
Physical Activity and Redox Balance in the Elderly: Signal Transduction Mechanisms

 , , , , ,
, , , , , 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Physical Activity and Redox Balance in the Young and Adults
3. Physical Activity and Redox Balance in the Adult and the Elderly
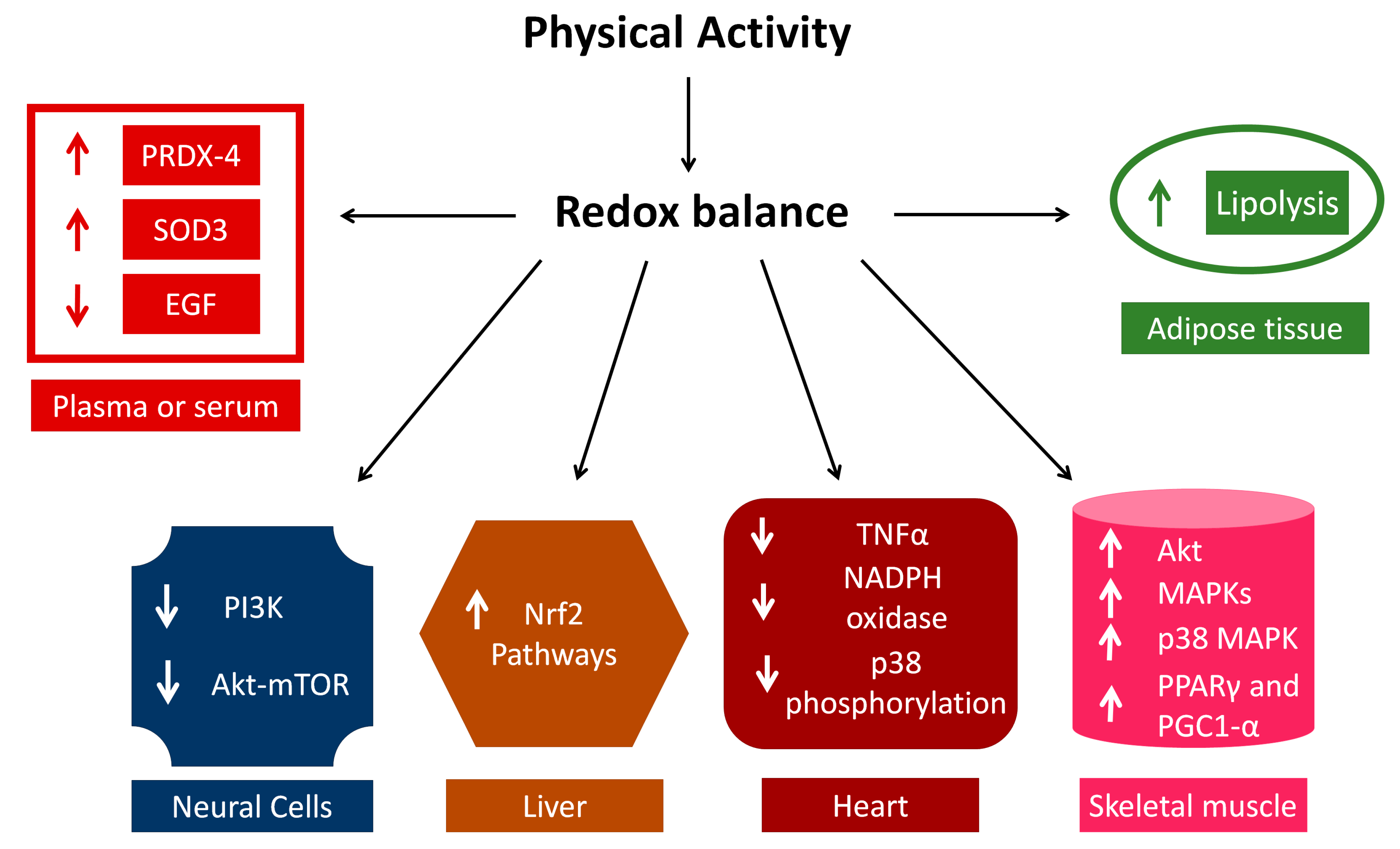
4. Insights into Signal Transduction
4.1. Plasma and Adipose Tissue
4.2. Nervous System
4.3. Liver
4.4. Skeletal and Cardiac Muscle
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SOD3 | Superoxide Dismutase 3 |
| PRDX-4 | Peroxiredoxin-4 |
| EGF | Epidermal Growth Factor |
| PI3Kinase | Phosphatidyl Inositol 3 Kinase |
| Akt | Protein Kinase B |
| mTOR | mammalian Target Of Rapamycin |
| Nrf2 | Nuclear factor erythroid-2-related factor 2 |
| TNF-alpha | Tumour Necrosis Factor alpha |
| NADPH | Nicotinamide Adenine Dinucleotide Phosphate |
| MAPKinase | Mitogen-Activated Protein Kinase |
| PPAR-gamma | Peroxisome Proliferator-Activated Receptor gamma |
| PGC-1-alpha | Peroxisome proliferator-activated receptor Gamma Coactivator 1 alpha |
References
- World Health Organization (WHO). Available online: https://www.who.int/news-room/q-a-detail/ageing-healthy-ageing-and-functional-ability (accessed on 30 October 2020).
- Ji, L.L.; Leeuwenburgh, C.; Leichtweis, S.; Gore, M.; Fiebig, R.; Hollander, J.; Bejma, J. Oxidative stress and aging. Role of exercise and its influences on antioxidant systems. Ann. N. Y. Acad. Sci. 1998, 854, 102–117. [Google Scholar] [CrossRef]
- Viña, J.; Olaso-Gonzalez, G.; Arc-Chagnaud, C.; De la Rosa, A.; Gomez-Cabrera, M.C. Modulating Oxidant Levels to Promote Healthy Aging. Antioxid. Redox Signal. 2020. [Google Scholar] [CrossRef]
- Ji, L.L. Antioxidants and oxidative stress in exercise. Proc. Soc. Exp. Biol. Med. 1999, 222, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Massafra, C.; Gioia, D.; De Felice, C.; Picciolini, E.; De Leo, V.; Bonifazi, M.; Bernabei, A. Effects of estrogens and androgens on erythrocyte antioxidant superoxide dismutase, catalase and glutathione peroxidase activities during the menstrual cycle. J. Endocrinol. 2000, 167, 447–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Kwak, Y.S. Impact of aerobic and anaerobic exercise training on oxidative stress and antioxidant defense in athletes. J. Exerc. Rehabil. 2016, 12, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, H.E.; Weimann, A.; Loft, S. Methods to detect DNA damage by free radicals: Relation to exercise. Proc. Nutr. Soc. 1999, 58, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Debevec, T.; Millet, G.P.; Pialoux, V. Hypoxia-Induced Oxidative Stress Modulation with Physical Activity. Front. Physiol. 2017, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Tofas, T.; Draganidis, D.; Deli, C.K.; Georgakouli, K.; Fatouros, I.G.; Jamurtas, A.Z. Exercise-Induced Regulation of Redox Status in Cardiovascular Diseases: The Role of Exercise Training and Detraining. Antioxidants 2019, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- de Sousa, C.V.; Sales, M.M.; Rosa, T.S.; Lewis, J.E.; de Andrade, R.V.; Simões, H.G. The Antioxidant Effect of Exercise: A Systematic Review and Meta-Analysis. Sports Med. 2017, 47, 277–293. [Google Scholar] [CrossRef]
- Nocella, C.; Cammisotto, V.; Pigozzi, F.; Borrione, P.; Fossati, C.; D’Amico, A.; Cangemi, R.; Peruzzi, M.; Gobbi, G.; Ettorre, E.; et al. Impairment between Oxidant and Antioxidant Systems: Short- and Long-term Implications for Athletes’ Health. Nutrients 2019, 11, 1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, R.F.; de Moraes, S.R.A.; Augusto, R.L.; de Freitas Zanona, A.; Matos, D.; Aidar, F.J.; da Silveira Andrade-da-Costa, B.L. Endurance training on rodent brain antioxidant capacity: A meta-analysis. Neurosci. Res. 2019, 145, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pinho, R.A.; Aguiar, A.S., Jr.; Radák, Z. Effects of Resistance Exercise on Cerebral Redox Regulation and Cognition: An Interplay between Muscle and Brain. Antioxidants 2019, 8, 529. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Rossman, M.J.; Gifford, J.R.; Bharath, L.P.; Bauersachs, J.; Richardson, R.S.; Abel, E.D.; Symons, J.D.; Riehle, C. Exercise training improves vascular mitochondrial function. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H821–H829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, H.T.; So, W.Y. The effects of aerobic exercise training on oxidant-antioxidant balance, neurotrophic factor levels, and blood-brain barrier function in obese and non-obese men. J. Sport Health Sci. 2017, 6, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Di Liegro, C.M.; Schiera, G.; Proia, P.; Di Liegro, I. Physical Activity and Brain Health. Genes 2019, 10, 720. [Google Scholar] [CrossRef] [Green Version]
- Arad, A.D.; Basile, A.J.; Albu, J.; DiMenna, F.J. No Influence of Overweight/Obesity on Exercise Lipid Oxidation. Int. J. Mol. Sci. 2020, 21, 1614. [Google Scholar] [CrossRef] [Green Version]
- Sadowska-Krępa, E.; Kłapcińska, B.; Pokora, I.; Domaszewski, P.; Kempa, K.; Podgórski, T. Effects of Six-Week Ginkgo biloba Supplementation on Aerobic Performance, Blood Pro/Antioxidant Balance, and Serum Brain-Derived Neurotrophic Factor in Physically Active Men. Nutrients 2017, 9, 803. [Google Scholar] [CrossRef] [Green Version]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Cavarretta, E.; Peruzzi, M.; Del Vescovo, R.; Di Pilla, F.; Gobbi, G.; Serdoz, A.; Ferrara, R.; Schirone, L.; Sciarretta, S.; Nocella, C.; et al. Dark Chocolate Intake Positively Modulates Redox Status and Markers of Muscular Damage in Elite Football Athletes: A Randomized Controlled Study. Oxidative Med. Cell Longev. 2018, 2018, 4061901. [Google Scholar] [CrossRef]
- Georgakouli, K.; Fatouros, I.G.; Fragkos, A.; Tzatzakis, T.; Deli, C.K.; Papanikolaou, K.; Koutedakis, Y.; Jamurtas, A.Z. Exercise and Redox Status Responses Following Alpha-Lipoic Acid Supplementation in G6PD Deficient Individuals. Antioxidants 2018, 7, 162. [Google Scholar] [CrossRef] [Green Version]
- Kabasakalis, A.; Kyparos, A.; Tsalis, G.; Loupos, D.; Pavlidou, A.; Kouretas, D.; Kabasakalis, A.; Tsalis, G.; Zafrana, E.; Loupos, D.; et al. Effects of endurance and high-intensity swimming exercise on the redox status of adolescent male and female swimmers. J. Sports Sci. 2014, 32, 747–756. [Google Scholar] [CrossRef]
- Kabasakalis, A.; Kyparos, A.; Tsalis, G.; Loupos, D.; Pavlidou, A.; Kouretas, D. Blood oxidative stress markers after ultramarathon swimming. J. Strength Cond. Res. 2011, 25, 805–811. [Google Scholar] [CrossRef]
- Bellafiore, M.; Bianco, A.; Battaglia, G.; Naccari, M.S.; Caramazza, G.; Padulo, J.; Chamari, K.; Paoli, A.; Palma, A. Training session intensity affects plasma redox status in amateur rhythmic gymnasts. J. Sport Health Sci. 2019, 8, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, C.; Musci, R.; Whedbee, M. Vitamin supplementation and resistance exercise-induced muscle hypertrophy: Shifting the redox balance scale? J. Physiol. 2015, 593, 2991–2992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, G.; Hamarsland, H.; Cumming, K.T.; Johansen, R.E.; Hulmi, J.J.; Borsheim, E.; Wiig, H.; Garthe, I.; Raastad, T. Vitamin C and E supplementation alters protein signalling after a strength training session, but not muscle growth during 10 weeks of training. J. Physiol. 2014, 592, 5391–5408. [Google Scholar] [CrossRef]
- Bruns, D.R.; Ehrlicher, S.E.; Khademi, S.; Biela, L.M.; Peelor, F.F.; Miller, B.F.; Hamilton, K.L. Differential effects of vitamin C or protandim on skeletal muscle adaptation to exercise. J. Appl. Physiol. 2018, 125, 661–671. [Google Scholar] [CrossRef]
- Traustadóttir, T.; Davies, S.S.; Su, Y.; Choi, L.; Brown-Borg, H.M.; Roberts, L.J., 2nd; Harman, S.M. Oxidative stress in older adults: Effects of physical fitness. Age (Dordr.) 2012, 34, 969–982. [Google Scholar] [CrossRef] [Green Version]
- Aldred, S.; Rohalu, M. A moderate intensity exercise program did not increase the oxidative stress in older adults. Arch. Gerontol. Geriatr. 2011, 53, 350–353. [Google Scholar] [CrossRef] [PubMed]
- Done, A.J.; Traustadóttir, T. Aerobic exercise increases resistance to oxidative stress in sedentary older middle-aged adults. A pilot study. Age (Dordr.) 2016, 38, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Landers-Ramos, R.Q.; Corrigan, K.J.; Guth, L.M.; Altom, C.N.; Spangenburg, E.E.; Prior, S.J.; Hagberg, J.M. Short-term exercise training improves flow-mediated dilation and circulating angiogenic cell number in older sedentary adults. Appl. Physiol. Nutr. Metab. 2016, 41, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Pierno, S.; Tricarico, D.; Liantonio, A.; Mele, A.; Digennaro, C.; Rolland, J.-F.; Bianco, G.; Villanova, L.; Merendino, A.; Camerino, G.M.; et al. An olive oil derived antioxidant mixture ameliorates the age-related decline of skeletal muscle function. Age 2014, 36, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, M.H.; Allam, M.M. Resveratrol and/or exercise training counteract aging-associated decline of physical endurance in aged mice; targeting mitochondrial biogenesis and function. J. Physiol. Sci. 2018, 68, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Gliemann, L.; Nyberg, M.; Hellsten, Y. Effects of exercise training and resveratrol on vascular health in aging. Free Radic. Biol. Med. 2016, 98, 165–176. [Google Scholar] [CrossRef]
- Najjar, F.; Rizk, F.; Carnac, G. Protective effect of Rhus coriaria fruit extracts against hydrogen peroxide-induced oxidative stress in muscle progenitors and zebrafish embryos. PeerJ 2017, 5, e4144. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.C.; Crecelius, A.R.; Larson, D.G.; Dinenno, F.A. Acute ascorbic acid ingestion increases skeletal muscle blood flow and oxygen consumption via local vasodilation during graded handgrip exercise in older adults. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H360–H368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fougere, B.; van Kan, G.A.; Vellas, B.; Cesari, M. Redox Systems, Antioxidants and Sarcopenia. Curr. Protein Pept. Sci. 2018, 19, 643–648. [Google Scholar] [CrossRef]
- Angulo, J.; El Assar, M.; Álvarez-Bustos, A.; Rodríguez-Mañas, L. Physical activity and exercise: Strategies to manage frailty. Redox Biol. 2020, 35, 101513. [Google Scholar] [CrossRef]
- Szentesi, P.; Csernoch, L.; Dux, L.; Keller-Pintér, A. Changes in Redox Signaling in the Skeletal Muscle with Aging. Oxid. Med. Cell Longev. 2019, 2019, 4617801. [Google Scholar] [CrossRef]
- Verdijk, L.B.; Snijders, T.; Drost, M.; Delhaas, T.; Kadi, F.; van Loon, L.J.C. Satellite cells in human skeletal muscle; from birth to old age. Age 2014, 36, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Blocquiaux, S.; Gorski, T.; Van Roie, E.; Ramaekers, M.; Van Thienen, R.; Nielens, H.; Delecluse, C.; De Bock, K.; Thomis, M. The effect of resistance training, detraining and retraining on muscle strength and power, myofibre size, satellite cells and myonuclei in older men [published correction appears in Exp Gerontol. Exp. Gerontol. 2020, 133, 110860. [Google Scholar] [CrossRef]
- Fry, C.S.; Noehren, B.; Mula, J.; Ubele, M.F.; Westgate, P.M.; Kern, P.A.; Peterson, C.A. Fibre type-specific satellite cell response to aerobic training in sedentary adults. J. Physiol. 2014, 592, 2625–2635. [Google Scholar] [CrossRef]
- Pittaluga, M.; Sgadari, A.; Dimauro, I.; Tavazzi, B.; Parisi, P.; Caporossi, D. Physical exercise and redox balance in type 2 diabetics: Effects of moderate training on biomarkers of oxidative stress and DNA damage evaluated through comet assay. Oxid. Med. Cell Longev. 2015, 981242. [Google Scholar] [CrossRef]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative stress in neurodegeneration. Adv. Pharmacol. Sci. 2011, 2011, 572634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinet, C.; Gonzalo, H.; Fleitas, C.; Menal, M.J.; Egea, J. Oxidative stress and neurodegenerative diseases: A neurotrophic approach. Curr. Drug Targets. 2015, 16, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.P.; Tran, N.; Geekiyanage, H.; Liu, L.; Chan, C. Curcumin-induced upregulation of the anti-tau cochaperone BAG2 in primary rat cortical neurons. Neurosci. Lett. 2013, 554, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.X.; Liang, J.H.; Xu, Y.; Wang, Y.Q. Effects of physical activity and exercise on the cognitive function of patients with Alzheimer disease: A meta-analysis. BMC Geriatr. 2019, 19, 181. [Google Scholar] [CrossRef]
- Accattato, F.; Greco, M.; Pullano, S.A.; Carè, I.; Fiorillo, A.S.; Pujia, A.; Montalcini, T.; Foti, D.P.; Brunetti, A.; Gulletta, E. Effects of acute physical exercise on oxidative stress and inflammatory status in young, sedentary obese subjects. PLoS ONE 2017, 12, e0178900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatzinikolaou, A.; Fatouros, I.; Petridou, A.; Jamurtas, A.; Avloniti, A.; Douroudos, I.; Mastorakos, G.; Lazaropoulou, C.; Papassotiriou, I.; Tournis, S.; et al. Adipose tissue lipolysis is upregulated in lean and obese men during acute resistance exercise. Diabetes Care. 2008, 31, 1397–1399. [Google Scholar] [CrossRef] [Green Version]
- Karner-Rezek, K.; Knechtle, B.; Fenzl, M.; Gredig, J.; Rosemann, T. Does continuous endurance exercise in water elicit a higher release of ANP and BNP and a higher plasma concentration of FFAs in pre-obese and obese men than high intensity intermittent endurance exercise?‹study protocol for a randomized controlled trial. Trials 2013, 14, 328. [Google Scholar] [CrossRef] [Green Version]
- Mrakic-Sposta, S.; Gussoni, M.; Vezzoli, A.; Dellanoce, C.; Comassi, M.; Giardini, G.; Bruno, R.M.; Montorsi, M.; Corciu, A.; Greco, F.; et al. Acute Effects of Triathlon Race on Oxidative Stress Biomarkers. Oxid. Med. Cell Longev. 2020, 2020, 3062807. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, D.B.; Willis, L.H.; Slentz, C.A.; Hoselton, A.; Kelly, L.; Huebner, J.L.; Kraus, V.B.; Moss, J.; Muehlbauer, M.J.; Spielmann, G.; et al. Ten weeks of high-intensity interval walk training is associated with reduced disease activity and improved innate immune function in older adults with rheumatoid arthritis: A pilot study. Arthritis Res. Ther. 2018, 20, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Kader, S.M.; Al-Jiffri, O.H. Impact of aerobic versus resisted exercise training on systemic inflammation biomarkers and quality of Life among obese post-menopausal women. Afr. Health Sci. 2019, 19, 2881–2891. [Google Scholar] [CrossRef] [PubMed]
- Fatouros, I.; Chatzinikolaou, A.; Paltoglou, G.; Petridou, A.; Avloniti, A.; Jamurtas, A.; Goussetis, E.; Mitrakou, A.; Mougios, V.; Lazaropoulou, C.; et al. Acute resistance exercise results in catecholaminergic rather than hypothalamic-pituitary-adrenal axis stimulation during exercise in young men. Stress 2010, 13, 461–468. [Google Scholar] [CrossRef]
- Hirose, L.; Nosaka, K.; Newton, M.; Laveder, A.; Kano, M.; Peake, J.; Suzuki, K. Changes in inflammatory mediators following eccentric exercise of the elbow flexors. Exerc. Immunol. Rev. 2004, 10, 75–90. [Google Scholar] [PubMed]
- Ost, M.; Coleman, V.; Kasch, J.; Klaus, S. Regulation of myokine expression: Role of exercise and cellular stress. Free Radic. Biol. Med. 2016, 98, 78–89. [Google Scholar] [CrossRef]
- Radak, Z.; Suzuki, K.; Higuchi, M.; Balogh, L.; Boldogh, I.; Koltai, E. Physical exercise, reactive oxygen species and neuroprotection. Free Radic. Biol. Med. 2016, 98, 187–196. [Google Scholar] [CrossRef]
- Vilela, T.C.; de Andrade, V.M.; Radak, Z.; de Pinho, R.A. The role of exercise in brain DNA damage. Neural Regen Res. 2020, 15, 1981–1985. [Google Scholar] [CrossRef]
- Hsu, C.L.; Best, J.R.; Davis, J.C.; Nagamatsu, L.S.; Wang, S.; Boyd, L.A.; Hsiung, G.R.; Voss, M.W.; Eng, J.J.; Liu-Ambrose, T. Aerobic exercise promotes executive functions and impacts functional neural activity among older adults with vascular cognitive impairment. Br. J. Sports Med. 2018, 52, 184–191. [Google Scholar] [CrossRef]
- Feter, N.; Spanevello, R.M.; Soares, M.S.P.; Spohr, L.; Pedra, N.S.; Bona, N.P.; Freitas, M.P.; Gonzales, N.G.; Ito, L.G.M.S.; Stefanello, F.M.; et al. How does physical activity and different models of exercise training affect oxidative parameters and memory? Physiol. Behav. 2019, 201, 42–52. [Google Scholar] [CrossRef]
- Kou, X.; Chen, D.; Chen, N. Physical Activity Alleviates Cognitive Dysfunction of Alzheimer’s Disease through Regulating the mTOR Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 1591. [Google Scholar] [CrossRef] [Green Version]
- Done, A.J.; Newell, M.J.; Traustadóttir, T. Effect of exercise intensity on Nrf2 signalling in young men. Free Radic. Res. 2017, 51, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Mendoza, N.; Morales-González, Á.; Madrigal-Santillán, E.O.; Madrigal-Bujaidar, E.; Álvarez-González, I.; García-Melo, L.F.; Anguiano-Robledo, L.; Fregoso-Aguilar, T.; Morales-Gonzalez, J.A. Antioxidant and Adaptative Response Mediated by Nrf2 during Physical Exercise. Antioxidants 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Protective Effects of Sulforaphane on Exercise-Induced Organ Damage via Inducing Antioxidant Defense Responses. Antioxidants 2020, 9, 136. [Google Scholar] [CrossRef] [Green Version]
- Merry, T.L.; MacRae, C.; Pham, T.; Hedges, C.P.; Ristow, M. Deficiency in ROS-sensing nuclear factor erythroid 2-like 2 causes altered glucose and lipid homeostasis following exercise training. Am. J. Physiol. Cell Physiol. 2020, 318, C337–C345. [Google Scholar] [CrossRef]
- Huang, D.-D.; Fan, S.-D.; Chen, X.-Y.; Yan, X.-L.; Zhang, X.-Z.; Ma, B.-W.; Yu, D.-Y.; Xiao, W.-Y.; Zhuang, C.-L.; Yu, Z. Nrf2 deficiency exacerbates frailty and sarcopenia by impairing skeletal muscle mitochondrial biogenesis and dynamics in an age-dependent manner. Exp. Gerontol. 2019, 119, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.Y.; Chen, J.L.; Xiao, M.H. The effect of exercise, resveratrol or their combination on sarcopenia in aged rats via regulation of AMPK/Sirt 1 pathway. Exp. Gerontol. 2017, 98, 177–183. [Google Scholar]
- Abou-Samra, M.; Selvais, C.M.; Boursereau, R.; Lecompte, S.; Noel, L.; Brichard, S.M. AdipoRon, a new therapeutic prospect for Duchenne muscular dystrophy. J. Cachexia Sarcopenia Muscle 2020, 11, 518–533. [Google Scholar] [CrossRef]
- Kim, E.N.; Lim, J.H.; Kim, M.Y.; Ban, T.H.; Jang, I.A.; Yoon, H.E.; Park, C.W.; Chang, Y.S.; Choi, B.S. Resveratrol, an Nrf2 activator, ameliorates aging-related progressive renal injury. Aging (Albany N. Y.) 2018, 10, 83–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Luo, X.; Ma, Z.; Zhang, B.; Li, S.; Duan, X.; Yang, B.; Zhang, J. Swimming Exercise Protects against Insulin Resistance via Regulating Oxidative Stress through Nox4 and AKT Signaling in High-Fat Diet-Fed Mice. J. Diabetes Res. 2020, 2020, 2521590. [Google Scholar] [CrossRef] [Green Version]
- Lukjanenko, L.; Karaz, S.; Stuelsatz, P.; Gurriaran-Rodriguez, U.; Michaud, J.; Dammone, G.; Sizzano, F.; Mashinchian, O.; Ancel, S.; Migliavacca, E.; et al. Aging Disrupts Muscle Stem Cell Function by Impairing Matricellular WISP1 Secretion from Fibro-Adipogenic Progenitors. Cell Stem Cell 2019, 24, 433–446.e7. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, E.; Giammarioli, A.M.; Chiandotto, S.; Spoletini, I.; Rosano, G. Exercise-induced skeletal muscle remodeling and metabolic adaptation: Redox signaling and role of autophagy. Antioxid. Redox Signal. 2014, 21, 154–176. [Google Scholar] [CrossRef]
- Halling, J.F.; Jessen, H.; Nøhr-Meldgaard, J.; Buch, B.T.; Christensen, N.M.; Gudiksen, A.; Ringholm, S.; Neufer, P.D.; Prats, C.; Pilegaard, H. PGC-1α regulates mitochondrial properties beyond biogenesis with aging and exercise training. Am. J. Physiol Endocrinol. Metab. 2019, 317, E513–E525. [Google Scholar] [CrossRef]
- Cunha, T.F.; Bechara, L.R.; Bacurau, A.V.; Jannig, P.R.; Voltarelli, V.A.; Dourado, P.M.; Vasconcelos, A.R.; Scavone, C.; Ferreira, J.C.; Brum, P.C. Exercise training decreases NADPH oxidase activity and restores skeletal muscle mass in heart failure rats. J. Appl. Physiol. 2017, 122, 817–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roof, S.R.; Ho, H.-T.; Little, S.C.; Ostler, J.E.; Brundage, E.A.; Periasamy, M.; Villamena, F.A.; Györke, S.; Biesiadecki, B.J.; Heymes, C.; et al. Obligatory role of neuronal nitric oxide synthase in the heart’s antioxidant adaptation with exercise. J. Mol. Cell Cardiol. 2015, 81, 54–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Z.; Pan, S.S.; Shen, Y.J.; Ge, J. Exercise preconditioning-induced early and late phase of cardioprotection is associated with protein kinase C epsilon translocation. Circ. J. 2014, 78, 1636–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Ruíz, J.L.; Macías-López, A.; Alcalá-Vargas, F.; Guevara-Chávez, J.G.; Mejía-Uribe, A.; Silva-Palacios, A.; Zúñiga-Muñoz, A.; Zazueta, C.; Buelna-Chontal, M. Redox signaling in ischemic postconditioning protection involves PKCε and Erk1/2 pathways and converges indirectly in Nrf2 activation. Cell Signal. 2019, 64, 109417. [Google Scholar] [CrossRef]
- Buelna-Chontal, M.; Guevara-Chávez, J.G.; Silva-Palacios, A.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Zazueta, C. Nrf2-regulated antioxidant response is activated by protein kinase C in postconditioned rat hearts. Free Radic. Biol. Med. 2014, 74, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Carubbi, C.; Masselli, E.; Nouvenne, A.; Russo, D.; Galli, D.; Mirandola, P.; Gobbi, G.; Vitale, M. Laboratory diagnostics of inherited platelet disorders. Clin. Chem. Lab. Med. 2014, 52, 1091–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Marcantonio, D.; Galli, D.; Carubbi, C.; Gobbi, G.; Queirolo, V.; Martini, S.; Merighi, S.; Vaccarezza, M.; Maffulli, N.; Sykes, S.M.; et al. PKCε as a novel promoter of skeletal muscle differentiation and regeneration. Exp. Cell Res. 2015, 339, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Galli, D.; Carubbi, C.; Masselli, E.; Corradi, D.; Dei Cas, A.; Nouvenne, A.; Bucci, G.; Arcari, M.L.; Mirandola, P.; Vitale, M.; et al. PKCε is a negative regulator of PVAT-derived vessel formation. Exp. Cell Res. 2015, 330, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Masselli, E.; Carubbi, C.; Gobbi, G.; Mirandola, P.; Galli, D.; Martini, S.; Bonomini, S.; Crugnola, M.; Craviotto, L.; Aversa, F.; et al. Protein kinase Cɛ inhibition restores megakaryocytic differentiation of hematopoietic progenitors from primary myelofibrosis patients. Leukemia 2015, 29, 2192–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassini, A.; Zauli, G.; Migliaccio, G.; Migliaccio, A.R.; Pascuccio, M.; Pierpaoli, S.; Guidotti, L.; Capitani, S.; Vitale, M. Lineage-restricted expression of protein kinase C isoforms in hematopoiesis. Blood 1999, 93, 1178–1188. [Google Scholar] [CrossRef]
- Martini, S.; Soliman, T.; Gobbi, G.; Mirandola, P.; Carubbi, C.; Masselli, E.; Pozzi, G.; Parker, P.J.; Vitale, M. PKCε Controls Mitotic Progression by Regulating Centrosome Migration and Mitotic Spindle Assembly. Mol. Cancer Res. 2018, 16, 3–15. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, A.E.; Lennartz, M.R. Protein Kinase C-epsilon in Membrane Delivery during Phagocytosis. J. Immunol. Sci. 2018, 2, 26–32. [Google Scholar] [CrossRef]
- Basu, A. Regulation of Autophagy by Protein Kinase C-ε in Breast Cancer Cells. Int. J. Mol. Sci. 2020, 21, 4247. [Google Scholar] [CrossRef]
- Escobar, K.A.; Cole, N.H.; Mermier, C.M.; VanDusseldorp, T.A. Autophagy and aging: Maintaining the proteome through exercise and caloric restriction. Aging Cell 2019, 18, e12876. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.T.; Ang, S.J.; Tsai, S.Y. Sarcopenia: Tilting the Balance of Protein Homeostasis. Proteomics 2020, 20, e1800411. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zima, A.V.; Blatter, L.A. Redox regulation of cardiac calcium channels and transporters. Cardiovasc. Res. 2006, 71, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Cabassi, A.; Miragoli, M. Altered Mitochondrial Metabolism and Mechanosensation in the Failing Heart: Focus on Intracellular Calcium Signaling. Int. J. Mol. Sci. 2017, 18, 1487. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G. Overview on Peroxiredoxin. Mol. Cells. 2016, 39, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive oxygen species, aging and articular cartilage homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.J.; Knutson, S.T.; Soito, L.; Klomsiri, C.; Poole, L.B.; Fetrow, J.S. Analysis of the peroxiredoxin family: Using active-site structure and sequence information for global classification and residue analysis. Proteins 2011, 79, 947–964. [Google Scholar] [CrossRef] [Green Version]
- Milev, N.B.; Rhee, S.G.; Reddy, A.B. Cellular Timekeeping: It’s Redox o’Clock. Cold Spring Harb. Perspect Biol. 2018, 10, a027698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.G.; Wang, L.; Kaifu, T.; Li, J.; Li, X.; Li, L. Featured Article: Accelerated decline of physical strength in peroxiredoxin-3 knockout mice. Exp. Biol. Med. (Maywood) 2016, 241, 1395–1400. [Google Scholar] [CrossRef] [Green Version]
- Wadley, A.J.; Keane, G.; Cullen, T.; James, L.; Vautrinot, J.; Davies, M.; Hussey, B.; Hunter, D.J.; Mastana, S.; Holliday, A.; et al. Characterization of extracellular redox enzyme concentrations in response to exercise in humans. J. Appl. Physiol. 2019, 127, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Trewin, A.J.; Parker, L.; Shaw, C.S.; Hiam, D.S.; Garnham, A.P.; Levinger, I.; McConell, G.K.; Stepto, N.K. Acute HIIE elicits similar changes in human skeletal muscle mitochondrial H2O2 release, respiration, and cell signaling as endurance exercise even with less work. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R1003–R1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadley, A.J.; Aldred, S.; Coles, S.J. An unexplored role for Peroxiredoxin in exercise-induced redox signalling? Redox Biol. 2016, 8, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galli, D.; Carubbi, C.; Masselli, E.; Vaccarezza, M.; Presta, V.; Pozzi, G.; Ambrosini, L.; Gobbi, G.; Vitale, M.; Mirandola, P. Physical Activity and Redox Balance in the Elderly: Signal Transduction Mechanisms. Appl. Sci. 2021, 11, 2228. https://doi.org/10.3390/app11052228
Galli D, Carubbi C, Masselli E, Vaccarezza M, Presta V, Pozzi G, Ambrosini L, Gobbi G, Vitale M, Mirandola P. Physical Activity and Redox Balance in the Elderly: Signal Transduction Mechanisms. Applied Sciences. 2021; 11(5):2228. https://doi.org/10.3390/app11052228
Chicago/Turabian StyleGalli, Daniela, Cecilia Carubbi, Elena Masselli, Mauro Vaccarezza, Valentina Presta, Giulia Pozzi, Luca Ambrosini, Giuliana Gobbi, Marco Vitale, and Prisco Mirandola. 2021. "Physical Activity and Redox Balance in the Elderly: Signal Transduction Mechanisms" Applied Sciences 11, no. 5: 2228. https://doi.org/10.3390/app11052228
APA StyleGalli, D., Carubbi, C., Masselli, E., Vaccarezza, M., Presta, V., Pozzi, G., Ambrosini, L., Gobbi, G., Vitale, M., & Mirandola, P. (2021). Physical Activity and Redox Balance in the Elderly: Signal Transduction Mechanisms. Applied Sciences, 11(5), 2228. https://doi.org/10.3390/app11052228