1. Introduction
Morphological integration (MI) is the covariation between phenotypic structures that provide background information for inferring connections between traits in the course of development or as a result of shared functional demands [
1]. Integration and modularity are related to the degree of covariation between the different components of an anatomical structure [
1,
2].
Morphological modules are those structures that have components that covary strongly, but at the same time are quite independent of other modules [
1]. Morphological integration is understood to be the coordinated morphological variation of the components of a functional anatomical structure [
1,
2].
For the study of MI, the patterns of covariation are usually analysed by Partial Least Squares (PLS) and Escoufier RV coefficient [
3]. The multiset RV coefficient is a generalization of the RV coefficient which quantifies the association between two groups of variables [
4]. Both RV and PLS coefficient tests use the covariance matrix, even though both tests are different and respond to the question of covariation in different ways [
5].
The RV coefficient indicates the total amount of covariance, scaled by the amount of variation between two groups of variables [
2]. To evaluate the statistical significance of the obtained RV coefficient, permutation tests are used, as for example to simulate the null hypothesis of complete independence of the observations in the two groups of variables are randomly permuted.
The PLS is a multivariate analysis that uses the decomposition of the singular value of the covariance matrix between two partial warps matrices, corresponding to two forms (A and B) that are compared. Between them both, the covariation pattern is summarized by two groups of latent vectors (linear combinations), one for each block [
2]. This type of statistical test is analogous to the canonical correlation test, but in contrast to it, the vectors are not perpendicular to each other. PLS analysis produces conformation correlations between structures in an individual.
RV coefficient and PLS tests are two complementary statistical methods for evaluating possible covariance patterns, as the first analyses the total covariance between two matrices in relation to the variance of each matrix, whereas the second focuses on two of the maximal covariance axes [
3].
There have been few studies of bovid post-cranial osteology applying geometric morphometrics, most having been focused on autopodia [
6,
7]. Among ruminants, the hindlimb basipodium, or tarsus, comprises five bones: the astragalus (talus) and calcaneum, os centroquartale [
Os naviculocuboideum],-fused os tarsi centrale [
Os naviculare] and os tarsale IV [
Os cuboideum], os tarsale I [
Os cuneiforme mediale and
mediointermedium] and os tarsale II and III [
Os cuneïforme intermediolaterale]-fused os tarsale II [
Os cuneiforme intermedium] and os tarsale III [
Os cuneiforme laterale]- [
8,
9]. The calcaneum, known in ancient times as “peroneal tarsal bone”, presents a distal portion of its body compressed laterally, and a proximal projection which articulates with the lateral malleolus. The calcaneal tuberosity-
tuber calcanei-is well marked and on its plantar aspect it presents a wide and shallow groove -
sulcus calcanei- which is covered with cartilage in the living animal [
8]. All the bones of the foot are formed from a primary centre of ossification and the phalanges, metatarsals and calcaneum also have a secondary centre [
10]. The two ossification centres of the calcaneum are one for the body and the process of the calcaneum and the other for the distal half of the bone (
tuber calcanei) [
9]. Although the talo-calcaneal-navicular joint is usually considered a single joint complex, which is justifiable from a functional point of view, it is not so from an embryological point of view [
11], indicating the possibility of modularity of these two parts (body and the process of the calcaneum and the
tuber calcanei).
We are unaware of any study devoted to the MI on bones of the sheep tarsus, although recently we have published an article comparing calcanea of sheep and goats using geometric morphometric techniques [
12]. Here we attempt to understand whether a division between the tubercle and articular base on the calcaneum would have a modular foundation, as well as the respective level of morphological integration between both structures. The hypothesis of two-modularity of the development of the calcaneum was tested, using the RV coefficient. Then, the degree of morphological integration e between these two modules, which we defined above, was evaluated through an analysis of two PLS.
2. Materials and Methods
2.1. Sample
The data collected comes from 47 domestic sheep (Ovis aries) calcanea (os tarsi fibulare) belonging to several breeds housed in the Laboratório de Arqueociências of the Direção Geral do Património Cultural (Lisbon, Portugal) and in the National Museum of Natural History and Science (Lisbon, Portugal): Churra da Terra Quente (n = 15), Merina branca (n = 2), Merina preta (n = 4), Merina (n = 7), Soay (n = 1), and “unknown” (n = 18). We considered for the analysis only adults (tuber calcanei completely fused) and subadults with the tuber calcanei fused with epiphysial line visible. Right calcanea were measured, however, in the case of pathological broken, or missing right calcaneum the left one was used in its place. In these cases, to obtain a right-oriented calcaneum, a reflected image of the left calcanea was used. Pathological specimens were not studied. These photographic documents are kept in the fourth author’s files.
2.2. Processing of the Primary Data: Geometric Morphometrics
Photographs were taken as described in our previous article [
12]. They were taken orthogonally to the medial aspect of each calcaneum at 30 cm of distance between the camera and the calcanea. We used a Nikon
® D3400 (Nikon, Barcelona, Spain) digital camera in a fixed position equipped with an 18–55 mm objective set at a focal distance of 55 mm. To control the position and inclination of the camera, we used a spirit level, which ensured that the camera was situated identically in all photographs. A scale was included in each photograph to facilitate calibration by the software. We saved each image in JPG format.
A set of 20 2D coordinates of homologous anatomical landmarks was collected on the fibular aspect (
Figure 1) using the tpsDig v. 2.16 program [
13]. Landmark definitions are: (1) Most concave point below the
tuber calcanei; (2) Craniodorsal point of the
calcaneal tuber; (3) Most dorsal point of the
calcaneal tuber; (4) Caudoventral point of gastrocnemius enthesis; (5) Intersection point between the calcaneum neck and
sustentaculum tali; (6) Uppermost dorsal point of
sustentaculum tali; (7) Most caudal point of
sustentaculum tali; (8) Most cranioventral point of
sustentaculum tali; (9) Uppermost dorsal point of dorsal astragalus articular surface (
facies articulares talares); (10) Most cranial point of dorsal face of
facies articulares talares; (11) Most concave point between landmarks 10 and 12; (12) Most cranial point of malleolus articular surface; (13) Most concave point between landmarks 12 and 14; (14) Most ventral point of
facies articularis malleolaris; (15) Most concave ventral point to
facies articularis malleolaris; (16) Most cranioventral point of inferior
facies articulares talares; (17) Most cranioventral point of cubonavicular articular surface (
facies articularis cuboidea); (18) Most concave point of the caudal edge of the
facies articularis cuboidea; (19) Most caudal point of
facies articularis cuboidea; (20) Most concave point of the cranial edge of the
facies articularis cuboidea. Nomenclature has been adjusted to
Nomina Anatomica Veterinaria [
14].
Previously, to carry out any analysis, the consistence of the measurements (photographic and landmark positioning) were evaluated, and a repeatability experiment was conducted. Given that the last author of the present study is the person who developed the anatomical revision of the sheep calcanea based on geometric morphometrics [
12], intra-observer error was carried out by him. This was done repeating photographs of two specimens 10 times during 10 different days. During this period, when each repeated photograph was taken, the tripod and the camera were mounted and dismounted each time. In addition, we also randomly selected 10 photographs of 10 different specimens from our initial sample (47 calcanea). Then, on the 30 photographs, the 20 landmarks were located twice during a 10-day period. After digitalisation, we used a Procrustes ANOVA test both among and within repeated calcanea photographs to statistically evaluate the measurements consistency [
12].
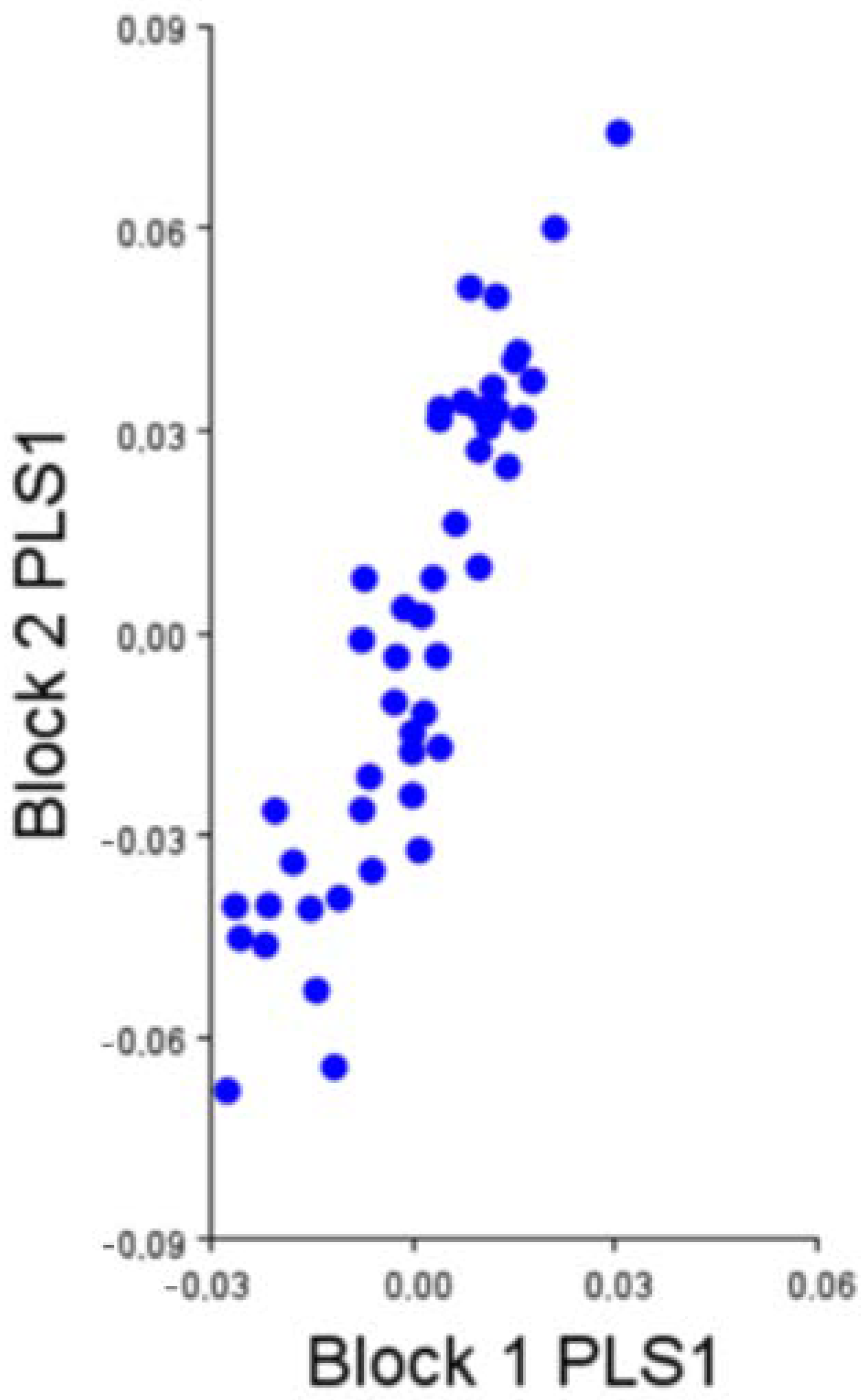
To test a hypothesis of bi-modularity of the development of the calcaneum, we divided the 20 2D landmarks into two different modules or blocks to describe the changes in size and shape of structure: one for the body and the process of the calcaneum (block 1 expressed by 4 landmarks: from landmark 1 to 4) and the other for the distal half of the bone (including talocalcaneal joint) (block 2 expressed by 16 landmarks: from landmark 5 to 20).
Differences due to rotation, translation and scale were eliminated for each module through a Generalized Procrustes Analysis (GPA). As a size estimator, Centroid Size (CS) is used [
15] (Bookstein 1991). CS is obtained by calculating the square root of the sum of the distances, raised to the square, from each anatomical point to the centroid [
1].
2.3. Allometry
Allometry (i.e., change of shape versus size) was studied in a multivariate regression using log-transformed CS to independent variable and shape scores as the dependent variable (number of randomization rounds: 10,000) for both modules independently.
2.4. Modularity
The modularity hypothesis was tested before carrying out analyses to evaluate canalization and morphological integration, using the method proposed by Klingenberg [
16]. The RV coefficient was computed to quantify the strength of the association between two subgroups of the landmarks. The RV multiset coefficient was obtained from the aligned coordinates of the different components and for 10,000 random partitions of the landmarks containing the same partitions (number) of landmarks. If one component has a significant modular structure in relation to another, that is, if they can be considered independent modules, the RV is expected to be less than 95% of the randomly obtained partitions (i.e., the covariation between the components will be significantly less than the covariation within each of the components) [
5].
2.5. Partial Least Squares
The Partial Least Squares (PLS) statistical test is useful to evaluate possible covariation patterns between two or more variable groups. In morphometry, this test is useful because at least one of these variables has information about the shape. So, PLS allows us to relate this form information to other kinds of data such as genetic conditions, molecular information or other shape variables [
5]. In fact, this test looks for the greatest covariation between two sets of variables through correlating pairs of linear combinations between these two sets of variables, which are vectors [
5]. The main goal of this test is to express the morphology of an object in a few dimensions of the covariance structure between these variables sets.
For all statistical analyses, MorphoJ v.1.07a [
17] and PAST v. 2.17c [
18] packages were used. A 95% confidence level was established for all tests.
4. Discussion
A module can be defined as a morphological unit within which there exists a high integration of several and/or strong interactions, but that at the same time they possess a certain degree of independence from these units [
1,
2]. Modularity describes the case in which the covariation patterns of traits are unevenly dispersed among them. It has been a topic of particular relevance in the last decade, especially in studies of developmental evolution, where several authors argue that it provides sufficient flexibility so that different traits evolve more easily under different functional demands [
2,
19,
20].
The calcaneum is the largest of the tarsal bones and articulates with the astragalus (talus) and some tarsal bones [
9]. The proximal articulation with the astragalus is extremely stable, with the two bones almost morticed together [
9]. Distally it articulates with the central and fourth tarsal bones [
9]. The effects of morphological integration were not very evident in the two modules considered on the calcaneum: tuberosity and base. However, while these two modules have a certain degree of modularity, both vary with much greater independence.
Both the astragalus and calcaneum ossify before birth in
ossa tarsi [
21]. The calcaneum develops from two different centres, one for the body and the process of the calcaneum and the other for the proximal half of the
tuber calcanei [
21]. This part, the
tuber calcanei, provides attachment for the common calcaneal tendon [
9]. The centres for the body and the process of the calcaneum appear before birth, the remaining centres appear after birth [
21]. Based on the evidence presented here, they appeared highly modularly structured, and their morphological integration was tested positively, although with a medium covariation.
It also seems to be clear that the components of the calcaneum are integrated between them, although they have different embryonic origins. In this way it was possible to show that the two modules do seem to exist but that they behave in a rather dependent way. We think this is due to the fact that they must evolve, develop and function as a joint and so in a coordinated way, as hock muscles insert onto different parts of the calcaneum: the tendons of the
triceps surae (gastrocnemius + soleus) and superficial digital flexor, and tarsal tendons of the
biceps femori and semitendinosus muscles make up the common calcanean tendon -the “hamstring” of quadrupeds- [
22] so the morphological integration of both parts of the calcaneum seem to share functions rather than precursors of development. Changes in this anatomical system and the coordination among its different elements are probably focused on a fitness finality. In fact, the morphology of the calcaneum reflects the functional adaptation of this anatomical element to the lifestyle of artiodactyls [
23]. To combine studies on biomechanical and morphological analysis can be informative on this anatomical system and its two-calcaneal modules. This kind of approach is interesting and helps to improve our understanding of the morphological integration of the sheep calcaneum.
Finally, we show by this specific example on the sheep calcaneum, how appropriate the application of geometric morphometrics is to study the phenomena of modularity and morphological integration. These statistical methods offer high flexibility, statistical power, and good shape visualization, along with well-developed and coherent mathematical foundations. The combination of geometry and multivariate statistical methods guaranteed a characterization broad and not redundant of the shape structure under study, and subsequently an adequate representation of the morphologies of interest, allowing the quantitative and multivariate analysis of the fact of morphological integration and modularity in joint bones.
This is the first time that the relative independence of the development of two modules of the calcaneum has been demonstrated. Due to this, and the lack of previous studies on the patterns of integration and modularity related to bone development it is not possible to compare the results of the present study with previous results. This study is an example of the information that studies on the patterns of integration and modularity give for the development of osseous processes.
5. Conclusions
This is one of the very few studies dealing with sheep calcanea. To perform this study, we used GM of landmark-based data, demonstrating how appropriate this methodology is in studying the phenomena of modularity and morphological integration in development processes.
The results obtained constitute the first which elucidates the relative independence of the development of two modules of the sheep calcaneum. According to our results the two centres of ossification existing in this bone correspond to two modules of development with a medium covariation. Despite having different embryonic origins, these two modules are integrated. This morphological integration of both parts of the calcaneum is probably due to the fact they share biomechanical functions.
This study shows the power that studies of the patterns of integration and modularity have to put in evidence the underlying development processes of an anatomical structure. Future work on this topic should try to elucidate other possible expressions that underlie each of these specific patterns of covariation to improve our understanding of the ontogenetic sequence followed during the conformation of these modules.








{kind=link}
{kind=link}
{kind=link}