Exercise Training of Secreted Protein Acidic and Rich in Cysteine (Sparc) KO Mice Suggests That Exercise-Induced Muscle Phenotype Changes Are SPARC-Dependent



Abstract
:Featured Application
Abstract
1. Secreted Protein Acidic and Rich in Cysteine as an Exercise-Induced Gene
2. Animal Experimental Design, Material and Methods
2.1. Mice Exercise Protocol and Running Speed Determination
2.2. Fasting Lactate and Oral Glucose Gavage-Dependant Lactate Levels
2.3. Grip Power Test
2.4. Body and Tissue Weights
2.5. Western Blotting
2.6. Statistical Analyses and Sample Size Determination
3. Results
3.1. Exercise Patterns, Running Speed and Lactate Concentrations
3.2. Body and Tissue Weights
3.3. Muscle Strength (Grip Power Tests)
3.4. COL1A1 and MT-CO1 Expressions in Tibialis Anterior Muscle
4. Discussion and Interpretation
4.1. Lactate Concentrations among the Indicators of Muscles Metabolic Performance
4.2. Body and Tissue Weights (Table 3)
4.3. Protein Expressions (Figure 1) and Muscle Strength (Table 4)
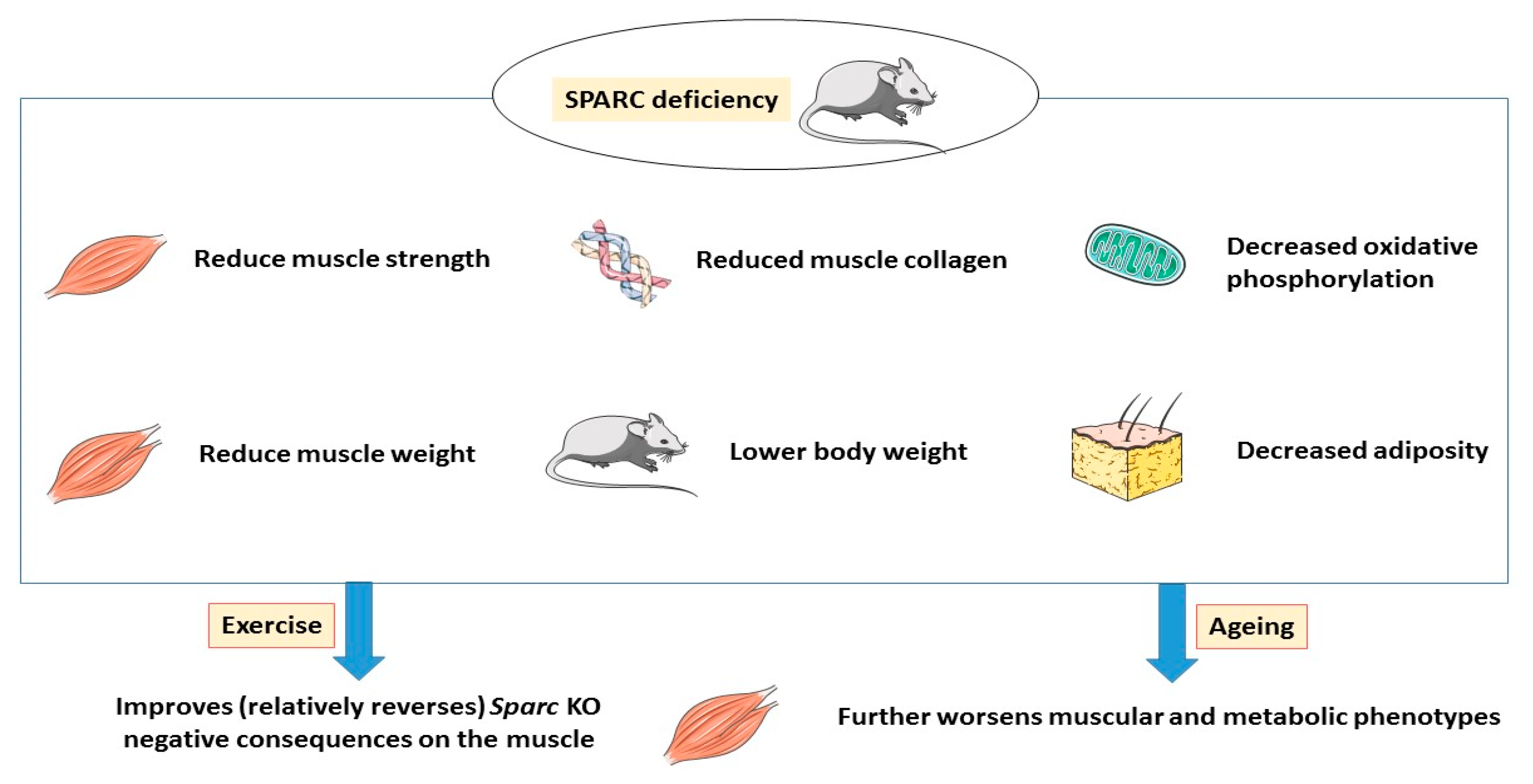
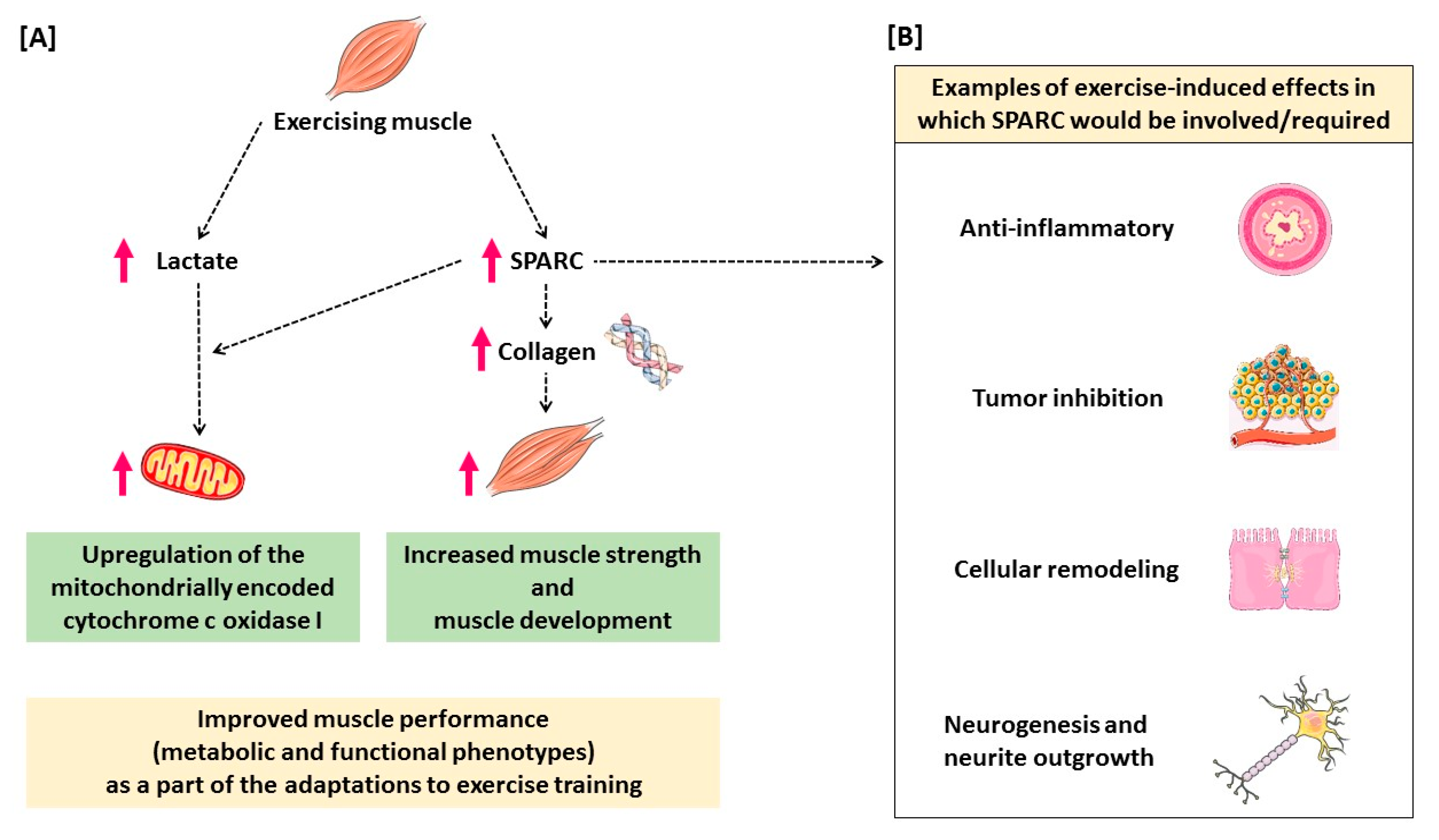
5. Conclusions and Hypothetical Mechanisms
6. Implications and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pedersen, B.K. The physiology of optimizing health with a focus on exercise as medicine. Annu. Rev. Physiol. 2019, 81, 607–627. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W.; Sakuma, K. Skeletal muscle: Novel and intriguing characteristics as a secretory organ. BioDiscovery 2013, 7, e8942. [Google Scholar] [CrossRef]
- Ghanemi, A.; Melouane, A.; Yoshioka, M.; St-Amand, J. Exercise and high-fat diet in obesity: Functional genomics perspectives of two energy homeostasis pillars. Genes 2020, 11, 875. [Google Scholar] [CrossRef] [PubMed]
- Berryhill, B.L.; Kane, B.; Stramer, B.M.; Fini, M.E.; Hassell, J.R. Increased SPARC accumulation during corneal repair. Exp. Eye Res. 2003, 77, 85–92. [Google Scholar] [CrossRef]
- Phan, E.; Ahluwalia, A.; Tarnawski, A.S. Role of SPARC—Matricellular protein in pathophysiology and tissue injury healing. Implications for gastritis and gastric ulcers. Med. Sci. Monit. 2007, 13, RA25–RA30. [Google Scholar]
- Riedl, I.; Yoshioka, M.; Nishida, Y.; Tobina, T.; Paradis, R.; Shono, N.; Tanaka, H.; St-Amand, J. Regulation of skeletal muscle transcriptome in elderly men after 6 weeks of endurance training at lactate threshold intensity. Exp. Gerontol. 2010, 45, 896–903. [Google Scholar] [CrossRef]
- Melouane, A.; Yoshioka, M.; Kanzaki, M.; St-Amand, J. Sparc, an EPS-induced gene, modulates the extracellular matrix and mitochondrial function via ILK/AMPK pathways in C2C12 cells. Life Sci. 2019, 229, 277–287. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Takagi, T.; Tanimura, Y.; Takanami, Y.; Kawai, Y.; Sakuma, K.; Hang, L.P.; Mizushima, K.; Hirai, Y.; et al. A novel myokine, secreted protein acidic and rich in cysteine (SPARC), suppresses colon tumorigenesis via regular exercise. Gut 2013, 62, 882–889. [Google Scholar] [CrossRef]
- Matsuo, K.; Sato, K.; Suemoto, K.; Miyamoto-Mikami, E.; Fuku, N.; Higashida, K.; Tsuji, K.; Xu, Y.; Liu, X.; Iemitsu, M.; et al. A mechanism underlying preventive effect of high-intensity training on colon cancer. Med. Sci. Sports Exerc. 2017, 49, 1805–1816. [Google Scholar] [CrossRef]
- Norheim, F.; Raastad, T.; Thiede, B.; Rustan, A.C.; Drevon, C.A.; Haugen, F. Proteomic identification of secreted proteins from human skeletal muscle cells and expression in response to strength training. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1013–E1021. [Google Scholar] [CrossRef] [Green Version]
- Garneau, L.; Parsons, S.A.; Smith, S.R.; Mulvihill, E.E.; Sparks, L.M.; Aguer, C. Plasma myokine concentrations after acute exercise in non-obese and obese sedentary women. Front. Physiol. 2020, 11, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billat, V.L.; Mouisel, E.; Roblot, N.; Melki, J. Inter- and intrastrain variation in mouse critical running speed. J. Appl. Physiol. (1985) 2005, 98, 1258–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://insights.envigo.com/hubfs/resources/data-sheets/2018s-datasheet-0915.pdf (accessed on 9 August 2020).
- Norose, K.; Lo, W.K.; Clark, J.I.; Sage, E.H.; Howe, C.C. Lenses of SPARC-null mice exhibit an abnormal cell surface-basement membrane interface. Exp. Eye Res. 2000, 71, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Bradshaw, A.D.; Delany, A.M.; Sage, E.H. Inactivation of SPARC enhances high-fat diet-induced obesity in mice. Connect. Tissue Res. 2011, 52, 99–108. [Google Scholar] [CrossRef]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate threshold concepts: How valid are they? Sports Med. 2009, 39, 469–490. [Google Scholar] [CrossRef]
- Schefer, V.; Talan, M.I. Oxygen consumption in adult and AGED C57BL/6J mice during acute treadmill exercise of different intensity. Exp. Gerontol. 1996, 31, 387–392. [Google Scholar] [CrossRef]
- Nishida, Y.; Tokuyama, K.; Nagasaka, S.; Higaki, Y.; Shirai, Y.; Kiyonaga, A.; Shindo, M.; Kusaka, I.; Nakamura, T.; Ishibashi, S.; et al. Effect of moderate exercise training on peripheral glucose effectiveness, insulin sensitivity, and endogenous glucose production in healthy humans estimated by a two-compartment-labeled minimal model. Diabetes 2004, 53, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Shindo, M. The benefits of the low intensity training. Ann. Physiol. Anthr. 1992, 11, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Motoyama, M.; Sunami, Y.; Kinoshita, F.; Kiyonaga, A.; Tanaka, H.; Shindo, M.; Irie, T.; Urata, H.; Sasaki, J.; Arakawa, K. Blood pressure lowering effect of low intensity aerobic training in elderly hypertensive patients. Med. Sci. Sports Exerc. 1998, 30, 818–823. [Google Scholar]
- Sunami, Y.; Motoyama, M.; Kinoshita, F.; Mizooka, Y.; Sueta, K.; Matsunaga, A.; Sasaki, J.; Tanaka, H.; Shindo, M. Effects of low-intensity aerobic training on the high-density lipoprotein cholesterol concentration in healthy elderly subjects. Metabolism 1999, 48, 984–988. [Google Scholar] [CrossRef]
- Kawanishi, N.; Yano, H.; Mizokami, T.; Takahashi, M.; Oyanagi, E.; Suzuki, K. Exercise training attenuates hepatic inflammation, fibrosis and macrophage infiltration during diet induced-obesity in mice. Brain Behav. Immun. 2012, 26, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Shubert, T.; Zhao, C.; Gage, F.H. Exercise enhances learning and hippocampal neurogenesis in aged mice. J. Neurosci. 2005, 25, 8680–8685. [Google Scholar] [CrossRef] [PubMed]
- Rowlatt, C.; Chesterman, F.C.; Sheriff, M.U. Lifespan, age changes and tumour incidence in an ageing C57BL mouse colony. Lab. Anim. 1976, 10, 419–442. [Google Scholar] [CrossRef] [PubMed]
- Kunstyr, I.; Leuenberger, H.G. Gerontological data of C57BL/6J mice. I. Sex differences in survival curves. J. Gerontol. 1975, 30, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.P.; Hicks, P.S.; Ortiz, L.R.; Martinez, M.J.; Mandler, R.N. Quantitative measurement of muscle strength in the mouse. J. Neurosci. Methods 1995, 62, 15–19. [Google Scholar] [CrossRef]
- Takeshita, H.; Yamamoto, K.; Nozato, S.; Inagaki, T.; Tsuchimochi, H.; Shirai, M.; Yamamoto, R.; Imaizumi, Y.; Hongyo, K.; Yokoyama, S.; et al. Modified forelimb grip strength test detects aging-associated physiological decline in skeletal muscle function in male mice. Sci. Rep. 2017, 7, 42323. [Google Scholar] [CrossRef]
- Melouane, A.; Carbonell, A.; Yoshioka, M.; Puymirat, J.; St-Amand, J. Implication of SPARC in the modulation of the extracellular matrix and mitochondrial function in muscle cells. PLoS ONE 2018, 13, e0192714. [Google Scholar] [CrossRef] [Green Version]
- Zong, S.; Wu, M.; Gu, J.; Liu, T.; Guo, R.; Yang, M. Structure of the intact 14-subunit human cytochrome c oxidase. Cell Res. 2018, 28, 1026–1034. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Taylor, S.C.; Berkelman, T.; Yadav, G.; Hammond, M. A defined methodology for reliable quantification of Western blot data. Mol. Biotechnol. 2013, 55, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.C.; Posch, A. The design of a quantitative western blot experiment. Biomed. Res. Int. 2014, 2014, 361590. [Google Scholar] [CrossRef] [PubMed]
- Ghanemi, A.; Melouane, A.; Mucunguzi, O.; Yoshioka, M.; St-Amand, J. Energy and metabolic pathways in trefoil factor family member 2 (Tff2) KO mice beyond the protection from high-fat diet-induced obesity. Life Sci. 2018, 215, 190–197. [Google Scholar] [CrossRef]
- De Giorgio, M.R.; Yoshioka, M.; Riedl, I.; Moreault, O.; Cherizol, R.G.; Shah, A.A.; Blin, N.; Richard, D.; StAmand, J. Trefoil factor family member 2 (Tff2) KO mice are protected from high-fat diet-induced obesity. Obesity (Silver Spring) 2013, 21, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D.; Sage, E.H. SPARC, a matricellular protein that functions in cellular differentiation and tissue response to injury. J. Clin. Investig. 2001, 107, 1049–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brekken, R.A.; Sage, E.H. SPARC, a matricellular protein: At the crossroads of cell-matrix communication. Matrix Biol. 2001, 19, 816–827. [Google Scholar] [CrossRef]
- Lane, T.F.; Sage, E.H. The biology of SPARC, a protein that modulates cell-matrix interactions. FASEB J. 1994, 8, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Sage, E.H. SPARC, a matricellular glycoprotein with important biological functions. J. Histochem. Cytochem. 1999, 47, 1495–1506. [Google Scholar] [CrossRef] [Green Version]
- Mason, I.J.; Murphy, D.; Münke, M.; Francke, U.; Elliott, R.W.; Hogan, B.L. Developmental and transformation-sensitive expression of the Sparc gene on mouse chromosome 11. EMBO J. 1986, 5, 1831–1837. [Google Scholar] [CrossRef]
- Swaroop, A.; Hogan, B.L.; Francke, U. Molecular analysis of the cDNA for human SPARC/osteonectin/BM-40: Sequence, expression, and localization of the gene to chromosome 5q31-q33. Genomics 1988, 2, 37–47. [Google Scholar] [CrossRef]
- Brekken, R.A.; Sage, E.H. SPARC, a matricellular protein: At the crossroads of cell-matrix. Matrix Biol. 2000, 19, 569–580. [Google Scholar] [CrossRef]
- Francki, A.; Motamed, K.; McClure, T.D.; Kaya, M.; Murri, C.; Blake, D.J.; Carbon, J.G.; Sage, E.H. SPARC regulates cell cycle progression in mesangial cells via its inhibition of IGF-dependent signaling. J. Cell. Biochem. 2003, 88, 802–811. [Google Scholar] [CrossRef]
- Basu, A.; Kligman, L.H.; Samulewicz, S.J.; Howe, C.C. Impaired wound healing in mice deficient in a matricellular protein SPARC (osteonectin, BM-40). BMC Cell Biol. 2001, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Ghanemi, A.; Melouane, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and bioenergetics: Extracellular matrix, adipocytes remodeling and skeletal muscle metabolism. Int. J. Biochem. Cell Biol. 2019, 117, 105627. [Google Scholar] [CrossRef] [PubMed]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine: Metabolic and homeostatic properties beyond the extracellular matrix structure. Appl. Sci. 2020, 10, 2388. [Google Scholar] [CrossRef] [Green Version]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and cancer: A homeostatic hormone? Cytokine 2020, 127, 154996. [Google Scholar] [CrossRef] [PubMed]
- Ghanemi, A.; Yoshioka, M.; St-Amand, J. Secreted protein acidic and rich in cysteine and inflammation: Another homeostatic property? Cytokine 2020, 133, 155179. [Google Scholar] [CrossRef]
- Jørgensen, L.H.; Petersson, S.J.; Sellathurai, J.; Andersen, D.C.; Thayssen, S.; Sant, D.J.; Jensen, C.H.; Schrøder, H.D. Secreted protein acidic and rich in cysteine (SPARC) in human skeletal muscle. J. Histochem. Cytochem. 2009, 57, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.W.; Harper, S.J.; McVey, J.H.; Hogan, B.L. In vivo expression of mRNA for the Ca++-binding protein SPARC (osteonectin) revealed by in situ hybridization. J. Cell Biol. 1987, 105, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Sage, H.; Vernon, R.B.; Decker, J.; Funk, S.; Iruela-Arispe, M.L. Distribution of the calcium-binding protein SPARC in tissues of embryonic and adult mice. J. Histochem. Cytochem. 1989, 37, 819–829. [Google Scholar] [CrossRef]
- Framson, P.E.; Sage, E.H. SPARC and tumor growth: Where the seed meets the soil? J. Cell. Biochem. 2004, 92, 679–690. [Google Scholar] [CrossRef]
- Holloszy, J.O. Muscle metabolism during exercise. Arch. Phys. Med. Rehabil. 1982, 63, 231–234. [Google Scholar] [PubMed]
- Ferraro, E.; Giammarioli, A.M.; Chiandotto, S.; Spoletini, I.; Rosano, G. Exercise-induced skeletal muscle remodeling and metabolic adaptation: Redox signaling and role of autophagy. Antioxid. Redox Signal. 2014, 21, 154–176. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, L.H.; Jepsen, P.L.; Boysen, A.; Dalgaard, L.B.; Hvid, L.G.; Ørtenblad, N.; Ravn, D.; Sellathurai, J.; Møller-Jensen, J.; Lochmüller, H.; et al. SPARC interacts with actin in skeletal muscle in vitro and in vivo. Am. J. Pathol. 2017, 187, 457–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chlenski, A.; Guerrero, L.J.; Salwen, H.R.; Yang, Q.; Tian, Y.; La Madrid, A.M.; Mirzoeva, S.; Bouyer, P.G.; Xu, D.; Walker, M.; et al. Secreted protein acidic and rich in cysteine is a matrix scavenger chaperone. PLoS ONE 2011, 6, e23880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delany, A.M.; Amling, M.; Priemel, M.; Howe, C.; Baron, R.; Canalis, E. Osteopenia and decreased bone formation in osteonectin-deficient mice. J. Clin. Investig. 2000, 105, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Fertala, A.; Ratner, B.D.; Sage, E.H.; Jiang, S. Identifying the SPARC binding sites on collagen I and procollagen I by atomic force microscopy. Anal. Chem. 2005, 77, 6765–6771. [Google Scholar] [CrossRef]
- Giudici, C.; Raynal, N.; Wiedemann, H.; Cabral, W.A.; Marini, J.C.; Timpl, R.; Bächinger, H.P.; Farndale, R.W.; Sasaki, T.; Tenni, R. Mapping of SPARC/BM-40/osteonectin-binding sites on fibrillar collagens. J. Biol. Chem. 2008, 283, 19551–19560. [Google Scholar] [CrossRef] [Green Version]
- Omi, S.; Yamanouchi, K.; Nakamura, K.; Matsuwaki, T.; Nishihara, M. Reduced fibrillar collagen accumulation in skeletal muscle of secreted protein acidic and rich in cysteine (SPARC)-null mice. J. Vet. Med. Sci. 2019, 81, 1649–1654. [Google Scholar] [CrossRef] [Green Version]
- Sage, H.; Vernon, R.B.; Funk, S.E.; Everitt, E.A.; Angello, J. SPARC, a secreted protein associated with cellular proliferation, inhibits cell spreading in vitro and exhibits Ca+2-dependent binding to the extracellular matrix. J. Cell Biol. 1989, 109, 341–356. [Google Scholar] [CrossRef]
- Francki, A.; Bradshaw, A.D.; Bassuk, J.A.; Howe, C.C.; Couser, W.G.; Sage, E.H. SPARC regulates the expression of collagen type I and transforming growth factor-beta1 in mesangial cells. J. Biol. Chem. 1999, 274, 32145–32152. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, A.D.; Reed, M.J.; Sage, E.H. SPARC-null mice exhibit accelerated cutaneous wound closure. J. Histochem. Cytochem. 2002, 50, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melouane, A.; Yoshioka, M.; St-Amand, J. Extracellular matrix/mitochondria pathway: A novel potential target for sarcopenia. Mitochondrion 2020, 50, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Guan, Y.; Zhang, L.; Li, K.; Dong, C. SPARC interacts with AMPK and regulates GLUT4 expression. Biochem. Biophys. Res. Commun. 2010, 396, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Lira, V.A.; Benton, C.R.; Yan, Z.; Bonen, A. PGC-1alpha regulation by exercise training and its influences on muscle function and insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E145–E161. [Google Scholar] [CrossRef] [Green Version]
- Jäger, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. AMP-activated protein kinase (AMPK) action in skeletal muscle via direct phosphorylation of PGC-1alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.M.; Gallo, G. The role of mitochondria in axon development and regeneration. Dev. Neurobiol. 2018, 78, 221–237. [Google Scholar] [CrossRef]
- Han, S.M.; Baig, H.S.; Hammarlund, M. Mitochondria localize to injured axons to support regeneration. Neuron 2016, 92, 1308–1323. [Google Scholar] [CrossRef] [Green Version]
- Aoi, W.; Hirano, N.; Lassiter, D.G.; Björnholm, M.; Chibalin, A.V.; Sakuma, K.; Tanimura, Y.; Mizushima, K.; Takagi, T.; Naito, Y.; et al. Secreted protein acidic and rich in cysteine (SPARC) improves glucose tolerance via AMP-activated protein kinase activation. FASEB J. 2019, 33, 10551–10562. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, A.D. The role of secreted protein acidic and rich in cysteine (SPARC) in cardiac repair and fibrosis: Does expression of SPARC by macrophages influence outcomes? J. Mol. Cell. Cardiol. 2016, 93, 156–161. [Google Scholar] [CrossRef]
- McCurdy, S.; Baicu, C.F.; Heymans, S.; Bradshaw, A.D. Cardiac extracellular matrix remodeling: Fibrillar collagens and secreted protein acidic and rich in cysteine (SPARC). J. Mol. Cell. Cardiol. 2010, 48, 544–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Sun, X.; Guo, J.; Lu, G. SPARC support the expansion of cord blood stem cells in vitro. Cell Res. 2008, 18, S49. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wang, L.Y.; Li, C.Y.; Wu, J.Y.; Zhang, Y.T.; Pang, K.P.; Wei, Y.; Du, L.Q.; Liu, M.; Wu, X.Y. SPARC promotes self-renewal of limbal epithelial stem cells and ocular surface restoration through JNK and p38-MAPK signaling pathways. Stem Cells 2020, 38, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, K.; Trayhurn, P. Lactate and the GPR81 receptor in metabolic regulation: Implications for adipose tissue function and fatty acid utilisation by muscle during exercise. Br. J. Nutr. 2011, 106, 1310–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladden, L.B. Muscle as a consumer of lactate. Med. Sci. Sports Exerc. 2000, 32, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Gladden, L.B. Lactate metabolism: A new paradigm for the third millennium. J. Physiol. 2004, 558, 5–30. [Google Scholar] [CrossRef]
- Hashimoto, T.; Hussien, R.; Oommen, S.; Gohil, K.; Brooks, G.A. Lactate sensitive transcription factor network in L6 cells: Activation of MCT1 and mitochondrial biogenesis. FASEB J. 2007, 21, 2602–2612. [Google Scholar] [CrossRef]
- Hashimoto, T.; Brooks, G.A. Mitochondrial lactate oxidation complex and an adaptive role for lactate production. Med. Sci. Sports Exerc. 2008, 40, 486–494. [Google Scholar] [CrossRef]
- Gladden, L.B. A lactatic perspective on metabolism. Med. Sci. Sports Exerc. 2008, 40, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, I. Blood lactate. Sports Med. 1986, 3, 10–25. [Google Scholar] [CrossRef]
- Campolongo, M.; Benedetti, L.; Podhajcer, O.L.; Pitossi, F.; Depino, A.M. Hippocampal SPARC regulates depression-related behavior. Genes Brain Behav. 2012, 11, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Leveille, G.A.; Chakrabarty, K. Diurnal variations in tissue glycogen and liver weight of meal-fed rats. J. Nutr. 1967, 93, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Kreitzman, S.N.; Coxon, A.Y.; Szaz, K.F. Glycogen storage: Illusions of easy weight loss, excessive weight regain, and distortions in estimates of body composition. Am. J. Clin. Nutr. 1992, 56 (Suppl. 1), 292s–293s. [Google Scholar] [CrossRef]
- Ma, C.L.; Ma, X.T.; Wang, J.J.; Liu, H.; Chen, Y.F.; Yang, Y. Physical exercise induces hippocampal neurogenesis and prevents cognitive decline. Behav. Brain Res. 2017, 317, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.F.; Paes, F.; Arias-Carrión, O.; Ferreira Rocha, N.B.; de Sá Filho, A.S.; Machado, S. Neural mechanisms of exercise: Anti-depression, neurogenesis, and serotonin signaling. CNS Neurol. Disord. Drug Targets 2015, 14, 1307–1311. [Google Scholar] [CrossRef]
- Uddin, M.S.; Hossain, M.F.; Mamun, A.A.; Shah, M.A.; Hasana, S.; Bulbul, I.J.; Sarwar, M.S.; Mansouri, R.A.; Ashraf, G.M.; Rauf, A.; et al. Exploring the multimodal role of phytochemicals in the modulation of cellular signaling pathways to combat age-related neurodegeneration. Sci. Total Environ. 2020, 725, 138313. [Google Scholar] [CrossRef]
- Hung, C.W.; Chen, Y.C.; Hsieh, W.L.; Chiou, S.H.; Kao, C.L. Ageing and neurodegenerative diseases. Ageing Res. Rev. 2010, 9 (Suppl. 1), S36–S46. [Google Scholar] [CrossRef]
- Mahalakshmi, B.; Maurya, N.; Lee, S.-D.; Bharath Kumar, V. Possible neuroprotective mechanisms of physical exercise in neurodegeneration. Int. J. Mol. Sci. 2020, 21, 5895. [Google Scholar] [CrossRef]
- Ang, E.-T.; Tai, Y.-K.; Lo, S.-Q.; Seet, R.; Soong, T.-W. Neurodegenerative diseases: Exercising toward neurogenesis and neuroregeneration. Front. Aging Neurosci. 2010, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yan, T.; Chu, J.M.; Chen, Y.; Dunnett, S.; Ho, Y.S.; Wong, G.T.; Chang, R.C. The beneficial effects of physical exercise in the brain and related pathophysiological mechanisms in neurodegenerative diseases. Lab. Investig. 2019, 99, 943–957. [Google Scholar] [CrossRef]
- Pedersen, L.; Idorn, M.; Olofsson, G.H.; Lauenborg, B.; Nookaew, I.; Hansen, R.H.; Johannesen, H.H.; Becker, J.C.; Pedersen, K.S.; Dethlefsen, C.; et al. Voluntary running suppresses tumor growth through epinephrine- and IL-6-dependent NK cell mobilization and redistribution. Cell Metab. 2016, 23, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedell-Neergaard, A.S.; Lang Lehrskov, L.; Christensen, R.H.; Legaard, G.E.; Dorph, E.; Larsen, M.K.; Launbo, N.; Fagerlind, S.R.; Seide, S.K.; Nymand, S.; et al. Exercise-induced changes in visceral adipose tissue mass are regulated by IL-6 signaling: A randomized controlled trial. Cell Metab. 2019, 29, 844–855.e3. [Google Scholar] [CrossRef] [PubMed]
- Ellingsgaard, H.; Hojman, P.; Pedersen, B.K. Exercise and health—Emerging roles of IL-6. Curr. Opin. Physiol. 2019, 10, 49–54. [Google Scholar] [CrossRef]
- Sakuma, K.; Yamaguchi, A. Sarcopenic obesity and endocrinal adaptation with age. Int. J. Endocrinol. 2013, 2013, 204164. [Google Scholar] [CrossRef] [Green Version]
- Candow, D.G.; Chilibeck, P.D. Differences in size, strength, and power of upper and lower body muscle groups in young and older men. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Kalyani, R.R.; Corriere, M.; Ferrucci, L. Age-related and disease-related muscle loss: The effect of diabetes, obesity, and other diseases. Lancet Diabetes Endocrinol. 2014, 2, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.N.; Choi, K.M. The implications of sarcopenia and sarcopenic obesity on cardiometabolic disease. J. Cell. Biochem. 2015, 116, 1171–1178. [Google Scholar] [CrossRef]
- Dhingra, R.; Vasan, R.S. Age as a risk factor. Med. Clin. N. Am. 2012, 96, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Keenan, C.R.; White, R.H. Age as a risk factor for venous thromboembolism after major surgery. Curr. Opin. Pulm. Med. 2005, 11, 398–402. [Google Scholar] [CrossRef]
- Koshiyama, M.; Tamaki, K.; Ohsawa, M. Age-specific incidence rates of atrial fibrillation and risk factors for the future development of atrial fibrillation in the Japanese general population. J. Cardiol. 2020, 77, 88–92. [Google Scholar] [CrossRef]
- Zhang, H.; Rogers, K.; Sukkar, L.; Jun, M.; Kang, A.; Young, T.; Campain, A.; Cass, A.; Chow, C.K.; Comino, E.; et al. Prevalence, incidence and risk factors of diabetes in Australian adults aged ≥ 45 years: A cohort study using linked routinely-collected data. J. Clin. Transl. Endocrinol. 2020, 22, 100240. [Google Scholar] [CrossRef] [PubMed]
- Mosher, C.L.; Weber, J.M.; Frankel, C.W.; Neely, M.L.; Palmer, S.M. Risk factors for mortality in lung transplant recipients aged ≥ 65 years: A retrospective cohort study of 5815 patients in the scientific registry of transplant recipients. J. Heart Lung Transplant. 2020. [Google Scholar] [CrossRef]
- Norose, K.; Clark, J.I.; Syed, N.A.; Basu, A.; Heber-Katz, E.; Sage, E.H.; Howe, C.C. SPARC deficiency leads to early-onset cataractogenesis. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2674–2680. [Google Scholar] [PubMed]
- Singh, M. Mood, food, and obesity. Front. Psychol. 2014, 5, 925. [Google Scholar] [CrossRef] [Green Version]
- Ortolani, D.; Oyama, L.M.; Ferrari, E.M.; Melo, L.L.; Spadari-Bratfisch, R.C. Effects of comfort food on food intake, anxiety-like behavior and the stress response in rats. Physiol. Behav. 2011, 103, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, A.D.; Graves, D.C.; Motamed, K.; Sage, E.H. SPARC-null mice exhibit increased adiposity without significant differences in overall body weight. Proc. Natl. Acad. Sci. USA 2003, 100, 6045–6050. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, A.J.; Abdelhafiz, A.H. Cardiometabolic disease in the older person: Prediction and prevention for the generalist physician. Cardiovasc. Endocrinol. Metab. 2020, 9, 90–95. [Google Scholar] [CrossRef]
- Islam, H.; Hood, D.A.; Gurd, B.J. Looking beyond PGC-1α: Emerging regulators of exercise-induced skeletal muscle mitochondrial biogenesis and their activation by dietary compounds. Appl. Physiol. Nutr. Metab. 2020, 45, 11–23. [Google Scholar] [CrossRef]
- Su, J.; Chen, X.; Kanekura, T. A CD147-targeting siRNA inhibits the proliferation, invasiveness, and VEGF production of human malignant melanoma cells by down-regulating glycolysis. Cancer Lett. 2009, 273, 140–147. [Google Scholar] [CrossRef]
- Gillies, R.J.; Raghunand, N.; Karczmar, G.S.; Bhujwalla, Z.M. MRI of the tumor microenvironment. J. Magn. Reason. Imaging 2002, 16, 430–450. [Google Scholar] [CrossRef]
- Gladden, L.B. Current trends in lactate metabolism: Introduction. Med. Sci. Sports Exerc. 2008, 40, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Lactate shuttles in nature. Biochem. Soc. Trans. 2002, 30, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Current concepts in lactate exchange. Med. Sci. Sports Exerc. 1991, 23, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Hohenester, E.; Göhring, W.; Timpl, R. Crystal structure and mapping by site-directed mutagenesis of the collagen-binding epitope of an activated form of BM-40/SPARC/osteonectin. EMBO J. 1998, 17, 1625–1634. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.J. The role of collagen in the development of muscle and its relationship to eating quality. J. Anim. Sci. 1985, 60, 1580–1587. [Google Scholar] [CrossRef]
- Au, E.; Richter, M.W.; Vincent, A.J.; Tetzlaff, W.; Aebersold, R.; Sage, E.H.; Roskams, A.J. SPARC from olfactory ensheathing cells stimulates Schwann cells to promote neurite outgrowth and enhances spinal cord repair. J. Neurosci. 2007, 27, 7208–7221. [Google Scholar] [CrossRef]
- Lee, M.H.; Amin, N.D.; Venkatesan, A.; Wang, T.; Tyagi, R.; Pant, H.C.; Nath, A. Impaired neurogenesis and neurite outgrowth in an HIV-gp120 transgenic model is reversed by exercise via BDNF production and Cdk5 regulation. J. Neurovirol. 2013, 19, 418–431. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.J.; Lau, P.W.; Roskams, A.J. SPARC is expressed by macroglia and microglia in the developing and mature nervous system. Dev. Dyn. 2008, 237, 1449–1462. [Google Scholar] [CrossRef]
- Brekken, R.A.; Puolakkainen, P.; Graves, D.C.; Workman, G.; Lubkin, S.R.; Sage, E.H. Enhanced growth of tumors in SPARC null mice is associated with changes in the ECM. J. Clin. Investig. 2003, 111, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Puolakkainen, P.A.; Brekken, R.A.; Muneer, S.; Sage, E.H. Enhanced growth of pancreatic tumors in SPARC-null mice is associated with decreased deposition of extracellular matrix and reduced tumor cell apoptosis. Mol. Cancer Res. 2004, 2, 215–224. [Google Scholar]
- Said, N.; Frierson, H.F.; Sanchez-Carbayo, M.; Brekken, R.A.; Theodorescu, D. Loss of SPARC in bladder cancer enhances carcinogenesis and progression. J. Clin. Investig. 2013, 123, 751–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.P.; Hsiao, M. Exercise-induced SPARC prevents tumorigenesis of colon cancer. Gut 2013, 62, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Aoi, W. Possibility of the novel myokine SPARC: A mechanistic approach to colon cancer prevention by physical exercise. Jpn. J. Phys. Fit. Sports Med. 2013, 62, 263–271. [Google Scholar] [CrossRef]
- Konhilas, J.P.; Maass, A.H.; Luckey, S.W.; Stauffer, B.L.; Olson, E.N.; Leinwand, L.A. Sex modifies exercise and cardiac adaptation in mice. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H2768–H2776. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, Y.; Wang, H.; Chen, W.; Liu, X. Effect of force level and gender on pinch force perception in healthy adults. Iperception 2020, 11, 2041669520927043. [Google Scholar] [CrossRef]
- Sundberg, S. Maximal oxygen uptake in relation to age in blind and normal boys and girls. Acta Paediatr. Scand. 1982, 71, 603–608. [Google Scholar] [CrossRef]
- Hafen, P.S.; Vehrs, P.R. Sex-related differences in the maximal lactate steady state. Sports 2018, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ji, L. Gender differences in peak blood lactate concentration and lactate removal. Ann. Sports Med. Res. 2016, 3, 1088. [Google Scholar]
- Green, H.J.; Fraser, I.G.; Ranney, D.A. Male and female differences in enzyme activities of energy metabolism in vastus lateralis muscle. J. Neurol. Sci. 1984, 65, 323–331. [Google Scholar] [CrossRef]
- Yoshioka, M.; St-Pierre, S.; Suzuki, M.; Tremblay, A. Effects of red pepper added to high-fat and high-carbohydrate meals on energy metabolism and substrate utilization in Japanese women. Br. J. Nutr. 1998, 80, 503–510. [Google Scholar] [CrossRef]
- Yoshioka, M.; Lim, K.; Kikuzato, S.; Kiyonaga, A.; Tanaka, H.; Shindo, M.; Suzuki, M. Effects of red-pepper diet on the energy metabolism in men. J. Nutr. Sci. Vitam. (Tokyo) 1995, 41, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Simoneau, J.A.; Bouchard, C. Human variation in skeletal muscle fiber-type proportion and enzyme activities. Am. J. Physiol. 1989, 257, E567–E572. [Google Scholar] [CrossRef] [PubMed]
- Simoneau, J.A.; Lortie, G.; Boulay, M.R.; Thibault, M.C.; Thériault, G.; Bouchard, C. Skeletal muscle histochemical and biochemical characteristics in sedentary male and female subjects. Can. J. Physiol. Pharm. 1985, 63, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Di Luigi, L.; Romanelli, F.; Sgrò, P.; Lenzi, A. Andrological aspects of physical exercise and sport medicine. Endocrine 2012, 42, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Isacco, L.; Duché, P.; Boisseau, N. Influence of hormonal status on substrate utilization at rest and during exercise in the female population. Sports Med. 2012, 42, 327–342. [Google Scholar] [CrossRef] [PubMed]
- Amelink, G.J.; Bär, P.R. Exercise-induced muscle protein leakage in the rat. Effects of hormonal manipulation. J. Neurol. Sci. 1986, 76, 61–68. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Young | Old | 2-Way ANOVA | ||||||
|---|---|---|---|---|---|---|---|---|
| Wild-type | Knockout | Wild-type | Knockout | A | G | A × G | ||
| During an incremental exercise test (wk 4) | ||||||||
| LT speed | m/min | 8.7± 0.4 | 7.8 ± 0.5 | 5.5 ± 0.6 | 5.4 ± 0.4 | Y > O | - | - |
| Lactate at rest | mM | 2.8 ± 0.2 | 2.6 ± 0.2 | 3.1 ± 0.4 | 3.8 ± 0.2 | O > Y | - | KO: O > Y |
| Lactate at LT | mM | 3.2 ± 0.2 | 3.1 ± 0.3 | 3.4 ± 0.4 | 3.3 ± 0.5 | - | - | - |
| During LT training (wk 1–12) | ||||||||
| Mean exercise speed | m/min | 7.5 ± 0.2 | 7.6 ± 0.2 | 5.5 ± 0.0 | 5.3 ± 0.0 | Y > O | - | - |
| Total exercise time | min | 3332 ± 52 | 3353 ± 45 | 3334 ± 1 | 3335 ± 3 | - | - | - |
| Total exercise distance | m | 24,903 ± 763 | 25,445 ± 670 | 18,219 ± 41 | 17,811 ± 156 | Y > O | - | - |
| During the last LT training (wk 12) | ||||||||
| Speed | m/min | 7.8 ± 0.2 | 7.9 ± 0.2 | 5.5 ± 0.0 | 5.3 ± 0.1 | Y > O | - | - |
| Lactate | mM | 2.7 ± 0.3 | 2.9 ± 0.3 | 2.1 ± 0.2 | 2.5 ± 0.2 | Y > O | - | - |
| Young | Old | ANOVA | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wild-type | Knockout | Wild-type | Knockout | |||||||||||||
| Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | A | G | Ex | A × G | A × Ex | G × Ex | A × G × Ex | ||
| At sacrifice (12 h fast) | 3-way | |||||||||||||||
| Blood lactate | mM | 1.09 ± 0.12 | 0.88 ± 0.08 | 1.28 ± 0.38 | 1.24 ± 0.15 | 0.89 ± 0.07 | 0.99 ± 0.13 | 0.98 ± 0.08 | 0.75 ± 0.04 | Y ˃ O * | - | - | - | - | - | - |
| Post glucose gavage (6 h fast) | 4-way | |||||||||||||||
| Blood lactate | AUC | Y > O | - | - | - | - | - | - | ||||||||
| Pre | 615 ± 35 | 498 ± 63 | 479 ± 29 | 533 ± 44 | 429 ± 29 | 424 ± 24 | 473 ± 39 | 461 ± 54 | ||||||||
| At wk 5 | 529 ± 32 | 509 ± 34 | 503 ± 41 | 519 ± 38 | 422 ± 27 | 450 ± 42 | 479 ± 44 | 392 ± 32 | ||||||||
| After 12 wks | 465 ± 34 | 391 ± 27 | 452 ± 22 | 486 ± 45 | 440 ± 56 | 381 ± 20 | 414 ± 35 | 371 ± 28 | ||||||||
| Young | Old | 3-Way ANOVA | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wild-type | Knockout | Wild-type | Knockout | |||||||||||||
| Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | A | G | Ex | A × G | A × Ex | G × Ex | A × G × Ex | ||
| Body weight | g | 29.5 ± 0.7 | 28.9 ± 1.0 | 27.8 ± 0.6 | 25.0 ± 0.8 | 37.4 ± 2.1 | 35.2 ± 1.0 | 31.3 ± 1.5 | 30.4 ± 0.7 | O ˃ Y | WT ˃ KO | Sed > Ex * | - | - | - | - |
| Tissues weights | ||||||||||||||||
| Brain | mg | 430 ± 2 | 430 ± 6 | 448 ± 4 | 445 ± 3 | 432 ± 7 | 436 ± 2 | 451 ± 4 | 446 ± 4 | - | KO ˃ WT | - | - | - | - | - |
| % | 1.47 ± 0.03 | 1.50 ± 0.05 | 1.62 ± 0.04 | 1.80 ± 0.05 | 1.18 ± 0.05 | 1.25 ± 0.03 | 1.48 ± 0.07 | 1.47 ± 0.02 | Y ˃ O | KO ˃ WT | Ex > Sed * | - | - | - | WT-Sed: Y ˃ O, WT-Ex: Y ˃ O, KO-Ex: Y >> O | |
| Pituitary gland | mg | 1.52 ± 0.20 | 1.49 ± 0.10 | 1.47 ± 0.10 | 1.33 ± 0.10 | 1.77 ± 0.16 | 1.76 ± 0.13 | 1.78 ± 0.04 | 1.65 ± 0.12 | O ˃ Y | - | - | - | - | - | - |
| % | 0.0052 ± 0.0007 | 0.0052 ± 0.0003 | 0.0053 ± 0.0004 | 0.0054 ± 0.0004 | 0.0049 ± 0.0006 | 0.0050 ± 0.0004 | 0.0059 ± 0.0003 | 0.0055 ± 0.0005 | - | - | - | - | - | - | - | |
| Hypothalamus | mg | 8.7 ± 0.7 | 9.1 ± 0.6 | 8.0 ± 0.5 | 8.9 ± 0.8 | 10.7 ± 0.9 | 10.8 ± 0.8 | 9.4 ± 0.8 | 11.0 ± 0.5 | O ˃ Y | - | - | - | - | - | - |
| % | 0.030 ± 0.003 | 0.032 ± 0.003 | 0.029 ± 0.002 | 0.036 ± 0.003 | 0.030 ± 0.003 | 0.031 ± 0.003 | 0.030 ± 0.003 | 0.036 ± 0.002 | - | - | Ex ˃ Sed * | - | - | - | - | |
| Liver | mg | 984 ± 27 | 954 ± 33 | 984 ± 35 | 899 ± 30 | 1370 ± 136 | 1114 ± 28 | 1174 ± 82 | 1101 ± 24 | O ˃ Y | - | Sed ˃ Ex | - | - | - | - |
| % | 3.34 ± 0.04 | 3.31 ± 0.09 | 3.54 ± 0.09 | 3.60 ± 0.04 | 3.59 ± 0.14 | 3.17 ± 0.04 | 3.71 ± 0.12 | 3.62 ± 0.05 | - | KO ˃ WT | Sed ˃ Ex * | - | Sed: O ˃ Y * | Ex: KO ˃ WT | - | |
| Heart | mg | 138 ± 3 | 149 ± 7 | 146 ± 6 | 131 ± 4 | 159 ± 6 | 135 ± 4 | 152 ± 5 | 159 ± 7 | O ˃ Y | - | - | KO: O ˃ Y | - | - | WT-Sed: O ˃ Y, WT-Ex: Y ˃ O *, KO-Ex: O ˃ Y |
| % | 0.47 ± 0.01 | 0.51 ± 0.01 | 0.53 ± 0.02 | 0.53 ± 0.01 | 0.43 ± 0.01 | 0.38 ± 0.01 | 0.50 ± 0.02 | 0.52 ± 0.02 | Y ˃ O | KO ˃ WT | - | WT: Y >> O | - | - | WT-Sed: Y ˃ O, WT-Ex: Y >> O | |
| Aorta | mg | 12.8 ± 1.5 | 12.0 ± 1.5 | 12.9 ± 1.3 | 9.0 ± 0.6 | 13.0 ± 1.5 | 12.3 ± 1.0 | 9.7 ± 1.0 | 9.9 ± 0.7 | - | WT ˃ KO | - | - | - | - | - |
| % | 0.044 ± 0.006 | 0.042 ± 0.006 | 0.047 ± 0.005 | 0.037 ± 0.003 | 0.034 ± 0.003 | 0.035 ± 0.003 | 0.031 ± 0.003 | 0.032 ± 0.002 | Y ˃ O | - | - | - | - | - | - | |
| White adipose tissue ** | mg | 1021 ± 206 | 812 ± 126 | 955 ± 138 | 684 ± 125 | 2913 ± 362 | 2649 ± 175 | 1804 ± 226 | 1505 ± 164 | O ˃ Y | WT ˃ KO | Sed > Ex * | WT: O >> Y, KO: O>Y | - | - | - |
| % | 3.36 ± 0.58 | 2.72 ± 0.36 | 3.36 ± 0.44 | 2.63 ± 0.38 | 7.52 ± 0.65 | 7.44 ± 0.32 | 5.51 ± 0.50 | 4.85 ± 0.43 | O ˃ Y | WT ˃ KO | - | WT: O >> Y, KO: O > Y | - | - | - | |
| Skeletal muscle *** | mg | 532 ± 11 | 524 ± 15 | 432 ± 8 | 413 ± 7 | 508 ± 11 | 510 ± 6 | 416 ± 8 | 417 ± 7 | Y ˃ O * | WT ˃ KO | - | - | - | - | |
| % | 1.81 ± 0.04 | 1.82 ± 0.05 | 1.56 ± 0.04 | 1.67 ± 0.05 | 1.39 ± 0.05 | 1.46 ± 0.05 | 1.36 ± 0.06 | 1.38 ± 0.03 | Y ˃ O | WT ˃ KO | - | WT: Y >> O, KO: Y ˃ O | - | - | - | |
| Tibialis anterior muscle | mg | 133 ± 3 | 138 ± 4 | 117 ± 3 | 114 ± 2 | 133 ± 6 | 127 ± 3 | 110 ± 4 | 118 ± 3 | Y ˃ O * | WT ˃ KO | - | - | - | - | WT-Ex: Y ˃ O |
| % | 0.45 ± 0.01 | 0.48 ± 0.01 | 0.42 ± 0.01 | 0.46 ± 0.01 | 0.36 ± 0.02 | 0.36 ± 0.01 | 0.36 ± 0.01 | 0.39 ± 0.02 | Y ˃ O | - | Ex > Sed | WT: Y >> O, KO: Y > O | - | - | - | |
| Young | Old | 3-Way ANOVA | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wild-type | Knockout | Wild-type | Knockout | |||||||||||||
| Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | Sedentary | Exercise | A | G | Ex | A × G | A × Ex | G × Ex | A × G × Ex | ||
| Grip power | ||||||||||||||||
| Mean | g | 240 ± 12 | 248 ± 11 | 226 ± 10 | 208 ± 11 | 216 ± 8 | 220 ± 8 | 182 ± 8 | 203 ± 9 | Y ˃ O | WT ˃ KO | - | - | - | - | - |
| g/BW | 8.2 ± 0.5 | 8.6 ± 0.4 | 8.1 ± 0.4 | 8.4 ± 0.4 | 5.8 ± 0.4 | 6.1 ± 0.2 | 5.9 ± 0.5 | 6.6 ± 0.3 | Y ˃ O | - | - | - | - | - | ||
| Max | g | 308 ± 12 | 301 ± 11 | 277 ± 11 | 250 ± 9 | 250 ± 8 | 257 ± 9 | 210 ± 8 | 235 ± 10 | Y ˃ O | WT ˃ KO | - | - | - | - | - |
| g/BW | 10.5 ± 0.4 | 10.5 ± 0.4 | 10.0 ± 0.4 | 10.1 ± 0.4 | 6.7 ± 0.5 | 7.2 ± 0.3 | 6.8 ± 0.5 | 7.6 ± 0.3 | Y ˃ O | - | Ex > Sed * | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghanemi, A.; Melouane, A.; Yoshioka, M.; St-Amand, J. Exercise Training of Secreted Protein Acidic and Rich in Cysteine (Sparc) KO Mice Suggests That Exercise-Induced Muscle Phenotype Changes Are SPARC-Dependent. Appl. Sci. 2020, 10, 9108. https://doi.org/10.3390/app10249108
Ghanemi A, Melouane A, Yoshioka M, St-Amand J. Exercise Training of Secreted Protein Acidic and Rich in Cysteine (Sparc) KO Mice Suggests That Exercise-Induced Muscle Phenotype Changes Are SPARC-Dependent. Applied Sciences. 2020; 10(24):9108. https://doi.org/10.3390/app10249108
Chicago/Turabian StyleGhanemi, Abdelaziz, Aicha Melouane, Mayumi Yoshioka, and Jonny St-Amand. 2020. "Exercise Training of Secreted Protein Acidic and Rich in Cysteine (Sparc) KO Mice Suggests That Exercise-Induced Muscle Phenotype Changes Are SPARC-Dependent" Applied Sciences 10, no. 24: 9108. https://doi.org/10.3390/app10249108
APA StyleGhanemi, A., Melouane, A., Yoshioka, M., & St-Amand, J. (2020). Exercise Training of Secreted Protein Acidic and Rich in Cysteine (Sparc) KO Mice Suggests That Exercise-Induced Muscle Phenotype Changes Are SPARC-Dependent. Applied Sciences, 10(24), 9108. https://doi.org/10.3390/app10249108