A Model for Assessing the Quantitative Effects of Heterogeneous Affinity in Malaria Transmission along with Ivermectin Mass Administration



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods, Model, and Analysis Tools
2.1. Implementing Heterogeneity in Malaria Spreading
2.2. Human-to-Mosquito Transmission and Drug Administration in a Heterogeneity Setting
2.3. Predictive Rates for Assessing the Strength of Malaria Transmission and the Annual Entomological Inoculation
3. Results
3.1. From Disease Persistence to Elimination in a Scenario of Increasing Heterogeneity
3.1.1. Heterogeneity and Ivermectin
3.1.2. The Role of Transmission Efficiency Combined with Ivermectin Treatment in the Heterogeneous Scenario
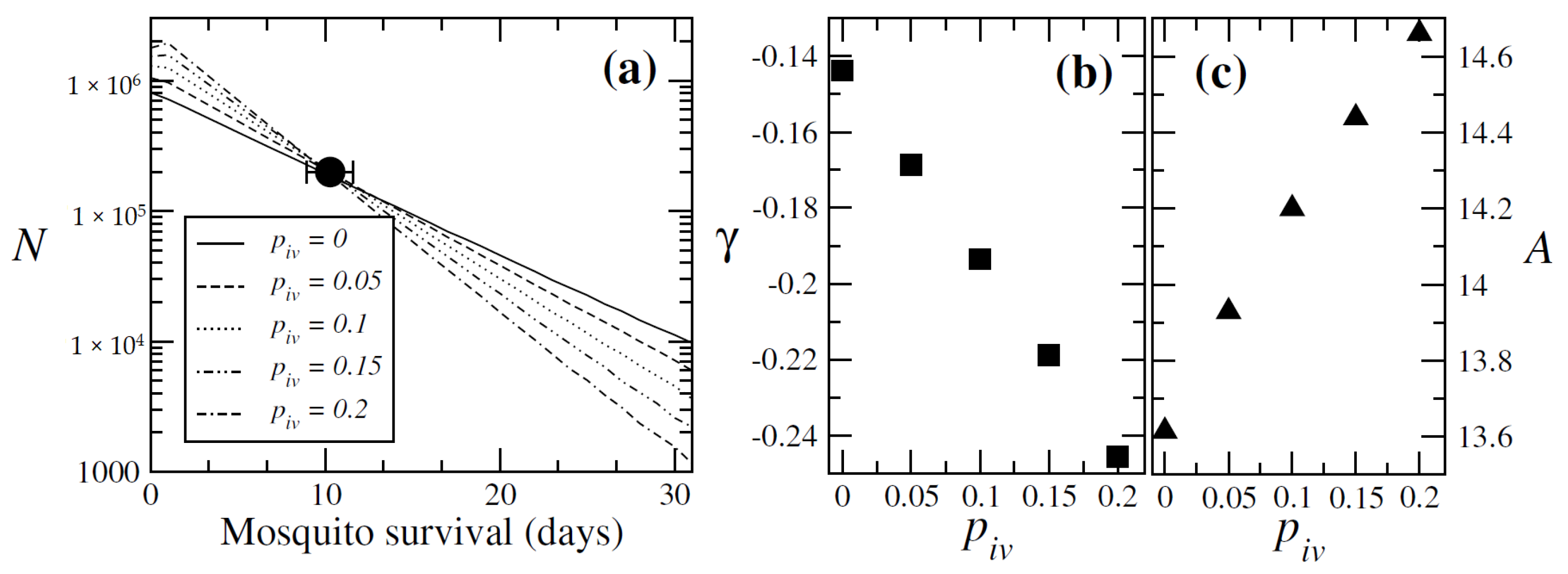
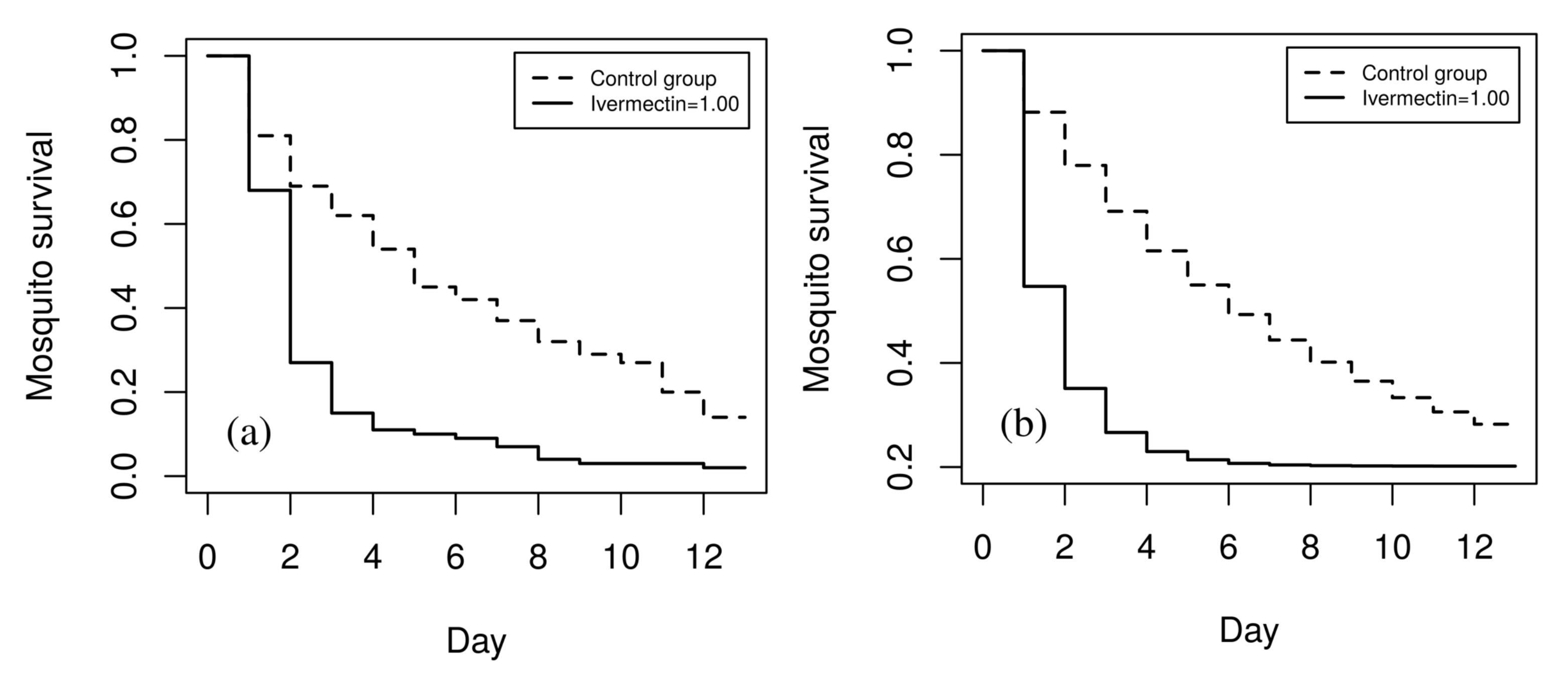
3.1.3. Mosquito Survival Patterns
3.1.4. Validation of the Ivermectin Parameter
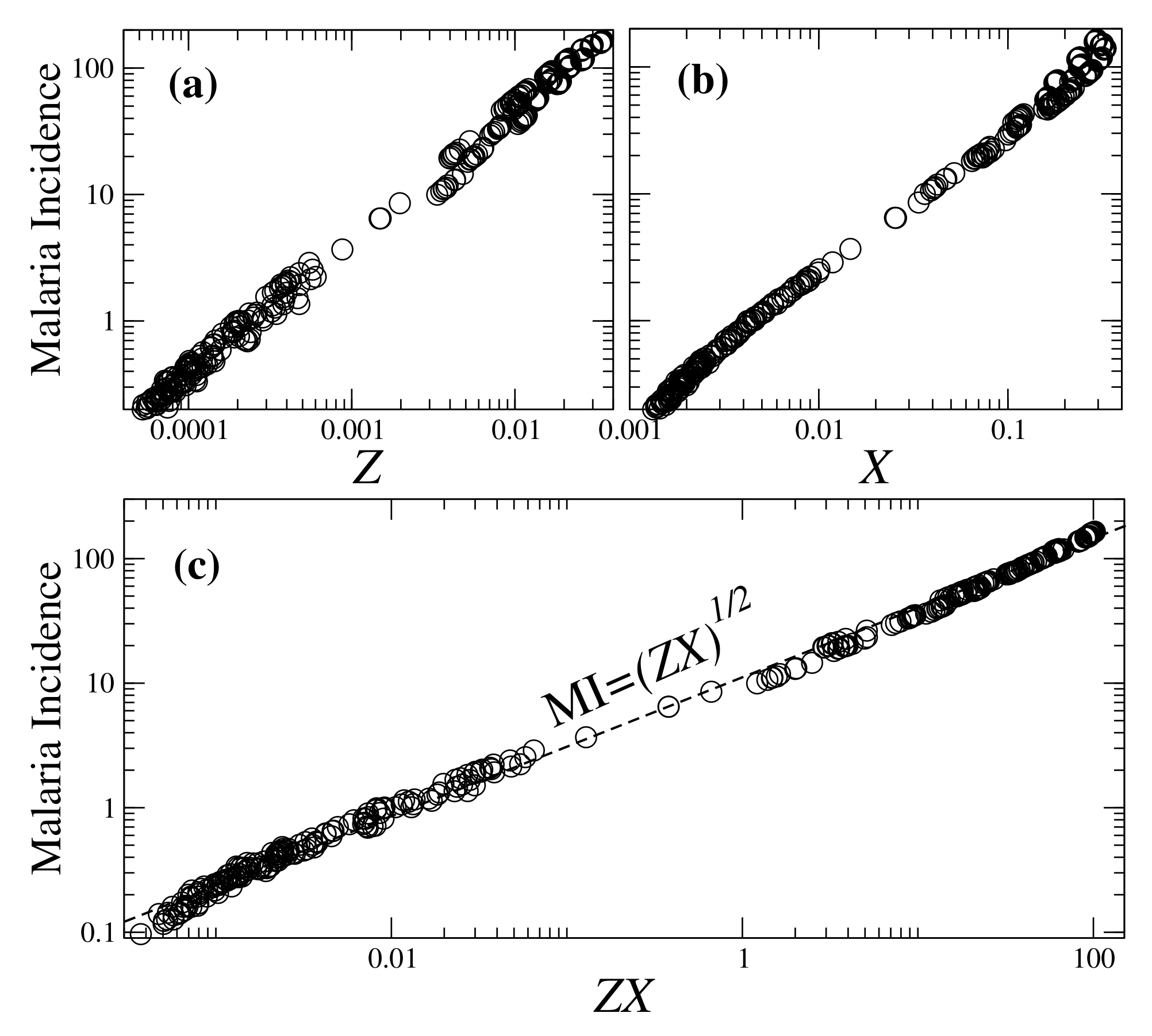
3.2. Towards Parameters for Assessing Malaria Incidence
Utility of Plasmodium Infection Metrics
3.3. Performance of Classical Ross–Macdonald Parameters in Heterogeneity-Driven Scenarios
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNICEF/WHO. Reversing the Incidence of Malaria 2000–2015. In WHO Global Malaria Programme; World Health Organization: Geneva, Switzerland, 2015; pp. 1–40. [Google Scholar]
- World Health Organization. World Malaria Report; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Acevedo, M.A.; Prosper, O.; Lopiano, K.; Ruktanonchai, N.; Caughlin, T.T.; Martcheva, M.; Osenberg, C.W.; Smith, D.L. Spatial heterogeneity, host movement and mosquito-borne disease transmission. PLoS ONE 2015, 10, e0127552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malede, A.; Alemu, K.; Aemero, M.; Robele, S.; Kloos, H. Travel to farms in the lowlands and inadequate malaria information significantly predict malaria in villages around Lake Tana, northwest Ethiopia: A matched case-control study. Malar. J. 2018, 17, 290. [Google Scholar] [CrossRef] [PubMed]
- Mukabana, W.R.; Takken, W.; Killeen, G.F.; Knols, B.G.J. Allomonal effect of breath contributes to differential attractiveness of humans to the African malaria vector Anopheles gambiae. Malar. J. 2004, 3, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.L.; McKenzie, F.E.; Snow, R.W.; Hay, S.I. Revisiting the basic reproductive number for malaria and its implications for malaria control. PLoS Biol. 2007, 5, e42. [Google Scholar] [CrossRef] [PubMed]
- Van der Waal, K.L.; Ezenwa, V.O. Heterogeneity in pathogen transmission: Mechanisms and methodology. Funct. Ecol. 2016, 30, 1606–1622. [Google Scholar] [CrossRef]
- Cooper, L.; Kang, S.Y.; Bisanzio, D.; Maxwell, K.; Rodriguez-Barraquer, I.; Greenhouse, B.; Drakeley, C.; Arinaitwe, E.; Staedke, S.G.; Gething, P.W.; et al. Pareto rules for malaria super-spreaders and super-spreading. Nat. Commun. 2019, 10, 1–9. [Google Scholar]
- Obala, A.A.; Kutima, H.L.; Nyamogoba, H.D.N.; Mwangi, A.W.; Simiyu, C.J.; Magak, G.N.; Khwa-Otsyula, B.O.; Ouma, J.H. Anopheles gambiae and Anopheles arabiensis population densities and infectivity in Kopere village, Western Kenya. J. Infect. Dev. Ctries. 2012, 6, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Dekker, T.; Takken, W.; Knols, B.G.J.; Bouman, E.; van de Laak, S.; de Bever, A.; Huisman, P.W.T. Selection of biting sites on a human host by Anopheles gambiae s. s., An. arabiensis and An. quadriannulatus. Entomol. Exp. Appl. 1998, 87, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Mahande, A.; Mosha, F.; Mahande, J.; Kweka, E. Feeding and resting behavior of malaria vector, Anopheles arabiensis with reference to zooprophylaxis. Malar. J. 2007, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Akogbéto, M.C.; Salako, A.S.; Dagnon, F.; Aikpon, R.; Kouletio, M.; Sovi, A.; Sezonlin, M. Blood feeding behavior comparison and contribution of Anopheles coluzzii and Anopheles gambiae, two sibling species living in sympatry, to malaria transmission in Alibori and Donga region, northern Benin, West Africa. Malar. J. 2018, 17, 307. [Google Scholar] [CrossRef]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Both infected and uninfected mosquitoes are attracted toward malaria infected birds. Malar. J. 2013, 12, 179. [Google Scholar] [CrossRef] [Green Version]
- De Moraes, C.M.; Stanczyk, N.M.; Betz, H.S.; Pulido, H.; Sim, D.G.; Read, A.F.; Mescher, M.C. Malaria-induced changes in host odors enhance mosquito attraction. Proc. Natl. Acad. Sci. USA 2014, 111, 11079–11084. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, R.; Mukabana, W.R.; Gouagna, L.C.; Koella, J.C. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol. 2005, 3, 1590–1593. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.E.; Dye, C.; Etard, J.F.; Smith, T.; Charlwood, J.D.; Garnett, G.P.; Hagan, P.; Hii, J.L.K.; Ndhlovu, P.D.; Quinnell, R.J.; et al. Heterogeneities in the transmission of infectious agents: Implications for the design of control programs. Proc. Natl. Acad. Sci. USA 1997, 94, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Pareto, V. Manuale di Economia Politica con una Introduzione Alla Scienza Sociale; Società Editrice Libraria: Torino, Italy, 1919; Volume 13. [Google Scholar]
- Brady, O.J.; Slater, H.C.; Pemberton-Ross, P.; Wenger, E.; Maude, R.J.; Ghani, A.C.; Penny, M.A.; Gerardin, J.; White, L.J.; Chitnis, M.; et al. Role of mass drug administration in elimination of Plasmodium falciparum malaria: A consensus modelling study. Lancet Glob. Health 2017, 5, e680–e687. [Google Scholar] [CrossRef] [Green Version]
- Bousema, T.; Drakeley, C.; Gesase, S.; Hashim, R.; Magesa, S.; Mosha, F.; Otieno, S.; Carneiro, I.; Cox, J.; Msuya, E.; et al. Identification of hot spots of malaria transmission for targeted malaria control. J. Infect. Dis. 2010, 201, 1764–1774. [Google Scholar] [CrossRef] [Green Version]
- Bousema, T.; Griffin, J.T.; Sauerwein, R.W.; Smith, D.L.; Churcher, T.S.; Takken, W.; Ghani, A.; Drakeley, C.; Gosling, R. Hitting hotspots: Spatial targeting of malaria for control and elimination. PLoS Med. 2012, 9, e1001165. [Google Scholar] [CrossRef] [Green Version]
- Noé, A.; Zaman, S.I.; Rahman, M.; Saha, A.K.; Aktaruzzaman, M.M.; Maude, R.J. Mapping the stability of malaria hotspots in Bangladesh from 2013 to 2016. Malar. J. 2018, 17, 259. [Google Scholar] [CrossRef] [Green Version]
- Chaccour, C.; Lines, J.; Whitty, C.J.M. Effect of Ivermectin on Anopheles gambiae Mosquitoes Fed on Humans: The Potential of Oral Insecticides in Malaria Control. J. Infect. Dis. 2010, 202, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Slater, H.C.; Foy, B.D.; Kobylinski, K.; Chaccour, C.; Watson, O.J.; Hellewell, J.; Aljayyoussi, G.; Bousema, T.; Burrows, J.; D’Alessandro, U. Ivermectin as a novel complementary malaria control tool to reduce incidence and prevalence: A modelling study. Lancet Infect. Dis. 2020, 20, P498–P508. [Google Scholar] [CrossRef]
- Sequeira, J.; Louçã, J.; Mendes, A.M.; Lind, P.G. Transition from endemic behavior to eradication of malaria due to combined drug therapies: An agent-model approach. J. Theor. Biol. 2020, 484, 110030. [Google Scholar] [CrossRef]
- Macdonald, G. The analysis of equilibrium in malaria. Trop. Dis. Bull. 1952, 49, 813–829. [Google Scholar]
- Ross, R. Some a Priori Pathometric Equations. Br. Med. J. 1915, 1, 546–547. [Google Scholar] [CrossRef] [Green Version]
- Biggs, J.; Raman, J.; Cook, J.; Hlongwana, K.; Drakeley, C.; Morris, N.; Serocharan, I.; Agubuzo, E.; Kruger, P.; Mabuza, A.; et al. Serology reveals heterogeneity of Plasmodium falciparum transmission in northeastern South Africa: Implications for malaria elimination. Malar. J. 2017, 16, 48. [Google Scholar] [CrossRef] [Green Version]
- Dye, C.; Hasibeder, G. Population dynamics of mosquito-borne disease: Effects of flies which bite some people more frequently than others. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 69–77. [Google Scholar] [CrossRef]
- Alemu, K.; Worku, A.; Berhane, Y.; Kumie, A. Spatiotemporal clusters of malaria cases at village level, northwest Ethiopia. Malar. J. 2014, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Camarda, G.; Jirawatcharadech, P.; Priestley, R.S.; Saif, A.; March, S.; Wong, M.H.; Leung, S.; Miller, A.B.; Baker, D.A.; Alano, P.; et al. Antimalarial activity of primaquine operates via a two-step biochemical relay. Nat. Commun. 2019, 10, 3226. [Google Scholar] [CrossRef]
- Lawpoolsri, S.; Klein, E.Y.; Singhasivanon, P.; Yimsamran, S.; Thanyavanich, N.; Maneeboonyang, W.; Hungerford, L.L.; Maguire, J.H.; Smith, D.L. Optimally timing primaquine treatment to reduce Plasmodium falciparum transmission in low endemicity Thai-Myanmar border populations. Malar. J. 2009, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Shekalaghe, S.; Mosha, D.; Hamad, A.; Mbaga, T.A.; Mihayo, M.; Bousema, T.; Drakeley, C.; Abdulla, S. Optimal timing of primaquine to reduce Plasmodium falciparum gametocyte carriage when co-administered with artemether–lumefantrine. Malar. J. 2020, 19, 34. [Google Scholar] [CrossRef]
- Bosson-Vanga, H.; Franetich, J.F.; Soulard, V.; Sossau, D.; Tefit, M.; Kane, B.; Vaillant, J.C.; Borrmann, S.; Müller, O.; Dereuddre-Bosquet, N.; et al. Differential activity of methylene blue against erythrocytic and hepatic stages of Plasmodium. Malar. J. 2018, 17, 143. [Google Scholar] [CrossRef]
- Mendes, A.M.; Albuquerque, I.S.; Machado, M.; Pissarra, J.; Meireles, P.; Prudêncio, M. Inhibition of Plasmodium liver infection by ivermectin. Antimicrob. Agents Chemother. 2017, 61, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobylinski, K.C.; Foy, B.D.; Richardson, J.H. Ivermectin inhibits the sporogony of Plasmodium falciparum in Anopheles gambiae. Malar. J. 2012, 11, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaccour, C.; Rabinovich, N.R. Advancing the repurposing of ivermectin for malaria. Lancet 2019, 393, 1480–1481. [Google Scholar] [CrossRef] [Green Version]
- Smit, M.R.; Ochomo, E.O.; Aljayyoussi, G.; Kwambai, T.K.; Abong’o, B.O.; Chen, T.; Bousema, T.; Slater, H.C.; Waterhouse, D.; Bayoh, N.M. Safety and mosquitocidal efficacy of high-dose ivermectin when co-administered with dihydroartemisinin-piperaquine in Kenyan adults with uncomplicated malaria (IVERMAL): A randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2018, 18, P615–P626. [Google Scholar] [CrossRef]
- Foy, B.D.; Alout, H.; Seaman, J.A.; Rao, S.; Magalhaes, T.; Wade, M.; Parikh, S.; Soma, D.D.; Sagna, A.D.; Fournet, F. Efficacy and risk of harms of repeat ivermectin mass drug administrations for control of malaria (RIMDAMAL): A cluster-randomised trial. Lancet 2019, 393, P1517–P1526. [Google Scholar] [CrossRef] [Green Version]
- Ouédraogo, A.L.; Bastiaens, G.J.; Tiono, A.B.; Guelbéogo, W.M.; Kobylinski, K.C.; Ouédraogo, A.; Barry, A.; Bougouma, E.C.; Nebie, I.; Ouattara, M.S.; et al. Efficacy and safety of the mosquitocidal drug ivermectin to prevent malaria transmission after treatment: A double-blind, randomized, clinical trial. Clin. Infect. Dis. 2015, 60, 357–365. [Google Scholar] [CrossRef]
- Azevedo, R.; Mendes, A.M.; Prudêncio, M. Inhibition of Plasmodium sporogonic stages by ivermectin and other avermectins. Parasites Vectors 2019, 12, 549. [Google Scholar] [CrossRef] [Green Version]
- Stresman, G.H.; Giorgi, E.; Baidjoe, A.; Knight, P.; Odongo, W.; Owaga, C.; Shagari, S.; Makori, E.; Stevenson, J.; Drakeley, C.; et al. Impact of metric and sample size on determining malaria hotspot boundaries. Sci. Rep. 2017, 7, 45849. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.L.; McKenzie, F.E. Statics and dynamics of malaria infection in Anopheles mosquitoes. Malar. J. 2004, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.P.; Bukirwa, H.; Njama-Meya, D.; Francis, D.; Kamya, M.R.; Rosenthal, P.J.; Dorsey, G. Use of the slide positivity rate to estimate changes in malaria incidence in a cohort of Ugandan children. Malar. J. 2009, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Boyce, R.M.; Reyes, R.; Matte, M.; Ntaro, M.; Mulogo, E.; Lin, F.C.; Siedner, M.J. Practical implications of the non-linear relationship between the test positivity rate and malaria incidence. PLoS ONE 2016, 11, e0152410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Hu, W.; Liu, H.; Xiao, Y.; Guo, Y.; Chen, S.; Zhao, L.; Tong, S. Can slide positivity rates predict malaria transmission? Malar. J. 2012, 11, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pombi, M.; Calzetta, M.; Guelbeogo, W.M.; Manica, M.; Perugini, E.; Pichler, V.; Mancini, E.; Sagnon, N.F.; Ranson, H.; Della Torre, A. Unexpectedly high Plasmodium sporozoite rate associated with low human blood index in Anopheles coluzzii from a LLIN-protected village in Burkina Faso. Sci. Rep. 2018, 8, 12806. [Google Scholar] [CrossRef] [PubMed]
- Bejon, P.; Williams, T.N.; Liljander, A.; Noor, A.M.; Wambua, J.; Ogada, E.; Olotu, A.; Osier, F.H.; Hay, S.I.; Färnert, A. Stable and unstable malaria hotspots in longitudinal cohort studies in Kenya. PLoS Med. 2010, 7, e1000304. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sequeira, J.; Louçã, J.; Mendes, A.M.; Lind, P.G. A Model for Assessing the Quantitative Effects of Heterogeneous Affinity in Malaria Transmission along with Ivermectin Mass Administration. Appl. Sci. 2020, 10, 8696. https://doi.org/10.3390/app10238696
Sequeira J, Louçã J, Mendes AM, Lind PG. A Model for Assessing the Quantitative Effects of Heterogeneous Affinity in Malaria Transmission along with Ivermectin Mass Administration. Applied Sciences. 2020; 10(23):8696. https://doi.org/10.3390/app10238696
Chicago/Turabian StyleSequeira, João, Jorge Louçã, António M. Mendes, and Pedro G. Lind. 2020. "A Model for Assessing the Quantitative Effects of Heterogeneous Affinity in Malaria Transmission along with Ivermectin Mass Administration" Applied Sciences 10, no. 23: 8696. https://doi.org/10.3390/app10238696
APA StyleSequeira, J., Louçã, J., Mendes, A. M., & Lind, P. G. (2020). A Model for Assessing the Quantitative Effects of Heterogeneous Affinity in Malaria Transmission along with Ivermectin Mass Administration. Applied Sciences, 10(23), 8696. https://doi.org/10.3390/app10238696