1. Introduction
In the last few years, bioelectronics, i.e., the branch of electronics which conjugates biological and inorganic matter, is assuming a relevant role in green technology. Besides the speculative interest of investigating with electronic/electrochemical methods how biological matter interacts with the environment, a major interest in bioelectronics is to suit some peculiar abilities of living matter (for example, to identify some specific molecules, to detect and use light, and so on) for practical applications, from energy harvesting to sensing action. In this framework, an increasing number of biomolecules, mainly proteins and aptamers, have been selected, produced, and analyzed with different experimental and theoretical techniques to be part of electronic devices [
1,
2,
3,
4].
Photosensitive proteins can convert electromagnetic energy into different forms of energy useful for their surviving. After illumination, proteins involved in photosynthesis inside the cell activate an electron transfer that, in turns, activates the ATP-ADP cycle transformations, also producing a proton pumping across the cell membrane [
4]. Due to their specific abilities in charge transfer, these proteins are expected to exhibit a higher conductivity with respect to other photosensitive proteins. This is the case of type I opsins, which do not transfer electrons after illumination, but simply implement an ion pumping. Consequently, proteins involved in photosynthesis are among the most studied proteins for electrical/electronic applications [
5,
6,
7,
8].
Both experimental and theoretical investigations are in progress to shed light on the intriguing mechanisms behind biomolecule activation. Among the different theoretical modelling, proteotronics aims to mimic the set of microscopic mechanisms which, inside the biomolecule, regulate its behavior [
9]. Within this framework, charge transport in bacterial Reaction Center (b
RC) of
Rhodobacter Sphaeroides is here investigated with the purposes of interpreting available data and suggesting new fields of applications. Specifically, the analysis is concerned with some structures of b
RC deposited in the protein data bank [
10] and obtained in different conditions of pH [
11].
All proteins, and in general living matter, continuously interact with their environment, i.e., they are open systems. Consequently, the task of providing a comprehensive model is still far from present possibilities. As an example, temperature, hydration, and of course, the presence of specific ligands, sensitively affect the protein stability and activity, producing a very complex framework of energy landscapes. Indeed, the protein structure is the result of an amino acids sequence (primary structure) which arranges to reach a minimum of the free energy, thus competing with entropy [
12]. This means that thousands of equivalent structures should be explored before a shortlist of the most reliable could be obtained. From one side, the landscape of protein energy states is quite complex and not still complete; it assumes that after folding the protein lives in the ground state of an energy funnel, in which it assumes the most stable configuration. From another side, it is not clear to what extent the configuration is stable with respect to the variation of different environmental conditions, or if a degeneracy in energy exists with different proteins living in different equivalent auto-states [
13]. The activation (due to energy/ligand binding) should shift the protein toward a different, inequivalent, energy funnel [
14]. Waiting for a conclusive description we can only argue in terms of the statistically most relevant structure as given, for example, by the crystallographic investigation (which produces the output after many crystals analysis) and that strictly refers to specific environmental conditions. When these conditions also change the output changes, often in a way that is poorly appreciable to the naked eye but that can induce severe transformation in the protein functioning. Photosensitive proteins are quite interesting in this respect, because the topological transformation they undergo after they have absorbed light energy can appear quite small but produces an indisputable change in their functioning, which has been detected, for example, as an increase of their conductivity [
15,
16,
17]. Due to the difficulty in performing protein crystallization and analysis, available data refer often to a specific combination of environmental conditions and a comparative analysis of the effects of tuning these conditions is quite difficult to be found.
Anyway, reduced, coarse grained models could capture the system dynamics in at least selected sets of environmental conditions. It is reasonable to suppose that an analysis of the protein around a minimum of the free energy, performed by producing small perturbations of only one significant variable, should drive the protein in a sub-manifold of energy states mainly ruled by that variable. Accordingly, in the present analysis, we focus on the role of pH on proteins in the native state (without illumination) and in the same temperature conditions, 100 K, which is well below the physiological temperature. Results are influenced by these conditions and the possibility that at increasing temperatures thermal fluctuations play a significant role cannot be excluded, and thus become the subject of desirable future investigations.
Proteins are influenced by hydration, from the very initial stages of folding to the establishment of the native configuration and eventually of the active configuration [
18,
19]. Therefore, it is not surprising that a relevant change in the amount of H
+ cations in the solution in which b
RC is present could suffice to arise the electrostatic field around the protein thus inducing a rearrangement of the electrically exposed surfaces. This phenomenon is of great practical interest since the conformational change of the protein structure induces a change of its electrical properties. Therefore, it becomes of relevant interest to investigate these conformational changes and evaluate their electrical effects in the perspective of developing new electronic devices. To this purpose, the role of pH is here investigated with the objective to find a correlation with an electrical response that, at this stage, can also serve to explore the protein structure. As previously mentioned, the energy state of the protein, and therefore of its conformation, depends on the environmental conditions. While it is easy to understand that, at least in the physiological range, the lowering of the temperature pushes a protein toward its minimum of energy, an analogue trend of the pH value is not a priori established. It is even not clear to what extent temperature and pH may influence each other. Starting from a set of data coming from crystallographic investigations performed on b
RC at different values of pH at the same temperature of 100 K [
11], the present investigation allows a clear separation of the effects of temperature and pH, thus avoiding possible effects of correlated influences.
Investigations on the electrical performances of biomolecules have a long tradition and cover different methodologies. Results of particular interest come from the electrochemical impedance spectroscopy (EIS), and the Conductive Atomic Force Microscopy (CAFM). The former one mainly performs analysis on macroscopic samples (typically with active surfaces of some cm
2) in strong coupling with a buffer/electrolytic solution and in the presence of a sufficiently weak AC applied voltage. Results account for both the sample and solution response and it is in general not easy to discriminate the contributes to the total impedance. Electronic models are continuously updated to give a microscopic interpretation of the very complex physico-chemical processes [
20,
21]. As a matter of fact, many devices, for example sensors, are characterized by the EIS technique, therefore the modelling of results is relevant for both the speculative and applicative aspect [
22]. Experiments performed with the EIS technique detect a sensitive enhancement of the cell conductance in light [
17,
21]. Specifically, it was observed that the charge transfer involves the protein, the quinones in solution and depends on the electrode. Furthermore, a sensitive magnification of conductance is observed in light.
CAFM can measure currents from a few to a single biomolecule appropriately bound on a conductive substrate in the presence of significant strong and DC applied electric fields. In this case as well, the experimental setup plays an important role in the result, for example, the specific protein anchoring [
12], or the measurement mode (for example, contact or tapping mode) [
15,
16,
23]. Photosensitive proteins sandwiched between an electrode and the CAFM tip have been studied under different conditions of illumination showing, in dark, an unexpectedly high conductance [
12,
23], which can be further enhanced in the presence of light [
15].
Concerning the theoretical interpretation of experimental results, we refer to the proteotronics, a recent branch of molecular electronics developed inside our research group since 2014, that evaluates the electrical properties of a protein by using an impedance network analog based on the protein tertiary structure [
9,
24]. By using this approach, we succeeded to reproduce and interpret experimental data performed in neutral or quasi-neutral pH condition and foresee the results in alkaline conditions. In particular, the results of the investigation predict an increase of conductance in analogy to that given by illumination, and that can be useful for applications in sensor devices.
3. Results
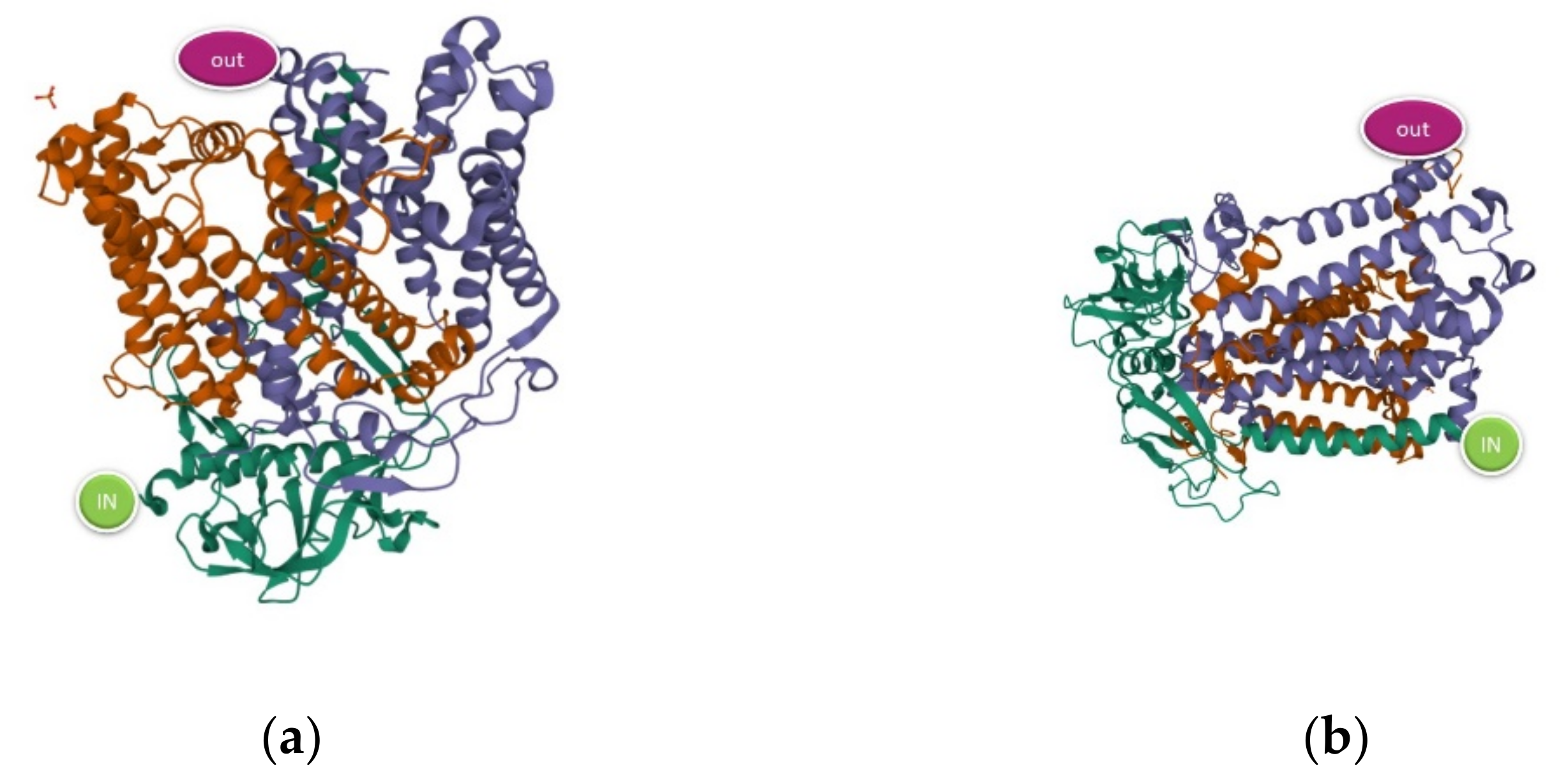
The theoretical model refers to a single protein taken as representative of the macroscopic electrical response. The role played by the position of the contacts can be of relevance for the results of both experiment and theory. Therefore, within a first estimate, we will consider two possible contact geometries, a first up-down as in the cell arrangement, and a second tilted of π/2, as it can happen in a nonhomogeneous sample and is visualized in
Figure 1.
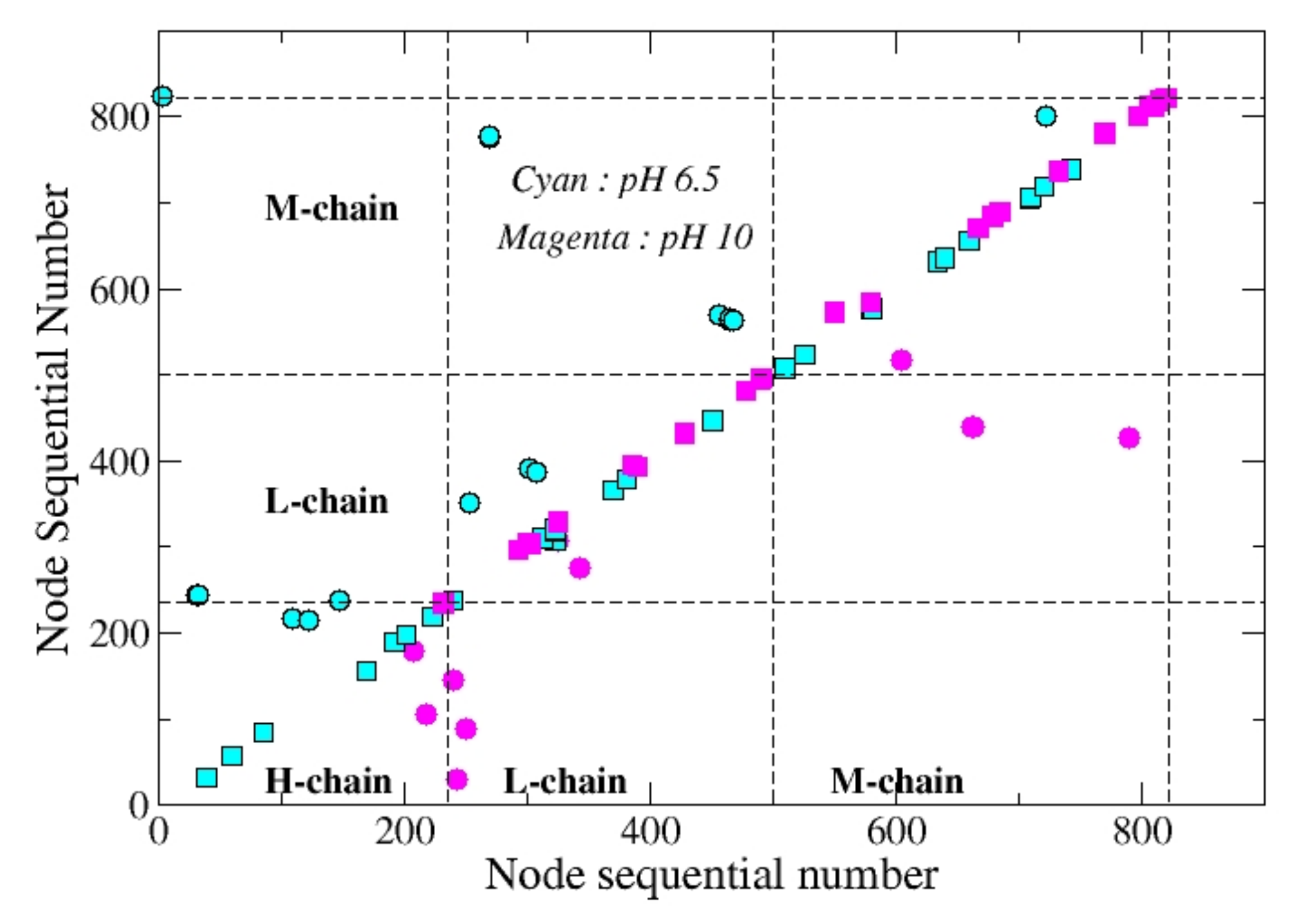
While sensitive structural differences are not impressive to the naked eye, they appear more clearly when the network analysis is performed. As an example, in
Figure 2 we report the set of contacts which characterize the structure in dark at two different pH values. Elements on the diagonal correspond to sequential amino acids. This figure shows that the main differences between the pH = 6.5 structure and the pH = 10 structure are mainly due to the position of sequential amino acids. As a matter of fact, at pH = 6.5 more contacts appear on chain H while at pH = 10 more contacts are on chains L and M. In other terms, by increasing the pH the protein deforms from bottom to top, thus becoming tighter toward the periplasmic side.
In the following we investigate two charge-transport regimes that are usually considered by experiments. To this purpose, a first subsection is devoted to the linear response regime, typically adopted within the EIS technique, and a second subsection is devoted to the non-linear transport regime, typically adopted within the CAFM technique. We assume as a benchmark the bRC structure in dark at pH = 6.5 (PDB code 2uww), as well as its impedance value.
3.1. Linear Response Regime
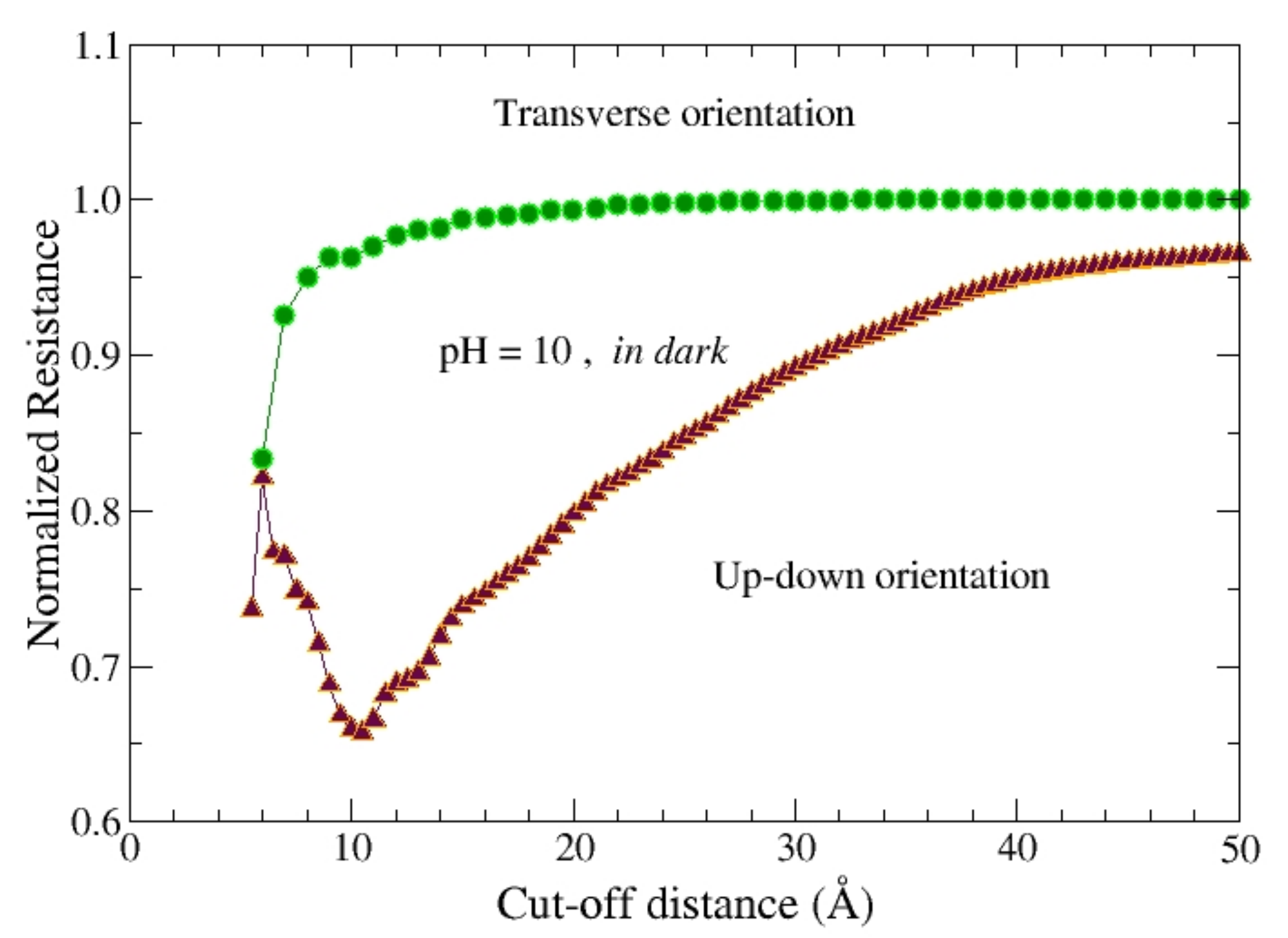
Under linear response regime all the parameters entering the local impedance in Equation (1) do not depend on the applied bias. Accordingly,
Figure 3 reports the normalized resistance of the single protein in dark, R (#)/R, i.e., the static (DC, i.e., ω = 0) global resistance R (#) of the protein, normalized to the benchmark value R, in dark, as a function of the cut-off distance between neighborhood nodes, D. In particular, the protein at pH = 10 (PDB code 2uxj) is compared with the protein at pH = 6.5 (PDB code 2uww). Calculations have been performed by using two different contact configurations as reported in
Figure 1.
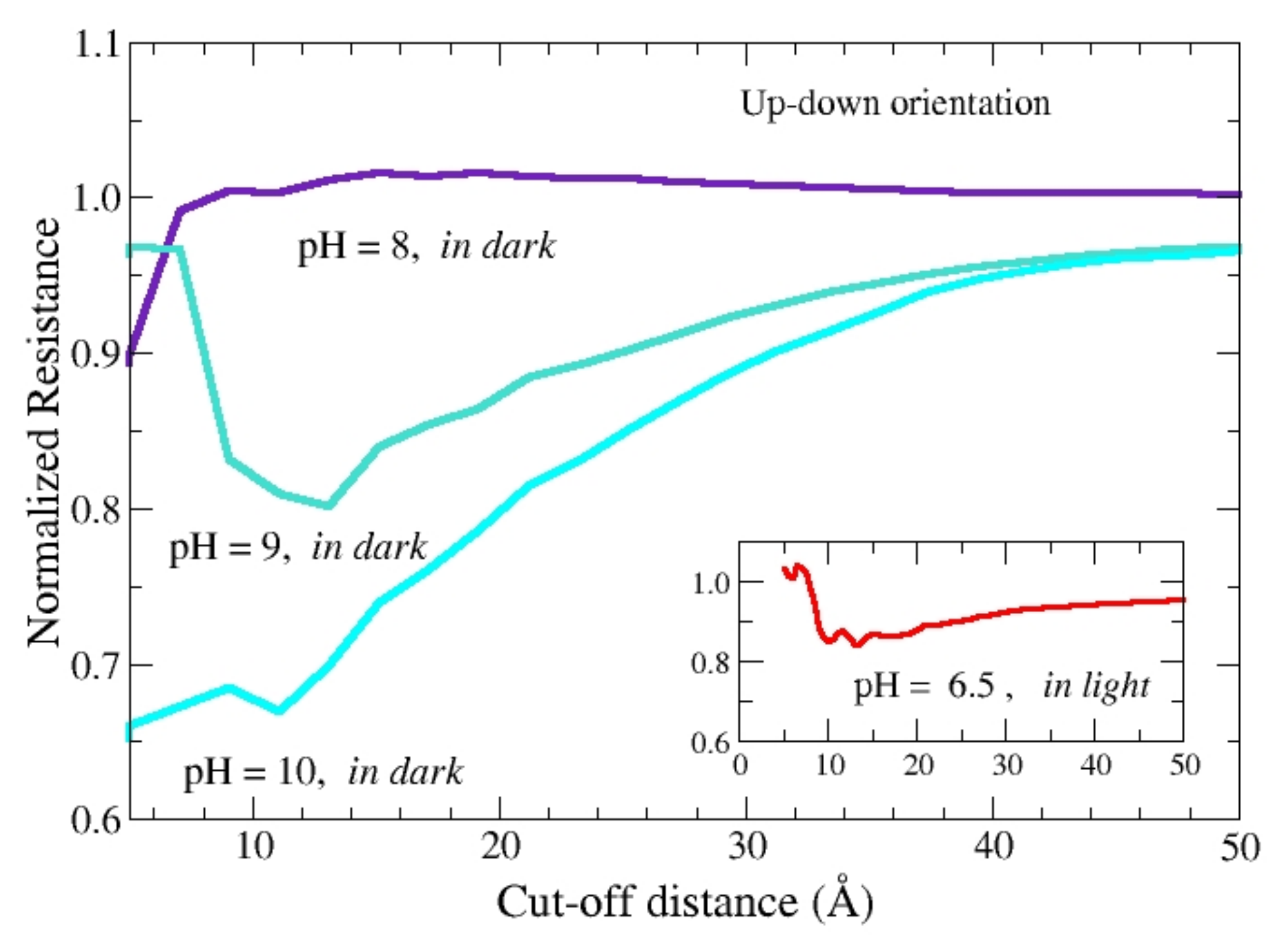
From the above comparison we conclude that the value of the pH plays a significant role only in the up-down orientation, where the value of the resistance stays well below that of the benchmark. By using the up-down orientation,
Figure 4 reports the normalized resistance of the single protein, R (#)/R, calculated at different values of pH. A higher sensitivity is observed at increasing values of pH. In particular, the best resolution is observed for the value
D = 10.5 Å of the cut-off distance, that compares well with the value
D = 6 Å found for the case of bacteriorhodopsin (bR), a protein like b
RC but with a smaller number of amino acids for about a factor of 4 [
14]. In the inset of
Figure 4 the resistance of the protein in light at pH 6.5 is compared with the benchmark resistance. In this case, the maximal resolution also shows up for a cut-off value
D = 10.5 Å, with a reduction of resistance in fair agreement with experiments [
12].
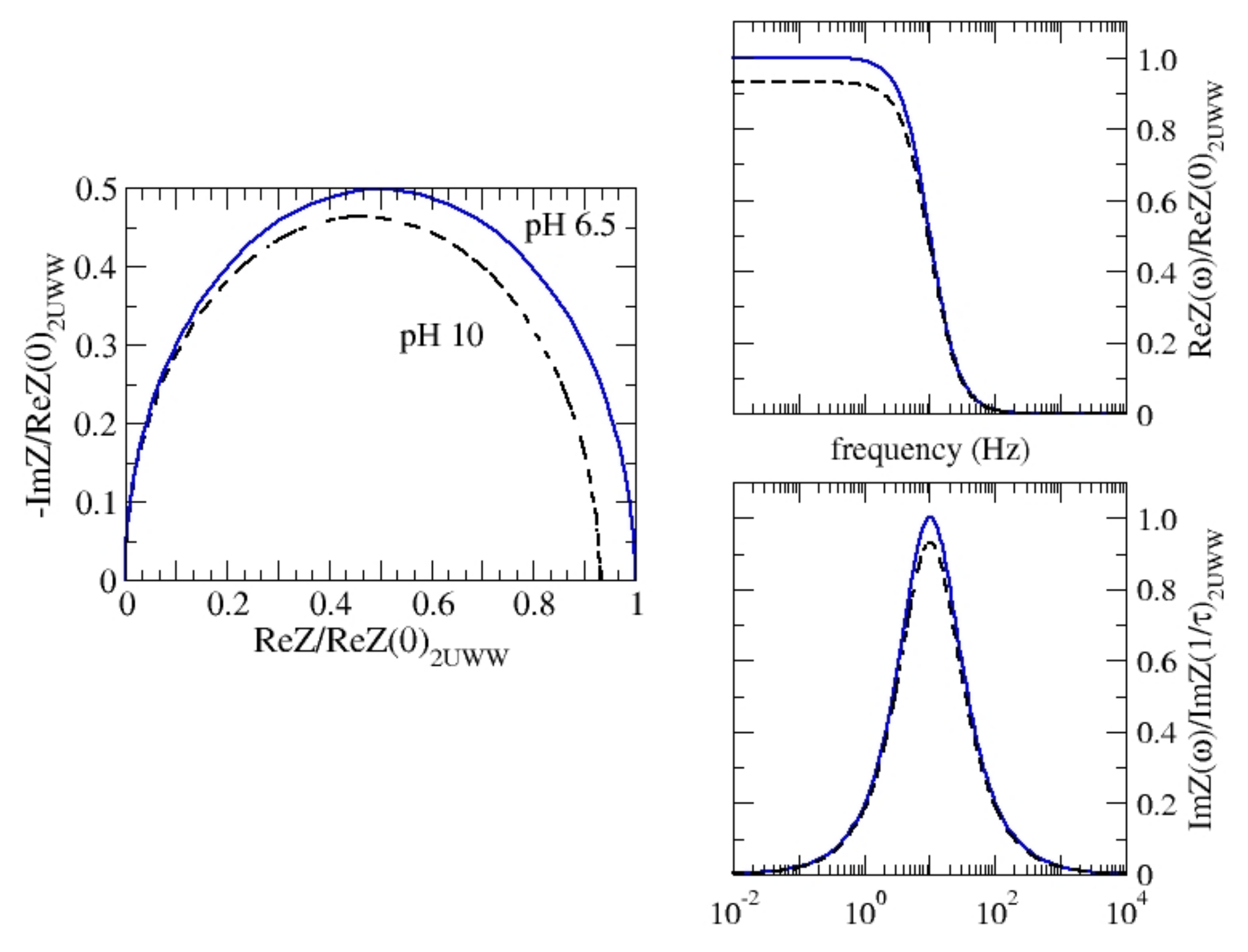
Finally, making use of the up-down orientation in dark, and taking
= 2.7 × 10
12 Ω Å we evaluate the complete impedance spectra for pH = 6.5 and 10 respectively, in the frequency range (10
−2–10
4 Hz) which covers the full experimental range. In this range, experiments detect the solution effects, mainly at the highest frequencies, and the electrode resistance, at the lowest frequencies. Otherwise, all the cell elements contribute to the global impedance response in the full frequency range [
20,
21]. The impedance model may be complex to account for several effects that could be detected by experiments. Anyway, the main contribution is given by Equation (1) and accounts for both the polarization and some leakage currents which are usually present in real dielectrics [
9,
14,
21,
25]. In an electrochemical cell conduction there is both an electronic and ionic origin, i.e., charge may travel through both organic/inorganic matter and the electrolytic solution. Furthermore, charge may accumulate between different solid/liquid or solid/solid interfaces [
21]. Equation (1) describes an elemental circuit consisting of a resistance (R) and a capacitor (C) in parallel, R//C, and its graphical representations in terms of frequency are called Bode plots. The imaginary vs. the real part of impedance is represented by the so-called Nyquist plot. Calculated Nyquist and Bode plots could be attributed to an ideal single R//C circuit (see
Figure 5), which means that the elemental R//C circuits in the protein (each of them representing a single link) exhibit a synchronous response. In other terms, a single relaxation time is detected, the same for both configurations, τ = 0.015 s. This value is compatible with the experimental data reported in the literature [
21]. With different configurations of geometries, their elemental capacitances exhibit different values that are larger for the case of a pH = 10, in fair agreement with the observation of a tighter shape with respect to the pH = 6.5 configuration.
3.2. Super-Linear Response Regime
To reproduce the super-linear behavior exhibited by the current-voltage (I-V) experiments obtained with a CAFM technique, at increasing values of an applied positive-voltage [
12], the microscopic model makes use of a sequential tunneling mechanism of charge transfer between different nodes that are assumed to be separated by an energy barrier Φ. Accordingly, the link resistivity,
is chosen to depend on the local voltage drop
between the couple of
j,
k amino acids involved in the link, as:
Specifically, the current response is simulated by using a stochastic Monte Carlo procedure to allow the charge transfer channels to reduce their initial resistivity from a
value to a
value at increasing of the potential drop between nodes [
14,
25,
26,
27,
28,
29]. This is on the wake of the well-known Simmons model for the charge injection in an electronic junction [
30]. Indeed, the model describes two different tunneling regimes: at low bias it envisages a direct tunneling mechanism, at high bias overtaken by an injection tunneling mechanism. At low bias, the condition
holds for most of the channels that take the same
value, thus the global response of the protein is like that of a high resistance Ohmic conductor. At increasing bias,
, an abrupt drop of the resistivity to the minimal value,
, occurs for the given channel and the global resistance of the protein will decrease accordingly. At further increasing of the bias, most of the channels will take the
value and the global response of the protein will be like that of a low resistance Ohmic conductor. The tunneling transition probabilities including direct,
and injection, or Fowler Nordheim (FN), mechanisms,
, reads:
where
is the reduced Planck constant,
e is the unit charge, and
m is the electron effective mass, here taken the same of the bare value.
Current-voltage characteristics under different conditions are reported in the following
Figure 6,
Figure 7 and
Figure 8.
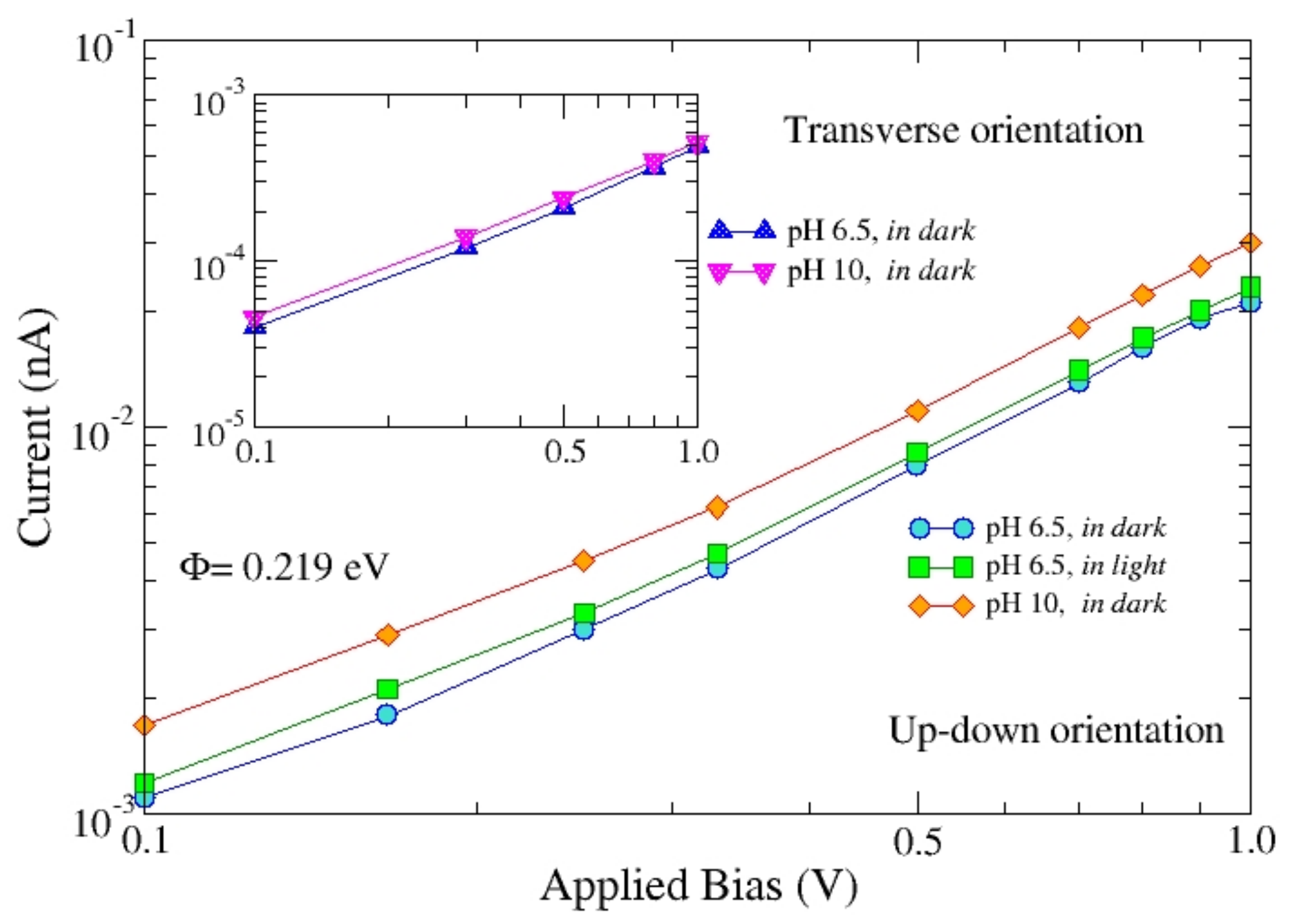
By using up-down and transverse orientations,
Figure 6 compares the
I-
V characteristics of b
RC in dark, at pH = 6.5 and 10. Furthermore, in the up-down orientation, the
I-
V characteristics of b
RC in dark and light at pH = 6.5 are compared. The current response of the up-down orientation is found to be of about 2 orders of magnitude higher than that of the transverse orientation. The sensitivity to the increase of pH is small for the transverse orientation, while it goes up to about 50% for the case of the up-down orientation (at 1 Volt). Calculations use the following parameters:
= 2.7 × 10
12 Ω Å,
= 2.7 × 10
4 Ω Å and Φ = 0.219 eV. These data confirm that the primary role of contact position does not disappear in the super-ohmic regime. On the other hand, they emphasize the small effect of illumination on the protein 3D arrangement.
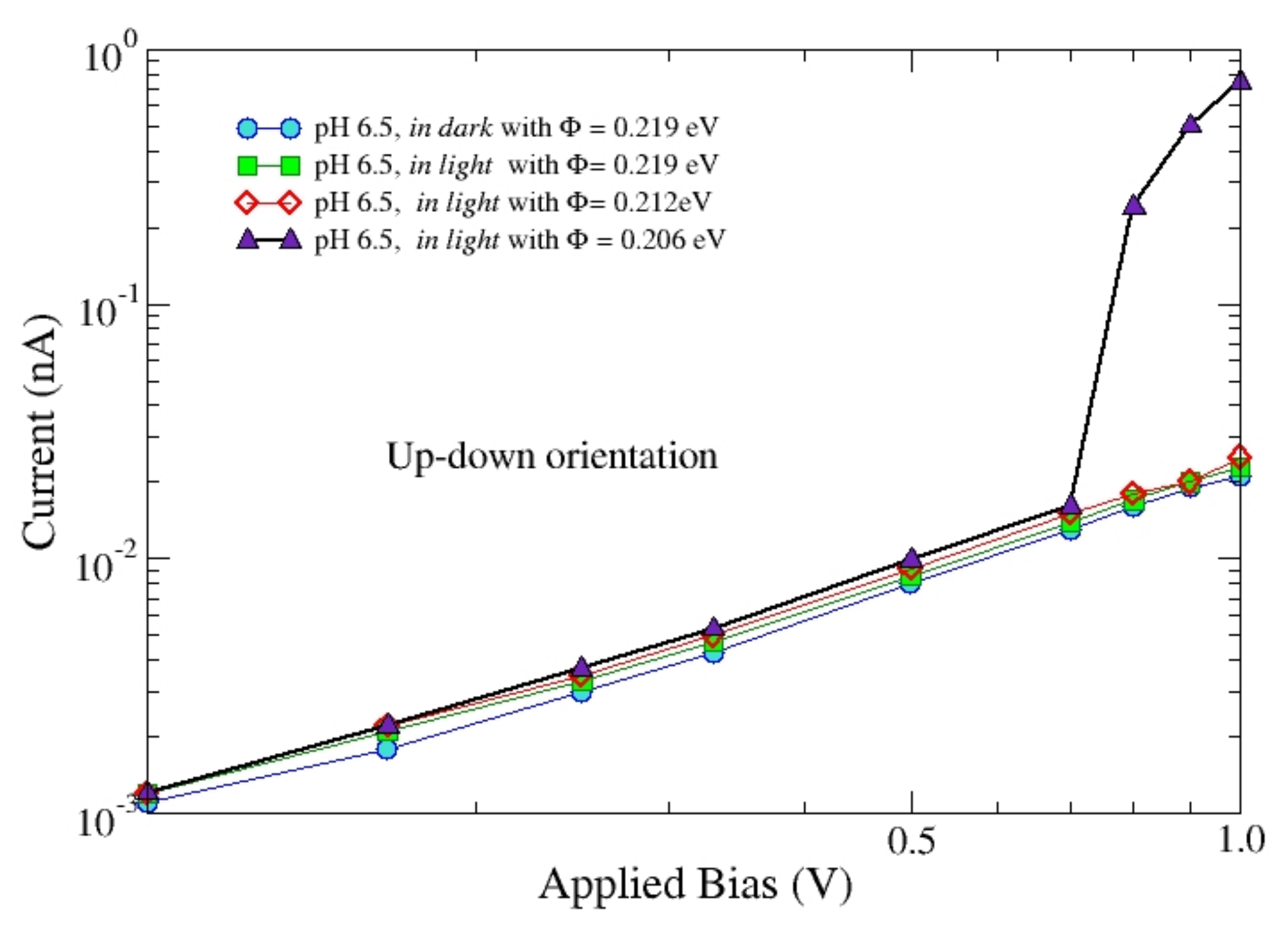
Figure 7 reports the
I-
V characteristics of b
RC in the up-down in dark orientation and in light at pH = 6.5 for a range of applied bias of interest for experiments.
Calculations make use of three values for the barrier height, Φ = 0.219 eV (squares), Φ = 0.212 eV (diamonds), and Φ = 0.206 eV (triangles). Even a small decrease of the barrier height is found to be responsible for a significant increase of the current in light for applied voltages above about 0.7 V, due to the anticipated onset of tunneling in the FN regime. By contrast, for the same value of the barrier height the results in light and in dark are practically the same apart from a small increase of the current of about 6%. These results suggest that in bRC, part of the light energy could be transferred to the electrons injected in the protein thus leading to a further increase of the current.
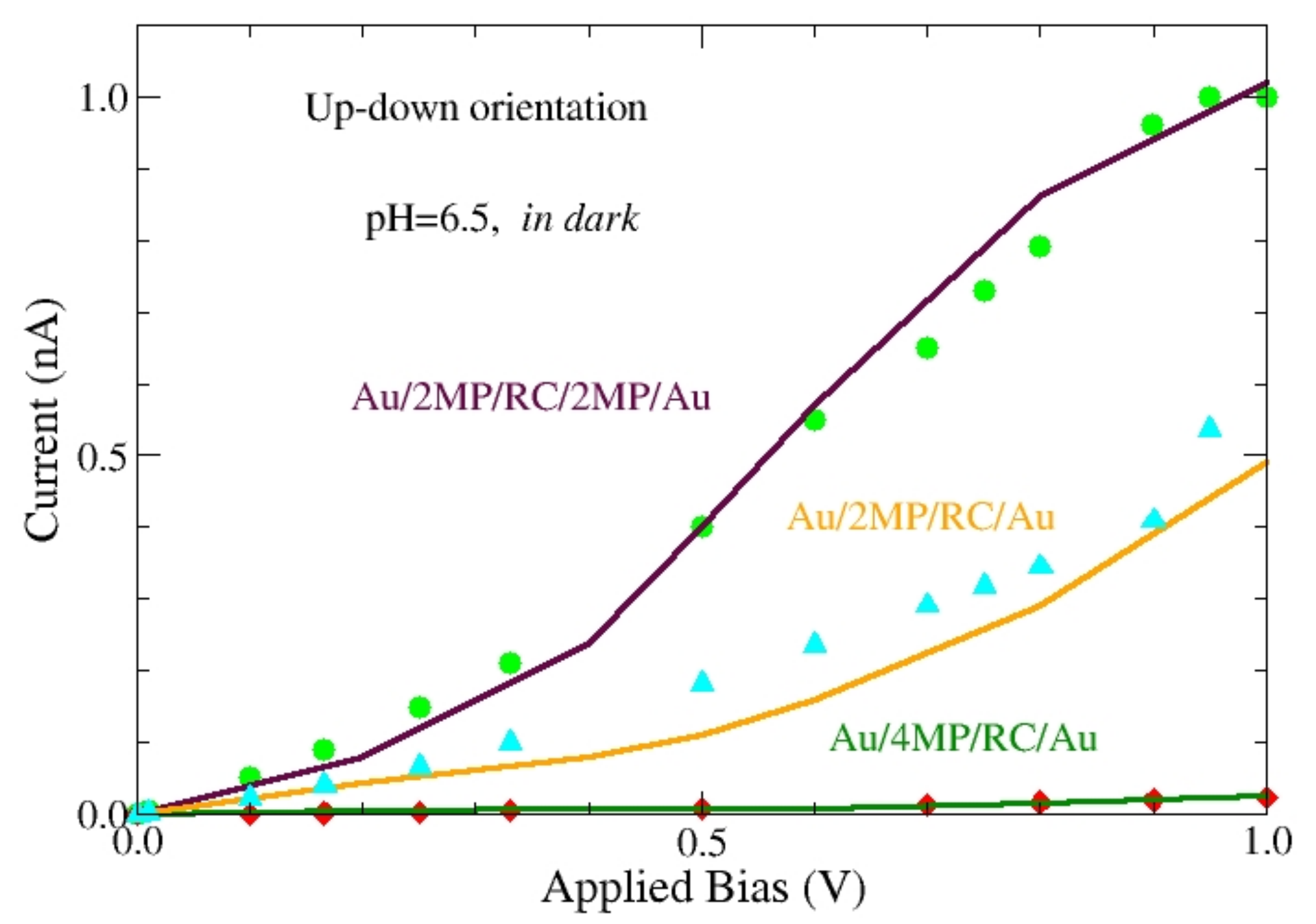
Figure 8 reports the
I-
V characteristics calculated in dark at pH = 6.5 for the protein in the up-down orientation (symbols) and are compared with experiments [
12], (continuous lines). In these experiments, performed with the CAFM, the protein was immobilized on an Au-electrode and contacted with the CAFM tip. Both the electrode and the tip have been coated with different chemicals. Here we focused on three different assemblies, say, Au/4MP/protein/Au, Au/2MP/protein/Au and Au/2MP/protein/2MP/Au, where 2MP indicates 2-mercaptopyridine and 4MP 4-mercaptopyridine [
12].
The satisfactory agreement between theory and experiments is obtained by using different values for
to account for the different chemical modifiers used to anchor the protein, while keeping the same values for the other parameters. Three different values of
have been used, namely: 2.7 × 10
12 Ω Å, 1.2 × 10
11 Ω Å and 5.4 × 10
12 Ω Å, to fit experiments performed by using, respectively, the Au/4MP/protein/Au, Au/2MP/protein/Au, and Au/2MP/protein/2MP/Au, assembly. This result suggests that the role played by the chemical modifiers of the electrode and/or the CAFM tip is to increase the effective conductivity of the protein and it is not due to the change of the potential barrier height. As a matter of fact, the shape of the
I-
V curve is practically the same for the three cases, while the absolute value is changed. Therefore, the reduced effective conductivity should be due to a better match of the Fermi level of the electrode with the LUMO/HOMO molecular orbitals of the protein, as suggested in Reference [
12]. As a matter of fact, having carried out the theoretical analysis with the same orientation (up-down), a realignment due to the chemical modifiers seems to be less relevant.
4. Discussion and Conclusions
By using the tertiary structure of the bRC protein, its electrical characteristics are investigated under dark and light conditions and at different values of the pH of the crystallized solution, ranging from weak acid up to strong alkaline values. Theoretical calculations are carried out in the framework of the proteotronics, a structure-minded approach to the recent field of protein-based electronics. Analysis explores the macromolecule electrical response in both linear and non-linear regime.
The procedure starts by mapping the macromolecule onto a graph which accounts for the protein structure. The graph is used to describe the electrical response of the protein in the philosophy of the lumped-element model, thus becoming an electrical network. A percolative-like process allows the analysis of the structure by using an ideal flow of charge that goes through the protein. The resistance that the network opposes to the charge flow measures its degree of connectivity and is a powerful tool to discriminate among different (although, sometimes, very similar) structures. In the present case, different pH values or different illumination conditions produce different networks, thus different electrical responses.
Specifically, in the linear regime, i.e., at sufficiently low values of an applied bias, calculations of the electro-chemical impedance spectroscopy suggest that bRC is significantly sensitive to the pH value of the solution in which it is crystallized. The protein undergoes a resistance reduction when the pH changes from weak acid (pH = 6.5) to alkaline conditions with pH values up to 10, where this change appears even more evident. In the non-linear regime, i.e., at applied voltage values up to few Volts, theory makes use of a stochastic Monte Carlo approach to mimic a charge transport based on a sequential-tunneling mechanism including direct and injected tunneling regimes. Current voltage characteristics evidence a super-linear behavior with an increase of the current above the Ohmic value up to about a factor of two, and to keep the sensitivity to the value of the pH and the presence of the light. The current-voltage characteristics compare qualitatively well with available experiments performed in quasi-neutral conditions and in dark.
These kinds of analysis should be of relevance for both basic and applied research. In the former case, the microscopic characterizations of proteins are still in their infancy when compared with simple inorganic materials, thus justifying the phenomenological approach used for the theoretical model. In the latter case, future application of these proteins in several branches of technology, medicine, solar energy harvesters, etc. are found to be very promising. From one side, they highlight the primary role of environment (here limited to the amount of cations on solution) in the physical-chemical (here electrical) response of biomaterial and how its effects are recognizable in the structure. Similar investigations concerning, for example, the role of temperature on structure and function of bRC, which should complement present data are at present not available, although desirable. From another side, they can be used, for example, to regulate resistance in such devices. Finally, the present investigation can be thought to provide a valuable hint for designing pH-sensors.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}