Inactivation Kinetics of Salmonella typhimurium and Staphylococcus aureus in Different Media by Dielectric Barrier Discharge Non-Thermal Plasma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Sample Preparation
2.2.1. Preparation of Bacteria Embedded on Solid Media
2.2.2. Preparation of Bacterial Suspensions
2.2.3. Sample Packaging
2.3. Plasma Treatment
2.4. Microbiological Recovery and Analysis
2.5. Bacterial Inactivation Model and Sublethal Injury Estimation
2.6. Statistical Analysis
3. Results and Discussion
3.1. Bacterial Inactivation Effect and Kinetics Model Analysis
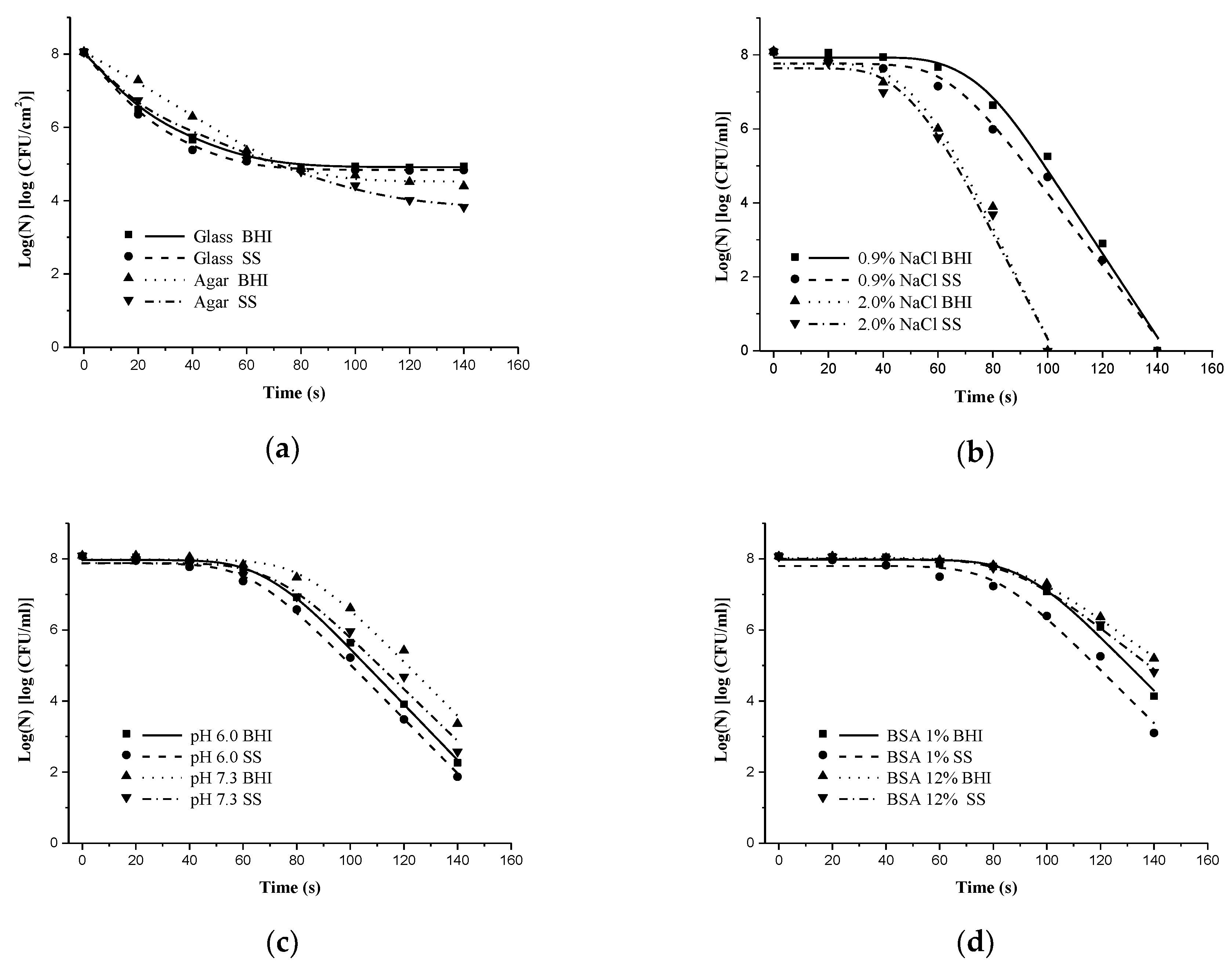
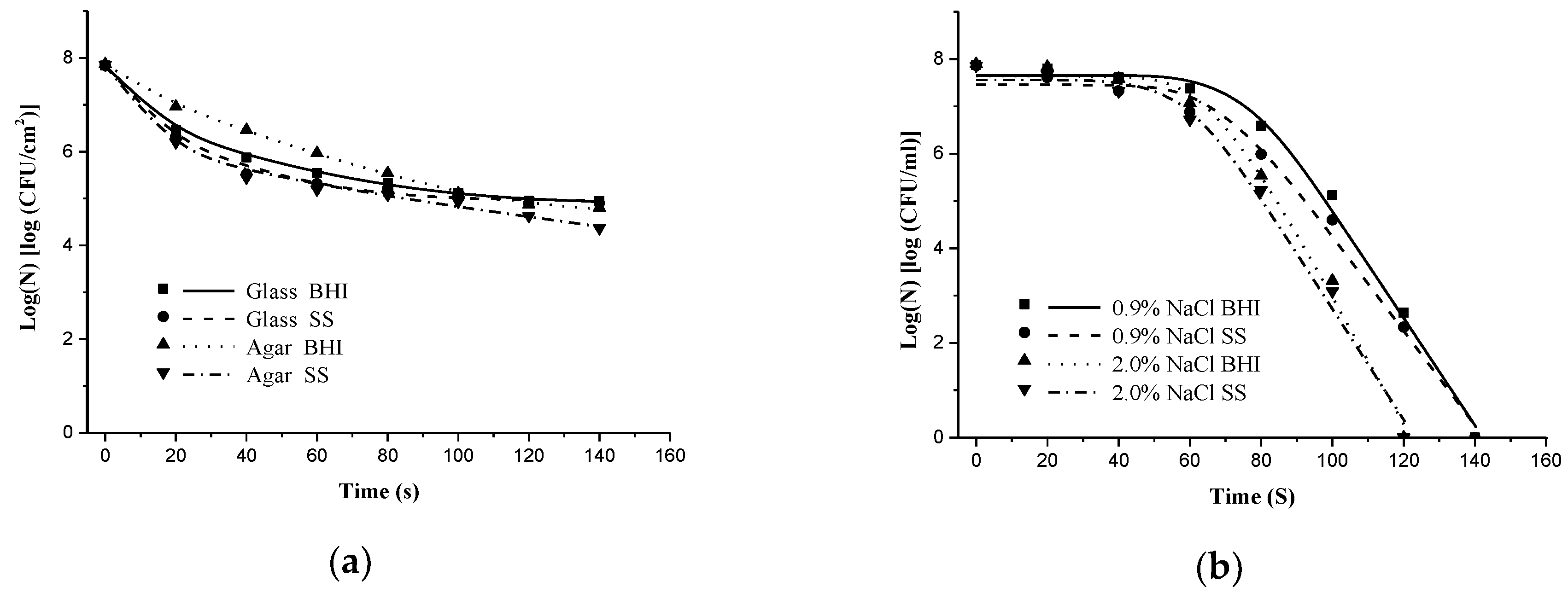
3.1.1. Effect of Solid Surface on Inactivation of S. typhimurium and S. aureus by DBD-NTP
3.1.2. Embedding S. Typhimurium and S. Aureus in Liquid Media
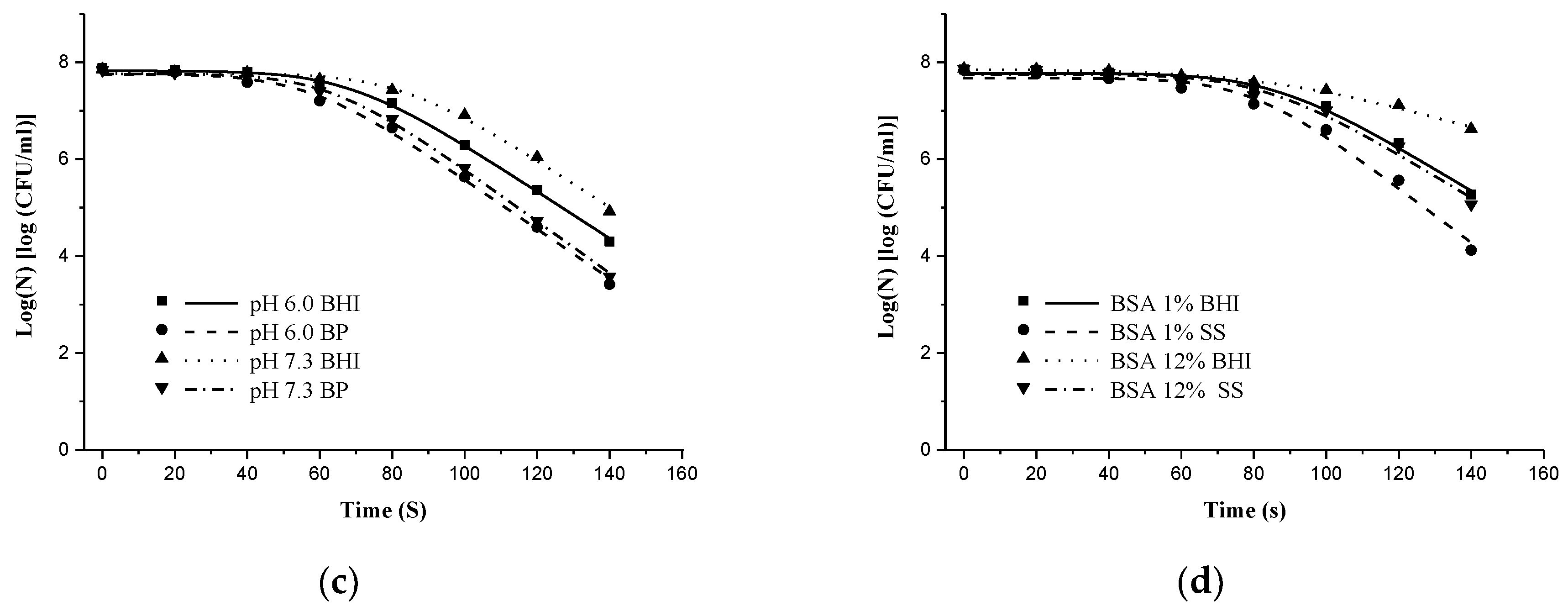
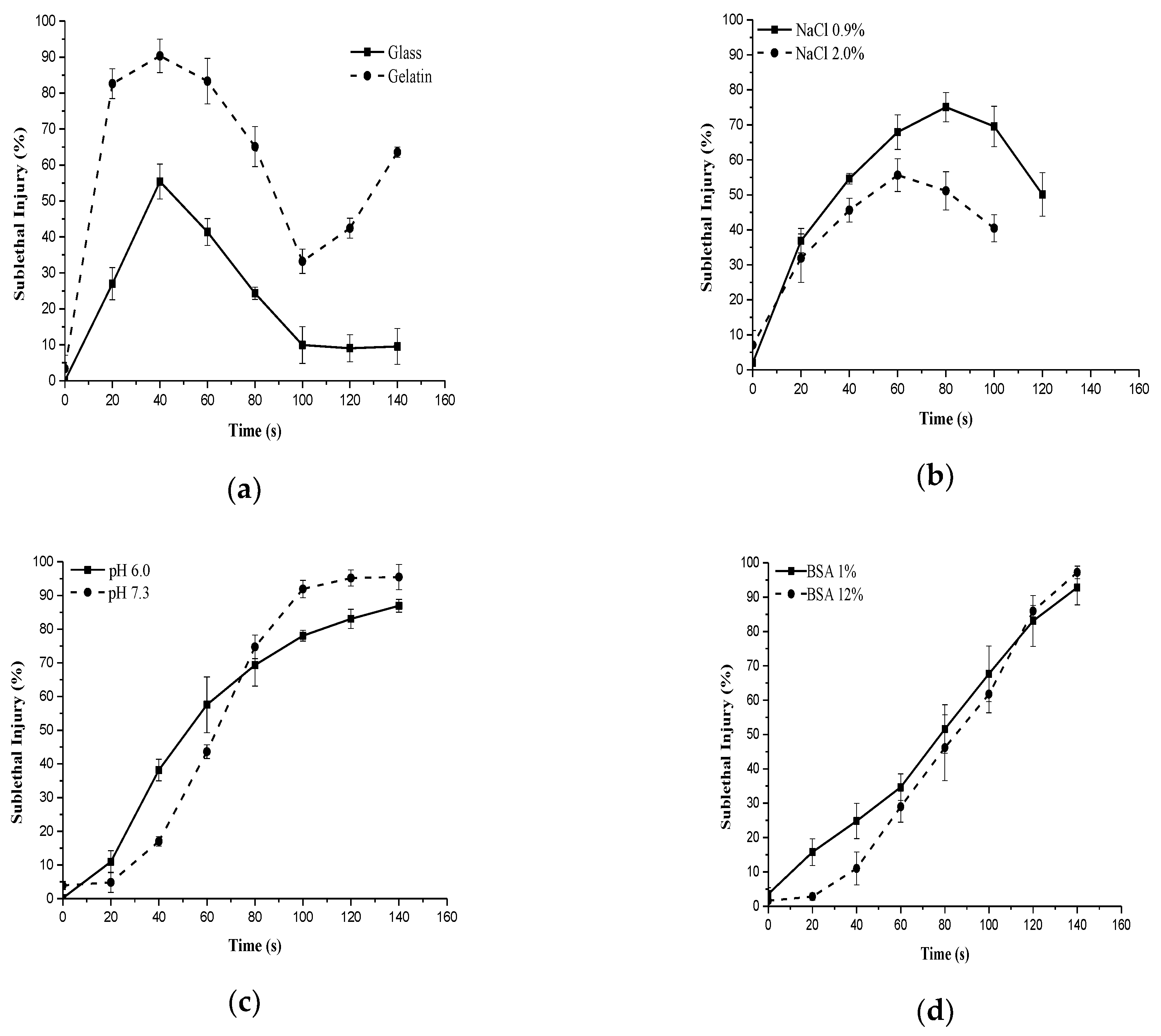
3.2. Effects of NTP on Sublethal Injury
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Surowsky, B.; Fröhling, A.; Gottschalk, N.; Schlüter, O.; Knorr, D. Impact of cold plasma on Citrobacter freundii in apple juice: Inactivation kinetics and mechanisms. Int. J. Food Microbiol. 2014, 174, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Calvo, T.; Álvarez-Ordóñez, A.; Prieto, M.; González-Raurich, M.; López, M. Influence of processing parameters and stress adaptation on the inactivation of Listeria monocytogenes by Non-Thermal Atmospheric Plasma (NTAP). Food Res. Int. 2016, 89, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Fröhling, A.; Durek, J.; Schnabel, U.; Ehlbeck, J.; Bolling, J.; Schlüter, O. Indirect plasma treatment of fresh pork: decontamination efficiency and effects on quality attributes. Innov. Food Sci. Emerg. Technol. 2012, 16, 381–390. [Google Scholar] [CrossRef]
- Hayashi, N.; Akiyoshi, Y.; Kobayashi, Y.; Kanda, K.; Ohshima, K.; Goto, M. Inactivation characteristics of Bacillus thuringiensis spore in liquid using atmospheric torch plasma using oxygen. Vacuum 2013, 88, 173–176. [Google Scholar] [CrossRef]
- Perni, S.; Shama, G.; Kong, M.G. Cold atmospheric plasma disinfection of cut fruit surfaces contaminated with migrating microorganisms. J. Food Prot. 2008, 71, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Ziuzina, D.; Patil, S.; Cullen, P.J.; Keener, K.M.; Bourke, P. Atmospheric cold plasma inactivation of Escherichia coli in liquid media inside a sealed package. J. Appl. Microbiol. 2013, 114, 778–787. [Google Scholar] [CrossRef] [PubMed]
- Rothrock, M.J.; Zhuang, H.; Lawrence, K.C.; Bowker, B.C.; Gamble, G.R.; Hiett, K.L. In-package inactivation of pathogenic and spoilage bacteria associated with poultry using dielectric barrier discharge-cold plasma treatments. Curr. Microbiol. 2017, 74, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Scholtz, V.; Pazlarova, J.; Souskova, H.; Khun, J.; Julak, J. Nonthermal plasma–A tool for decontamination and disinfection. Biotechnol. Adv. 2015, 33, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Bourke, P.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Keener, K. The potential of cold plasma for safe and sustainable food production. Trends Biotechnol. 2018, 36, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Kayes, M.M.; Critzer, F.J.; Kelly-Wintenberg, K.; Roth, J.R.; Montie, T.C.; Golden, D.A. Inactivation of foodborne pathogens using a one atmosphere uniform glow discharge plasma. Foodborne Pathog. Dis. 2007, 4, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Smet, C.; Noriega, E.; Rosier, F.; Walsh, J.L.; Valdramidis, V.P.; Van Impe, J.F. Influence of food intrinsic factors on the inactivation efficacy of cold atmospheric plasma: Impact of osmotic stress, suboptimal pH and food structure. Innov. Food Sci. Emerg. 2016, 38, 393–406. [Google Scholar] [CrossRef]
- Lee, K.; Paek, K.H.; Ju, W.T.; Lee, Y. Sterilization of bacteria, yeast, and bacterial endospores by atmospheric-pressure cold plasma using helium and oxygen. J. Microbiol. 2006, 44, 269–275. [Google Scholar] [PubMed]
- Hertwig, C.; Reineke, K.; Ehlbeck, J.; Knorr, D.; Schlüter, O. Decontamination of whole black pepper using different cold atmospheric pressure plasma applications. Food Control. 2015, 55, 221–229. [Google Scholar] [CrossRef]
- Hurst, A. Bacterial injury: A review. Can. J. Microbiol. 1977, 23, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Puligundla, P.; Lee, T.; Mok, C. Inactivation effect of dielectric barrier discharge plasma against foodborne pathogens on the surfaces of different packaging materials. Innov. Food Sci. Emerg. Technol. 2016, 36, 221–227. [Google Scholar] [CrossRef]
- Jahid, I.K.; Han, N.; Ha, S.-D. Inactivation kinetics of cold oxygen plasma depend on incubation conditions of Aeromonas hydrophila biofilm on lettuce. Food Res. Int. 2014, 55, 181–189. [Google Scholar] [CrossRef]
- Noriega, E.; Velliou, E.G.; Van Derlinden, E.; Mertens, L.; Van Impe, J.F.M. Role of growth morphology in the formulation of NaCl-based selective media for injury detection of Escherichia coli, Salmonella Typhimurium and Listeria innocua. Food Res. Int. 2014, 64, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Geeraerd, A.; Herremans, C.; Van Impe, J.F. Structural model requirements to describe microbial inactivation during a mild heat treatment. Int. J. Food. Microbiol. 2000, 59, 185–209. [Google Scholar] [CrossRef]
- Busch, S.V.; Donnelly, C.W. Development of a repair-enrichment broth for resuscitation of heat-injured Listeria monocytogenes and Listeria innocua. Appl. Environ. Microb. 1992, 58, 14–20. [Google Scholar]
- Moisan, M.; Barbeau, J.; Crevier, M.-C.; Pelletier, J.; Philip, N.; Saoudi, B. Plasma sterilization. Methods and mechanisms. Pure Appl. Chem. 2002, 74, 349–358. [Google Scholar] [CrossRef]
- Klockow, P.A.; Keener, K.M. Safety and quality assessment of packaged spinach treated with a novel ozone-generation system. LWT–Food Sci. Technol. 2009, 42, 1047–1053. [Google Scholar] [CrossRef]
- Kogelschatz, U. Dielectric-Barrier Discharges: Their history, discharge physics, and industrial applications. Plasma Chem. Plasma P. 2003, 23, 1–46. [Google Scholar] [CrossRef]
- Noriega, E.; Shama, G.; Laca, A.; Diaz, M.; Kong, M.G. Cold atmospheric gas plasma disinfection of chicken meat and chicken skin contaminated with Listeria innocua. Food Microbiol. 2011, 28, 1293–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroussi, M.; Leipold, F. Evaluation of the roles of reactive species, heat, and UV radiation in the inactivation of bacterial cells by air plasmas at atmospheric pressure. Int. J. Mass Spectrom. 2004, 233, 81–86. [Google Scholar] [CrossRef]
- Mendis, D.A.; Rosenberg, M.; Azam, F. A note on the possible electrostatic disruption of bacteria. IEEE Trans. Plasma Sci. 2000, 28, 1304–1306. [Google Scholar] [CrossRef]
- Laroussi, M.; Mendis, D.A.; Rosenberg, M. Plasma interaction with microbes. New J. Phys. 2003, 5, 41.1–41.10. [Google Scholar] [CrossRef]
- Dobrynin, D.; Fridman, G.; Friedman, G.; Fridman, A. Physical and biological mechanisms of direct plasma interaction with living tissue. New J. Phys. 2009, 11, 115020. [Google Scholar] [CrossRef] [Green Version]
- Critzer, F.J.; Kelly-Wintenberg, K.; South, S.L.; Golden, D.A. Atmospheric plasma inactivation of foodborne pathogens on fresh produce surfaces. J. Food Prot. 2007, 70, 2290–2296. [Google Scholar] [CrossRef] [PubMed]
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L.H. Low-temperature sterilization using gas plasmas: a review of the experiments and an analysis of the inactivation mechanisms. Int. J. Pharm. 2001, 226, 1–21. [Google Scholar] [CrossRef]
- Kim, H.-S.; Wright, K.; Piccioni, J.; Cho, D.J.; Cho, Y.I. Inactivation of bacteria by the application of spark plasma in produced water. Sep. Purif. Technol. 2015, 156, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Maheux, S.; Frache, G.; Thomann, J.S.; Clément, F.; Penny, C.; Belmonte, T.; Duday, D. Small unilamellar liposomes as a membrane model for cell inactivation by cold atmospheric plasma treatment. J. Phys. D Appl. Phys. 2016, 49, 344001. [Google Scholar] [CrossRef]
- Zhou, R.W.; Zhou, R.S.; Zhuang, J.X.; Zong, Z.C.; Zhang, X.H.; Liu, D.P.; Bazaka, K.; Ostrikov, K. Interaction of atmospheric-pressure air microplasmas with amino acids as fundamental processes in aqueous solution. PLoS ONE 2016, 11, e0155584. [Google Scholar] [CrossRef] [PubMed]
- Oehmigen, K.; Hähnel, M.; Brandenburg, R.; Wilke, C.; Weltmann, K.D.; von Woedtke, T. The role of acidifcation for antimicrobial activity of atmospheric pressure plasma in liquid. Plasma Process. Polym. 2010, 7, 250–257. [Google Scholar] [CrossRef]
- Stoffels, E.; Sakiyama, Y.; Graves, D.B. Cold atmospheric plasma: Charged species and their interactions with cells and tissues. IEEE T. Plasma Sci. 2008, 36, 1441–1457. [Google Scholar] [CrossRef]
- Scholtz, V.; Pazlarová, J.; Soušková, H.; Khun, J.; Julák, J. Nonthermal plasma—a tool for decontamination and disinfection. Biotechnol. Adv. 2015, 33, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Rashmei, Z.; Bornasi, H.; Ghoranneviss, M. Evaluation of treatment and disinfection of water using cold atmospheric plasma. J. Water Health 2016, 14, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Arjunan, K.P.; Obrusnik, A.; Jones, B.T.; Zajickova, L.; Ptasinska, S. Effect of additive oxygen on the reactive species profile and microbicidal property of a helium atmospheric pressure plasma jet. Plasma Process. Polym. 2016, 13, 1089–1105. [Google Scholar] [CrossRef]
- Fernández, A.; Thompson, A. The inactivation of Salmonella by cold atmospheric plasma treatment. Food Res. Int. 2012, 45, 678–684. [Google Scholar] [CrossRef]
- Patange, A.; Boehm, D.; Giltrap, M.; Lu, P.; Cullen, P.J.; Bourke, P. Assessment of the disinfection capacity and eco-toxicological impact of atmospheric cold plasma for treatment of food industry effluents. Sci. Total Environ. 2018, 631–632, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Wesche, A.M.; Gurtler, J.B.; Marks, B.P.; Ryser, E.T. Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. J. Food Prot. 2009, 72, 1121–1138. [Google Scholar] [CrossRef] [PubMed]
- Noriega, E.; Velliou, E.; Van Derlinden, E.; Mertens, L.; Van Impe, J.F. Effect of cell immobilization on heat-induced sublethal injury of Escherichia coli, Salmonella Typhimurium and Listeria innocua. Food Microbiol. 2013, 36, 355–364. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | log N0 (log (CFU/mL))/log N0 (log (CFU/cm2)) | Kmax | tl | log Nres (log (CFU/mL))/log Nres (log (CFU/cm2)) | D value 1 | Log Reduction 2 | RMSE 3 | R 2 | |
|---|---|---|---|---|---|---|---|---|---|
| NaC l0.9% | Total | 7.91 ± 0.10 aAB | 0.26 ± 0.011 aB | 73.3 ± 4.08 aC | 81.71 ± 3.72 aC | 8.07 ± 0.10 aA | 0.4002 | 0.9895 | |
| Uninjured | 7.76 ± 0.15 bBC | 0.22 ± 0.007 bB | 63.8 ± 1.08 bC | 73.6 ± 0.78 bD | 8.09 ± 0.07 aA | 0.4319 | 0.9876 | ||
| NaC l2% | Total | 7.76 ± 0.15 aB | 0.34 ± 0.011 aA | 50.2 ± 1.15 aD | 56.62 ± 0.94 aD | 8.09 ± 0.07 aA | 0.6705 | 0.9815 | |
| Uninjured | 7.64 ± 0.16 aC | 0.32 ± 0.015 aA | 48.2 ± 1.93 aD | 55.02±1.62 aE | 8.08 ± 0.09 aA | 0.7184 | 0.9783 | ||
| PBS pH = 6.0 | Total | 7.93 ± 0.14 aAB | 0.21 ± 0.041 aC | 72.5 ± 1.42 aC | 83.2 ± 1.63 aC | 6.57 ± 1.23 aB | 0.1618 | 0.9968 | |
| Uninjured | 7.82 ± 0.15 aABC | 0.20 ± 0.031 aBC | 67.8 ± 3.31 aBC | 78.84 ± 2.97 aCD | 6.83 ± 0.99 aB | 0.2185 | 0.9948 | ||
| PBS pH = 7.3 | Total | 7.97 ± 0.12 aA | 0.17 ± 0.025 aD | 82.5 ± 4.18 aB | 95.25 ± 5.41 aB | 4.72 ± 0.88 aC | 0.2623 | 0.9863 | |
| Uninjured | 7.88 ± 0.14 aAB | 0.16 ± 0.007 aDE | 71.2 ± 2.85 bB | 84.34 ± 2.37 bC | 5.50 ± 0.03 aC | 0.3250 | 0.9842 | ||
| BSA 1% | Total | 7.97 ± 0.07 aA | 0.17 ± 0.004 aD | 91.4 ± 3.47 aA | 104.03 ± 3.48 aA | 3.93 ± 0.24 aCD | 0.2126 | 0.9867 | |
| Uninjured | 7.78 ± 0.19 aBC | 0.17 ± 0.033 aCD | 81.7 ± 4.25 bA | 94.49 ± 1.72 bB | 4.95 ± 0.74 aCD | 0.3515 | 0.9761 | ||
| BSA 12% | Total | 8.02 ± 0.10 aA | 0.12 ± 0.009 aE | 88.3 ± 2.56 aA | 106.03 ± 1.73 aA | 2.87 ± 0.20 aE | 0.0620 | 0.9980 | |
| Uninjured | 7.99 ± 0.08 aAB | 0.13 ± 0.020 aEF | 87.6 ± 5.81 aA | 104.01 ± 7.28 aA | 3.24 ± 0.71 aE | 0.0961 | 0.9961 | ||
| Glass | Total | 8.05 ± 0.07 bA | 0.09 ± 0.007 bF | 4.91 ± 0.61 aA | 9.11 ± 3.37 aF | 3.11 ± 0.58 aDE | 0.0432 | 0.9991 | |
| Uninjured | 8.04 ± 0.07 aA | 0.10 ± 0.001 aF | 4.83 ± 0.59 aA | 8.11 ± 3.35 aF | 3.20 ± 0.56 aE | 0.0665 | 0.9980 | ||
| Gelatin | Total | 8.06 ± 0.07 aA | 0.10 ± 0.005 aEF | 4.52 ± 0.15 aA | 23.66 ± 2.72 aE | 3.66 ± 0.02 bDE | 0.1083 | 0.9964 | |
| Uninjured | 8.06 ± 0.07 bA | 0.06 ± 0.001 bG | 3.83 ± 0.18 bB | 9.02 ± 1.68 bF | 4.23 ± 0.08 aD | 0.1629 | 0.9928 |
| Population | log N0 (log (CFU/mL))/log N0 (log (CFU/cm2)) | Kmax | tl | log Nres (log (CFU/mL))/log Nres (log (CFU/cm2)) | D value 1 | Log Reduction 2 | RMSE 3 | R 2 | |
|---|---|---|---|---|---|---|---|---|---|
| NaCl0.9% | Total | 7.65 ± 0.1 aA | 0.26 ± 0.015 aB | 74.57 ± 2.78 aC | 82.97 ± 2.29 aC | 7.87 ± 0.12 aA | 0.3392 | 0.9921 | |
| Uninjured | 7.46 ± 0.09 aC | 0.23 ± 0.013 bB | 67.27 ± 3.17 bB | 76.87 ± 2.61 bCD | 7.86 ± 0.11 aA | 0.4218 | 0.9874 | ||
| NaCl2% | Total | 7.65 ± 0.12 aA | 0.31 ± 0.021 aA | 64.44 ± 2.97 aD | 71.61 ± 2.48 aD | 7.89 ± 0.14 aA | 0.4283 | 0.9896 | |
| Uninjured | 7.55 ± 0.1 aBC | 0.27 ± 0.015 aA | 59.01 ± 2.86 aCD | 67.07 ± 2.41 aE | 7.86 ± 0.11 aA | 0.4726 | 0.9869 | ||
| PBS pH=6.0 | Total | 7.82 ± 0.06 aA | 0.11 ± 0.002 aC | 68.26 ± 0.81 aD | 87.98 ± 0.74 aC | 3.57 ± 0.06 bB | 0.069 | 0.9985 | |
| Uninjured | 7.78 ± 0.1 aA | 0.12 ± 0.007 aC | 55.94 ± 0.82 bD | 74.28 ± 1.31 bD | 4.46 ± 0.05 aB | 0.1464 | 0.9955 | ||
| PBS pH=7.3 | Total | 7.78 ± 0.11 aA | 0.11 ± 0.013 aC | 82.47 ± 5.77 aB | 102.28 ± 5.06 aB | 2.94 ± 0.34 bCD | 0.1012 | 0.9948 | |
| Uninjured | 7.76 ± 0.1 aA | 0.12 ± 0.007 aC | 62.66 ± 2.49 bBC | 80.54 ± 1.56 bC | 4.26 ± 0.08 aB | 0.089 | 0.9982 | ||
| BSA 1% | Total | 7.77 ± 0.11 aA | 0.1 ± 0.004 bC | 86.22 ± 1.16 aAB | 107.39 ± 2.05 aB | 2.59 ± 0.16 bD | 0.1176 | 0.9907 | |
| Uninjured | 7.67 ± 0.12 aAB | 0.13 ± 0.009 aC | 79.66 ± 4.62 aA | 96.62 ± 4.2 bB | 3.71 ± 0.3 aCD | 0.2096 | 0.9854 | ||
| BSA 12% | Total | 7.85 ± 0.14 aA | 0.05 ± 0.017 bD | 88.86 ± 1.51 aA | 134.57 ± 15.59 aA | 1.25 ± 0.43 bE | 0.0485 | 0.993 | |
| Uninjured | 7.75 ± 0.1 aA | 0.1 ± 0.002 aC | 83.48 ± 5.41 aA | 104.6 ± 5.05 bA | 2.8 ± 0.24 aD | 0.1838 | 0.9799 | ||
| Glass | Total | 7.84 ± 0.12 aA | 0.04 ± 0.011 aD | 4.88 ± 0.09 aA | 9.14 ± 2.14 aF | 2.9 ± 0.1 aCD | 0.0464 | 0.9987 | |
| Uninjured | 7.84 ± 0.09 aA | 0.06 ± 0.035 aD | 4.95 ± 0.05 aA | 7.44 ± 4.18 aF | 2.94 ± 0.14 aD | 0.1196 | 0.9919 | ||
| Gelatin | Total | 7.86 ± 0.08 aA | 0.06 ± 0.011 aD | 4.7 ± 0.09 aB | 25.47 ± 6.38 aE | 3.06 ± 0.06 aC | 0.0393 | 0.9993 | |
| Uninjured | 7.85 ± 0.08 aA | 0.02 ± 0.003 bE | 3.76 ± 0.7 aB | 3.44 ± 0.85 bF | 3.48 ± 0.07 aC | 0.1465 | 0.9901 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, M.; Zhuang, H.; Wang, J.; Yan, W.; Zhao, J.; Zhang, J. Inactivation Kinetics of Salmonella typhimurium and Staphylococcus aureus in Different Media by Dielectric Barrier Discharge Non-Thermal Plasma. Appl. Sci. 2018, 8, 2087. https://doi.org/10.3390/app8112087
Huang M, Zhuang H, Wang J, Yan W, Zhao J, Zhang J. Inactivation Kinetics of Salmonella typhimurium and Staphylococcus aureus in Different Media by Dielectric Barrier Discharge Non-Thermal Plasma. Applied Sciences. 2018; 8(11):2087. https://doi.org/10.3390/app8112087
Chicago/Turabian StyleHuang, Mingming, Hong Zhuang, Jiamei Wang, Wenjing Yan, Jianying Zhao, and Jianhao Zhang. 2018. "Inactivation Kinetics of Salmonella typhimurium and Staphylococcus aureus in Different Media by Dielectric Barrier Discharge Non-Thermal Plasma" Applied Sciences 8, no. 11: 2087. https://doi.org/10.3390/app8112087
APA StyleHuang, M., Zhuang, H., Wang, J., Yan, W., Zhao, J., & Zhang, J. (2018). Inactivation Kinetics of Salmonella typhimurium and Staphylococcus aureus in Different Media by Dielectric Barrier Discharge Non-Thermal Plasma. Applied Sciences, 8(11), 2087. https://doi.org/10.3390/app8112087