 | 45 | - | 4.84 | - | - | - | −0.21 | 16.45 | - | - | 180.41 183.23 | 6998.5 293.1 | 1.52 1.52 | 5.79 5.28 | 25 | |
| 171.91 | 24.24 | 0.33 | −18.24 | −25.30 | −0.23 | 11.70 | - | - | |||||||||||||
| 159.86 | 20.53 | 0.38 | −13.35 | −5.10 | −0.19 | 14.67 | −0.00025 | 21.05 | |||||||||||||
| Touriga Nacional | 1 −121 | 688.00 | 1.06 | 35.57 | 26.38 | 0 | 45 | - | 16.11 | - | - | - | −0.21 | 16.46 | - | - | 133.81 123.57 | 13,660.2 774.2 | 1.59 1.54 | 2.15 0.92 | 25 |
| 117.39 | 21.11 | 0.19 | −22.31 | −18.25 | −0.21 | 15.44 | - | - | |||||||||||||
| 124.23 | 19.76 | 0.47 | −27.59 | −0.002 | −0.14 | 17.19 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 199.94 | 0.80 | 8.19 | 28.71 | 0 | 45 | - | 5.20 | - | - | - | −0.21 | 16.45 | - | - | 165.37 144.19 | 12,985.3 823.9 | 1.57 1.55 | 2.40 0.42 | 25 | |
| 148.45 | 20.97 | 0.61 | −22.96 | −6.80 | −0.21 | 15.47 | - | - | |||||||||||||
| 150.90 | 20.54 | 0.26 | −8.36 | −0.72 | −0.14 | 17.87 | −0.00025 | 21.05 | |||||||||||||
| GDD | WANG | UNIFORC/UNICHILL/UNIFIED | BRIN h/BRIN d | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variety | t0 | Fcrit | Tb | Fcrit | Topt | Tmin | Tmax | Ccrit | Fcrit | a | b | c | d | e | w | z | Ccrit | Fcrit | Q10c | Tl | Th |
| Cabernet Sauvignon | 1 −121 | 765.16 | 0 | 68.54 | 17.99 | 0 | 45 | - | 16.42 | - | - | - | −0.21 | 16.45 | - | - | 182.31 177.95 | 11,075.3 508.7 | 1.54 1.55 | 2.07 1.49 | 25 |
| 145.70 | 24.79 | 0.18 | −20.62 | −20.10 | −0.19 | 11.91 | - | - | |||||||||||||
| 152.62 | 20.60 | 0.94 | −23.60 | −2.86 | −0.22 | 12.08 | -0.00025 | 21.05 | |||||||||||||
| 60 −152 | 225.25 | 3.16 | 11.83 | 28.64 | 0 | 45 | - | 7.31 | - | - | - | −0.21 | 16.46 | - | - | 196.01 193.98 | 11,942.8 496.5 | 1.56 1.61 | 1.91 1.80 | 25 | |
| 176.29 | 25.03 | 0.18 | −21.03 | −18.95 | −0.19 | 11.89 | - | - | |||||||||||||
| 180.78 | 20.59 | 0.87 | −27.91 | −4.13 | −0.19 | 12.69 | -0.00025 | 21.05 | |||||||||||||
| Chardonnay | 1 −121 | 527.48 | 0.91 | 22.21 | 28.11 | 0 | 45 | - | 11.02 | - | - | - | −0.22 | 16.91 | - | - | 150.00 167.46 | 6316.0 254.1 | 1.61 1.54 | 4.97 4.63 | 25 |
| 127.15 | 14.36 | 0.57 | −22.95 | −14.48 | −0.25 | 14.42 | - | - | |||||||||||||
| 130.52 | 20.32 | 0.88 | −24.70 | −2.52 | −0.27 | 11.40 | -0.00025 | 21.05 | |||||||||||||
| 60 −152 | 47.34 | 7.24 | 4.10 | 21.26 | 0 | 45 | - | 1.50 | - | - | - | −0.30 | 18.30 | - | - | 171.33 184.56 | 5931.7 266.4 | 1.61 1.55 | 5.35 4.44 | 25 | |
| 160.41 | 17.18 | 0.65 | −27.13 | −13.07 | −0.25 | 13.11 | - | - | |||||||||||||
| 163.81 | 20.45 | 0.85 | −28.65 | −3.25 | −0.26 | 11.35 | -0.00025 | 21.05 | |||||||||||||
| Grenache | 1 −121 | 653.93 | 0 | 54.66 | 19.74 | 0 | 45 | - | 13.61 | - | - | - | −0.21 | 16.50 | - | - | 184.09 172.80 | 6316.6 286.7 | 1.58 1.66 | 4.31 3.87 | 25 |
| 126.06 | 8.07 | 1.04 | −26.44 | −8.24 | −0.26 | 16.68 | - | - | |||||||||||||
| 151.30 | 20.59 | 0.78 | −24.27 | −5.82 | −0.24 | 10.49 | -0.00025 | 21.05 | |||||||||||||
| 60 −152 | 113.43 | 4.85 | 6.61 | 29.66 | 0 | 45 | - | 3.96 | - | - | - | −0.23 | 17.10 | - | - | 182.40 198.81 | 8882.5 324.5 | 1.67 1.60 | 2.97 3.11 | 25 | |
| 150.54 | 10.30 | 0.90 | −28.01 | −10.48 | −0.24 | 16.05 | - | - | |||||||||||||
| 173.87 | 20.58 | 0.80 | −25.81 | −5.45 | −0.24 | 11.04 | -0.00025 | 21.05 | |||||||||||||
| Gewürrtztraminer | 1 −121 | 178.85 | 6.24 | 11.97 | 31.26 | 0 | 45 | - | 8.95 | - | - | - | −0.23 | 18.17 | - | - | 251.39 264.22 | 5324.3 228.9 | 1.71 1.60 | 5.01 4.53 | 25 |
| 154.79 | 13.62 | 0.88 | −28.20 | −14.15 | −0.26 | 14.32 | - | - | |||||||||||||
| 149.42 | 20.26 | 0.89 | −23.75 | −8.10 | −0.25 | 11.17 | -0.00025 | 21.05 | |||||||||||||
| 60 −152 | 314.55 | 2.55 | 12.43 | 29.91 | 0 | 45 | - | 9.12 | - | - | - | −0.22 | 16.46 | - | - | 280.20 291.30 | 5274.6 232.9 | 1.66 1.62 | 5.05 4.44 | 25 | |
| 157.90 | 12.92 | 0.93 | −26.85 | −13.41 | −0.25 | 14.65 | - | - | |||||||||||||
| 176.90 | 20.45 | 0.71 | −25.17 | −8.95 | −0.26 | 10.95 | −0.00025 | 21.05 | |||||||||||||
| Pinot Gris | 1 −121 | 126.62 | 7.39 | 12.26 | 31.26 | 0 | 45 | - | 7.61 | - | - | - | −0.25 | 18.35 | - | - | 271.42 269.51 | 6580.6 287.7 | 1.56 1.59 | 3.75 3.29 | 25 |
| 169.54 | 14.04 | 0.89 | −27.31 | −13.36 | −0.23 | 14.11 | - | - | |||||||||||||
| 173.20 | 20.38 | 0.69 | −22.47 | −9.57 | −0.25 | 10.82 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 359.67 | 1.77 | 15.09 | 28.93 | 0 | 45 | - | 9.48 | - | - | - | −0.21 | 16.47 | - | - | 292.53 299.62 | 6507.2 290.8 | 1.61 1.57 | 3.86 3.24 | 25 | |
| 188.72 | 14.01 | 0.85 | −25.77 | −12.46 | −0.24 | 14.19 | - | - | |||||||||||||
| 184.87 | 20.47 | 0.77 | −21.17 | −8.63 | −0.27 | 10.65 | −0.00025 | 21.05 | |||||||||||||
| Riesling | 1 −121 | 156.13 | 6.78 | 12.43 | 31.26 | 0 | 45 | - | 8.51 | - | - | - | −0.23 | 18.47 | - | - | 267.92 270.05 | 6755.6 317.7 | 1.65 1.63 | 3.75 2.84 | 25 |
| 167.21 | 13.93 | 0.94 | −25.59 | −11.82 | −0.24 | 14.39 | - | - | |||||||||||||
| 166.51 | 20.56 | 0.95 | −24.12 | −8.63 | −0.26 | 10.92 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 416.44 | 0.93 | 16.95 | 28.35 | 0 | 45 | - | 9.76 | - | - | - | −0.21 | 16.65 | - | - | 294.27 296.64 | 6967.0 297.4 | 1.66 1.66 | 3.58 3.20 | 25 | |
| 176.49 | 13.62 | 0.99 | −27.07 | −12.29 | −0.27 | 13.61 | - | - | |||||||||||||
| 174.36 | 20.30 | 1.03 | −25.01 | −7.57 | −0.26 | 10.71 | −0.00025 | 21.05 | |||||||||||||
| Touriga Franca | 1 −121 | 759.41 | 0.07 | 36.55 | 26.40 | 0 | 45 | - | 15.97 | - | - | - | −0.21 | 16.87 | - | - | 154.21 149.90 | 8897.4 363.8 | 1.54 1.57 | 4.39 4.42 | 25 |
| 137.46 | 19.81 | 0.69 | −22.91 | −8.82 | −0.24 | 14.33 | - | - | |||||||||||||
| 137.41 | 20.48 | 0.73 | −23.67 | −6.05 | −0.23 | 13.26 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 143.12 | 3.71 | 7.49 | 28.97 | 0 | 45 | - | 4.96 | - | - | - | −0.21 | 16.46 | - | - | 178.92 178.72 | 8466.3 387.1 | 1.54 1.56 | 4.63 4.20 | 25 | |
| 148.81 | 24.27 | 0.63 | −25.76 | −11.79 | −0.22 | 14.28 | - | - | |||||||||||||
| 165.50 | 20.42 | 0.64 | −22.47 | −5.83 | −0.21 | 13.90 | −0.00025 | 21.05 | |||||||||||||
| Touriga Nacional | 1 −121 | 710.02 | 0.77 | 37.39 | 26.29 | 0 | 45 | - | 16.73 | - | - | - | −0.21 | 16.49 | - | - | 106.82 118.99 | 15,757.2 694.1 | 1.81 1.58 | 2.14 1.98 | 25 |
| 108.60 | 19.98 | 0.59 | −23.76 | −7.24 | −0.22 | 15.56 | - | - | |||||||||||||
| 108.85 | 20.33 | 0.72 | −24.30 | −2.98 | −0.16 | 16.91 | −0.00025 | 21.05 | |||||||||||||
| 60 −152 | 176.18 | 1.89 | 9.42 | 26.87 | 0 | 45 | - | 4.86 | - | - | - | −0.21 | 16.66 | - | - | 134.59 140.60 | 16,289.3 691.0 | 1.71 1.65 | 1.86 1.92 | 25 | |
| 123.73 | 20.27 | 0.62 | −23.51 | −12.11 | −0.22 | 15.53 | - | - | |||||||||||||
| 139.52 | 20.67 | 0.82 | −25.39 | −3.31 | −0.16 | 16.66 | −0.00025 | 21.05 | |||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leolini, L.; Costafreda-Aumedes, S.; A. Santos, J.; Menz, C.; Fraga, H.; Molitor, D.; Merante, P.; Junk, J.; Kartschall, T.; Destrac-Irvine, A.; et al. Phenological Model Intercomparison for Estimating Grapevine Budbreak Date (Vitis vinifera L.) in Europe. Appl. Sci. 2020, 10, 3800. https://doi.org/10.3390/app10113800
Leolini L, Costafreda-Aumedes S, A. Santos J, Menz C, Fraga H, Molitor D, Merante P, Junk J, Kartschall T, Destrac-Irvine A, et al. Phenological Model Intercomparison for Estimating Grapevine Budbreak Date (Vitis vinifera L.) in Europe. Applied Sciences. 2020; 10(11):3800. https://doi.org/10.3390/app10113800
Chicago/Turabian StyleLeolini, Luisa, Sergi Costafreda-Aumedes, João A. Santos, Christoph Menz, Helder Fraga, Daniel Molitor, Paolo Merante, Jürgen Junk, Thomas Kartschall, Agnès Destrac-Irvine, and et al. 2020. "Phenological Model Intercomparison for Estimating Grapevine Budbreak Date (Vitis vinifera L.) in Europe" Applied Sciences 10, no. 11: 3800. https://doi.org/10.3390/app10113800
APA StyleLeolini, L., Costafreda-Aumedes, S., A. Santos, J., Menz, C., Fraga, H., Molitor, D., Merante, P., Junk, J., Kartschall, T., Destrac-Irvine, A., van Leeuwen, C., C. Malheiro, A., Eiras-Dias, J., Silvestre, J., Dibari, C., Bindi, M., & Moriondo, M. (2020). Phenological Model Intercomparison for Estimating Grapevine Budbreak Date (Vitis vinifera L.) in Europe. Applied Sciences, 10(11), 3800. https://doi.org/10.3390/app10113800