
Activity Strength within Optic Flow-Sensitive Cortical Regions Is Associated with Visual Path Integration Accuracy in Aged Adults


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cognitive Assessments
2.3. Functional MRI Paradigms
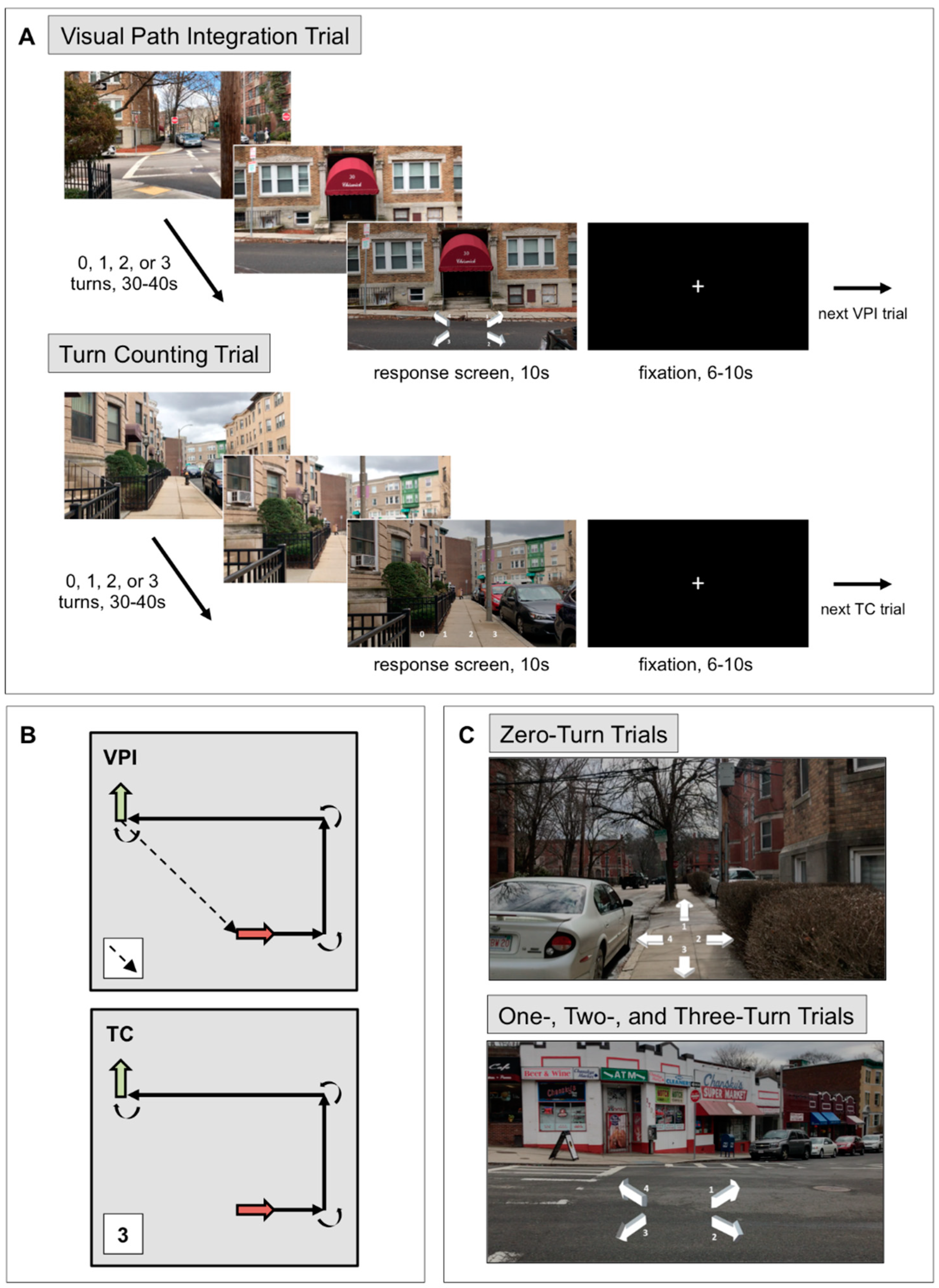
2.3.1. Visual Path Integration Task
2.3.2. Turn Counting Task
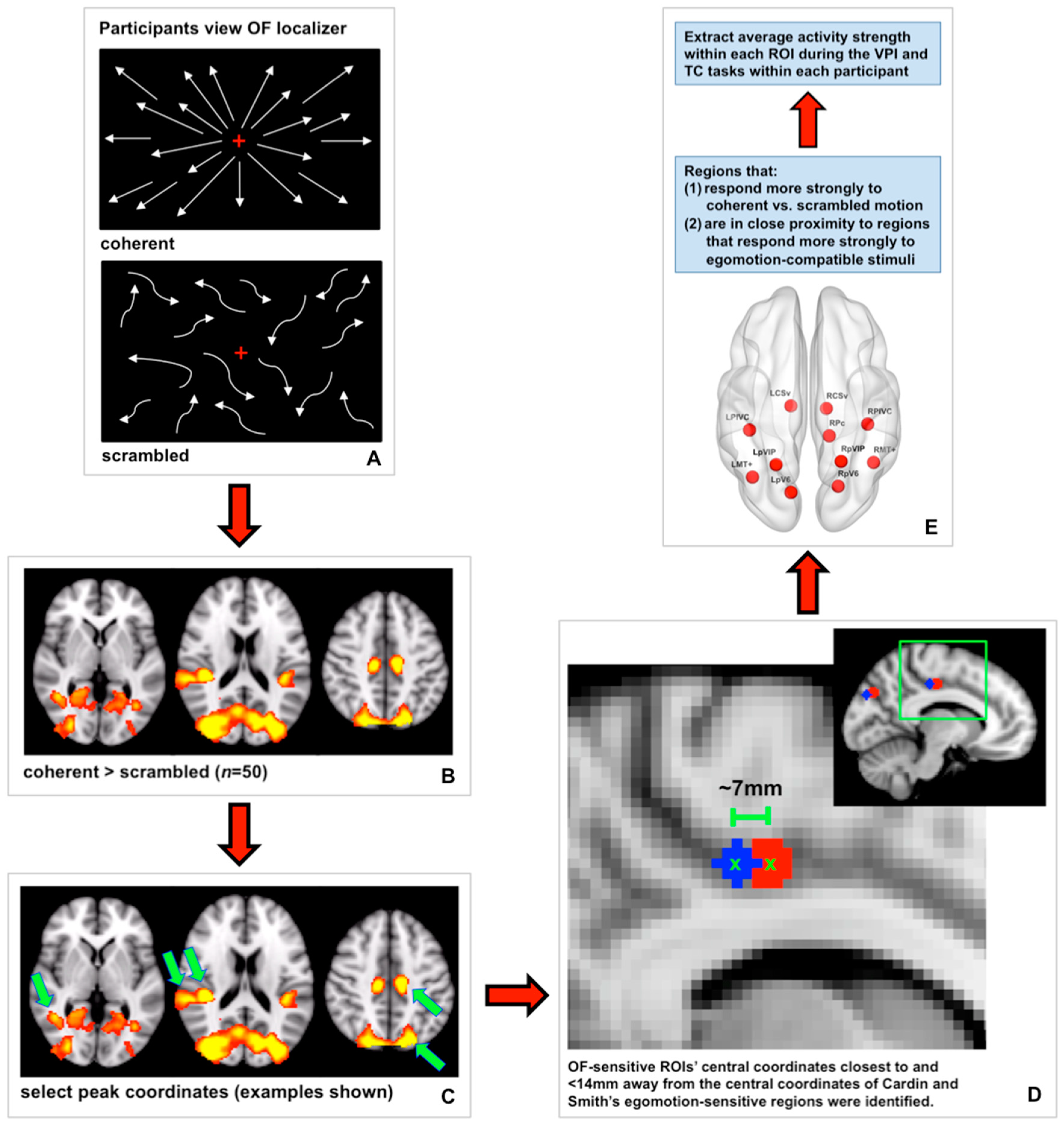
2.3.3. Optic Flow Localizer
2.4. Magnetic Resonance Imaging
2.5. Global Motion Coherence Thresholds
2.6. Image Processing
2.6.1. Preprocessing
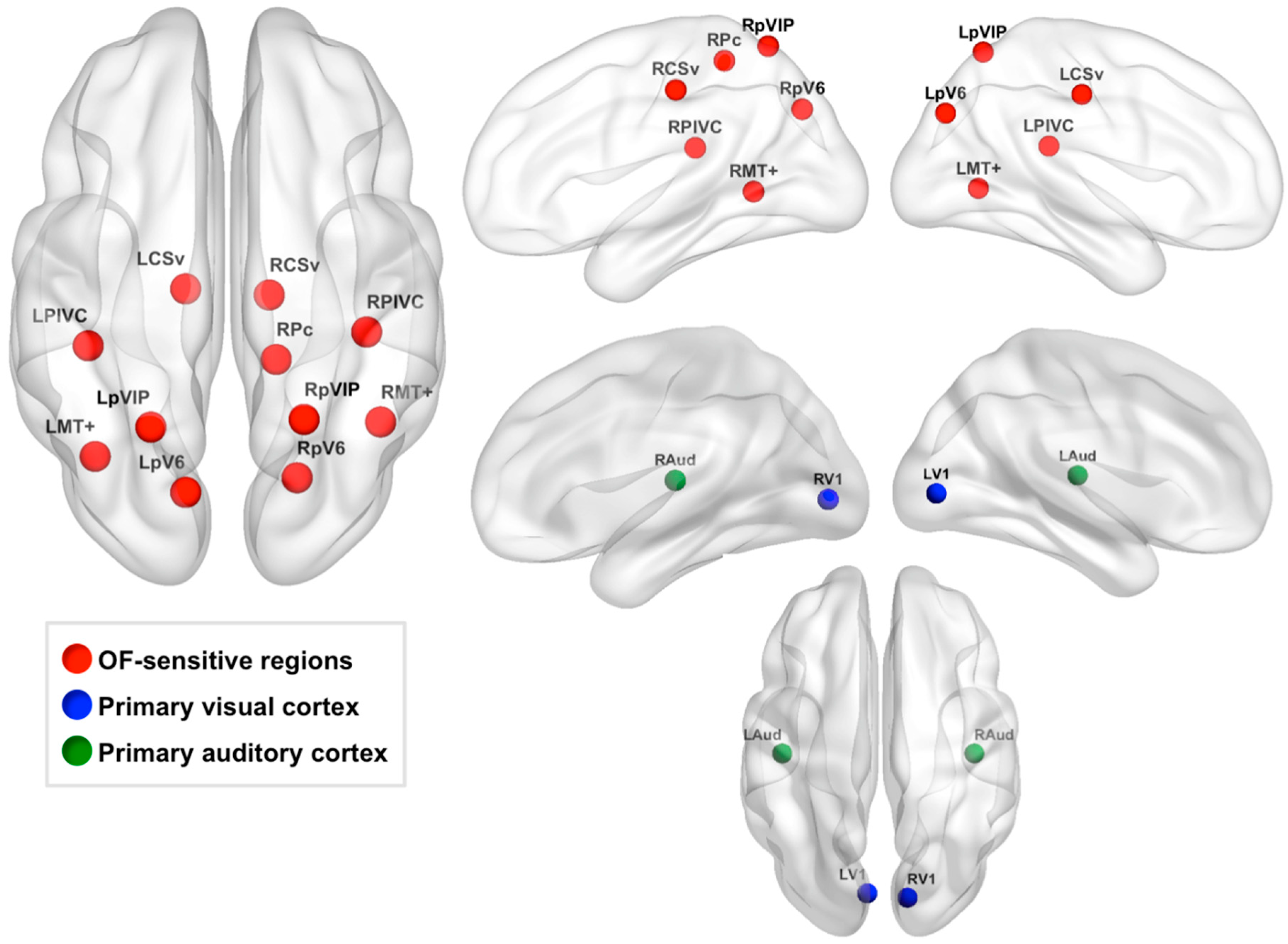
2.6.2. Defining OF-Sensitive ROIs
2.6.3. Defining Control ROIs
2.6.4. Measuring Brain Activity in ROIs during the VPI and TC Tasks
2.7. Effect of Age on VPI and TC Task Performance
2.8. Assessing the Effects of Age and Performance on OF-Sensitive Region Activity
3. Results
3.1. Effect of Age on VPI and TC Task Performance
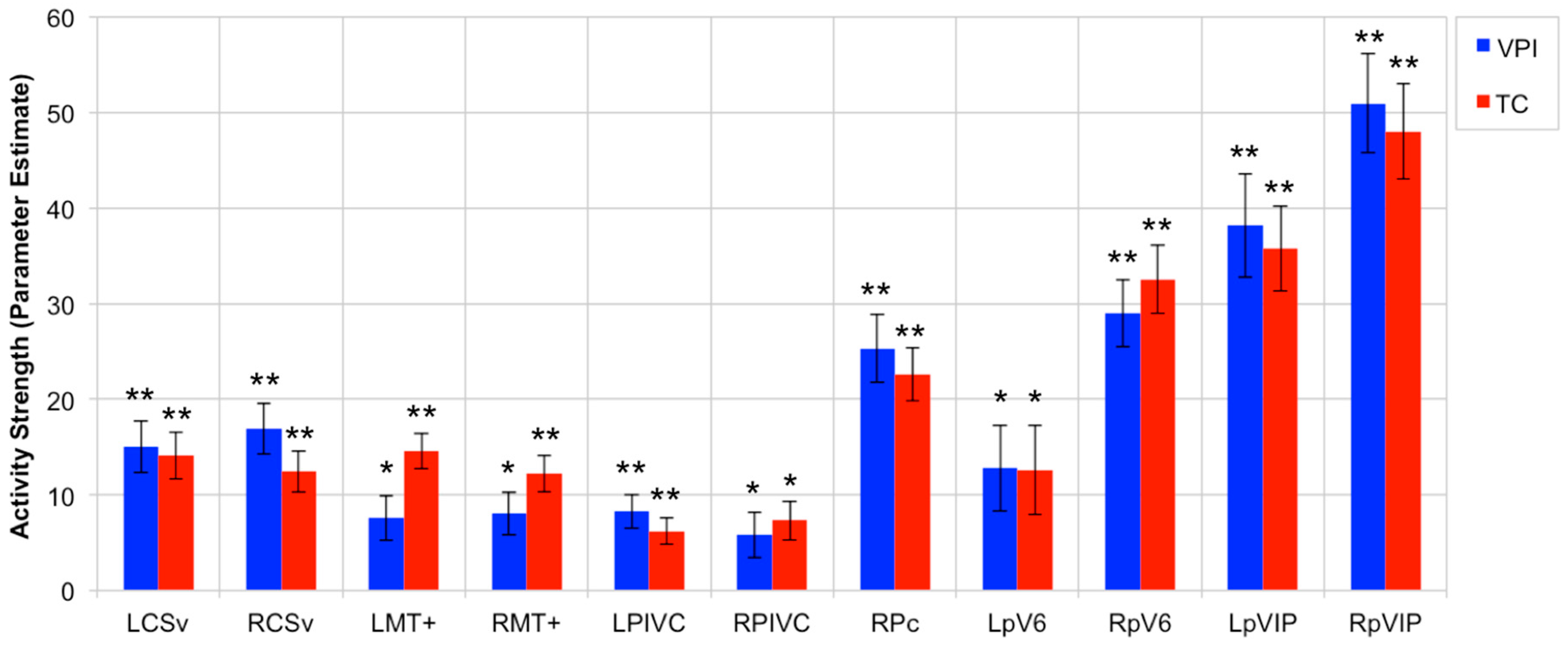
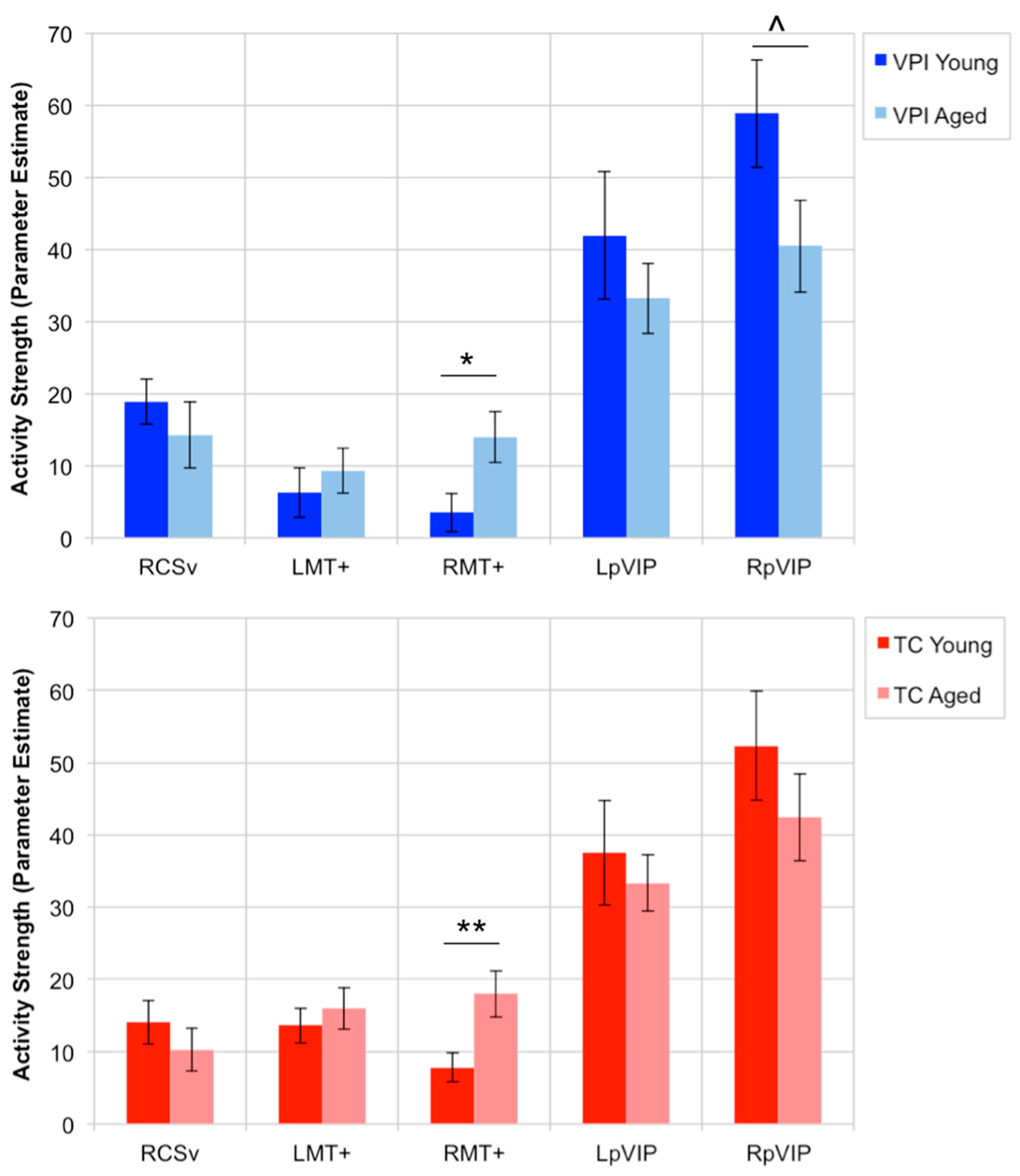
3.2. OF-Sensitive Region Activity Strength during VPI and TC Tasks
3.3. Effect of Age on OF-Sensitive Region Activity during VPI and TC
3.4. Relationship between VPI Accuracy and OF-Sensitive Region Activity Strength
4. Discussion
4.1. Activity in RCSv, LMT+, RMT+, LpVIP, and RpVIP during VPI Was Inversely Related to Global Radial Motion Thresholds
4.2. Stronger Activity in RMT+ in Aged Adults during VPI and TC
4.3. Activity Strength in LMT+, LpVIP, and RpVIP Is Associated with VPI Accuracy
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lester, A.W.; Moffat, S.D.; Wiener, J.M.; Barnes, C.A.; Wolbers, T. The aging navigational system. Neuron 2017, 95, 1019–1035. [Google Scholar] [CrossRef]
- Chrastil, E.R. Neural evidence supports a novel framework for spatial navigation. Psychon. Bull. Rev. 2013, 20, 208–227. [Google Scholar] [CrossRef]
- Marchette, S.A.; Bakker, A.; Shelton, A.L. Cognitive mappers to creatures of habit: Differential engagement of place and response learning mechanisms predicts human navigational behavior. J. Neurosci. 2011, 31, 15264–15268. [Google Scholar] [CrossRef] [PubMed]
- Wolbers, T.; Hegarty, M. What determines our navigational abilities? Trends Cogn. Sci. 2010, 14, 138–146. [Google Scholar] [CrossRef]
- Warren, W.H. Optic flow. In The Visual Neurosciences; Chalupa, L., Werner, J., Eds.; MIT Press: Cambridge, MA, USA, 2004; Volume 2, pp. 1247–1259. [Google Scholar]
- Antal, A.; Baudewig, J.; Paulus, W.; Dechent, P. The posterior cingulate cortex and planum temporale/parietal operculum are activated by coherent visual motion. Vis. Neurosci. 2008, 25, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braddick, O.J.; O’Brien, J.M.D.; Wattam-Bell, J.; Atkinson, J.; Hartley, T.; Turner, R. Brain areas sensitive to coherent visual motion. Perception 2001, 30, 61–72. [Google Scholar] [CrossRef]
- Cardin, V.; Smith, A.T. Sensitivity of human visual and vestibular cortical regions to egomotion-compatible visual stimulation. Cereb. Cortex 2009, 20, 1964–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitzalis, S.; Sdoia, S.; Bultrini, A.; Committeri, G.; Di Russo, F.; Fattori, P.; Galletti, C.; Galati, G. Selectivity to translational egomotion in human brain motion areas. PLoS ONE 2013, 8, e60241. [Google Scholar] [CrossRef] [Green Version]
- Cottereau, B.R.; Smith, A.T.; Rima, S.; Fize, D.; Héjja-Brichard, Y.; Renaud, L.; Lejards, C.; Vayssiere, N.; Trotter, Y.; Durand, J.-B. Processing of egomotion-consistent optic flow in the Rhesus Macaque Cortex. Cereb. Cortex 2017, 27, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Orban, G.A.; Fize, D.; Peuskens, H.; Denys, K.; Nelissen, K.; Sunaert, S.; Todd, J.; Vanduffel, W. Similarities and differences in motion processing between the human and macaque brain: Evidence from fMRI. Neuropsychologia 2003, 41, 1757–1768. [Google Scholar] [CrossRef]
- DeAngelis, G.C.; Angelaki, D.E. Visual-vestibular integration for self-motion perception. In The Neural Bases of Multisensory Processes; Murray, M.M., Wallace, M.T., Eds.; Taylor & Francis Group, LLC.: Boca Raton, FL, USA, 2012. [Google Scholar]
- Greenlee, M.W.; Frank, S.M.; Kaliuzhna, M.; Blanke, O.; Bremmer, F.; Churan, J.; Cuturi, L.F.; MacNeilage, P.R.; Smith, A.T. Multisensory integration in self motion perception. Multisens. Res. 2016, 29, 525–556. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Greenlee, M.W.; DeAngelis, G.C.; Angelaki, D.E. Distributed visual–vestibular processing in the cerebral cortex of man and macaque. Multisens. Res. 2017, 30, 91–120. [Google Scholar] [CrossRef] [Green Version]
- Bremmer, F.; Schlack, A.; Shah, N.J.; Zafiris, O.; Kubischik, M.; Hoffmann, K.; Fink, G.R. Polymodal motion processing in posterior parietal and premotor cortex: A human fMRI study strongly implies equivalencies between humans and monkeys. Neuron 2001, 29, 287–296. [Google Scholar] [CrossRef]
- Dukelow, S.P.; DeSouza, J.F.X.; Culham, J.C.; van den Berg, A.V.; Menon, R.S.; Vilis, T. Distinguishing subregions of the human MT complex using visual fields and pursuit eye movements. J. Neurophysiol. 2001, 86, 1991–2000. [Google Scholar] [CrossRef]
- Sereno, M.I.; Huang, R.-S. A human parietal face area contains aligned head-centered visual and tactile maps. Nat. Neurosci. 2006, 9, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.T.; Wall, M.B.; Williams, A.L.; Singh, K.D. Sensitivity to optic flow in human cortical areas MT and MST. Eur. J. Neurosci. 2006, 23, 561–569. [Google Scholar] [CrossRef]
- Wall, M.B.; Smith, A. The representation of egomotion in the human brain. Curr. Biol. 2008, 18, 191–194. [Google Scholar] [CrossRef] [Green Version]
- Cardin, V.; Hemsworth, L.; Smith, A. Adaptation to heading direction dissociates the roles of human MST and V6 in the processing of optic flow. J. Neurophysiol. 2012, 108, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Furlan, M.; Wann, J.P.; Smith, A. A Representation of changing heading direction in human cortical areas pVIP and CSv. Cereb. Cortex 2013, 24, 2848–2858. [Google Scholar] [CrossRef] [Green Version]
- Peuskens, H.; Sunaert, S.; Dupont, P.; Van Hecke, P.; Orban, G.A. Human brain regions involved in heading estimation. J. Neurosci. 2001, 21, 2451–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wutte, M.G.; Smith, M.T.; Flanagin, V.; Wolbers, T. Physiological signal variability in hMT+ reflects performance on a direction discrimination task. Front. Psychol. 2011, 2, 185. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Beer, A.L.; Furlan, M.; Mars, R.B. Connectivity of the cingulate sulcus visual area (CSv) in the human cerebral cortex. Cereb. Cortex 2017, 28, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutmore, T.R.; Hine, T.J.; Maberly, K.J.; Langford, N.M.; Hawgood, G. Cognitive and gender factors influencing navigation in a virtual environment. Int. J. Hum. Comput. Stud. 2000, 53, 223–249. [Google Scholar] [CrossRef]
- Kearns, M.J.; Warren, W.H.; Duchon, A.P.; Tarr, M.J. Path integration from optic flow and body senses in a homing task. Perception 2002, 31, 349–374. [Google Scholar] [CrossRef] [PubMed]
- Kirschen, M.P.; Kahana, M.J.; Sekuler, R.; Burack, B. Optic Flow Helps Humans Learn to Navigate through Synthetic Environments. Perception 2000, 29, 801–818. [Google Scholar] [CrossRef]
- Tan, D.S.; Czerwinski, M.; Robertson, G. Women go with the (optical) flow. In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Ft. Lauderdale, FL, USA, 5–10 April 2003; pp. 209–215. [Google Scholar]
- Tan, D.S.; Czerwinski, M.P.; Robertson, G.G. Large displays enhance optical flow cues and narrow the gender gap in 3-D virtual navigation. Hum. Factors J. Hum. Factors Ergon. Soc. 2006, 48, 318–333. [Google Scholar] [CrossRef] [Green Version]
- Tcheang, L.; Bülthoff, H.H.; Burgess, N. Visual influence on path integration in darkness indicates a multimodal representation of large-scale space. Proc. Natl. Acad. Sci. USA 2011, 108, 1152–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavcic, V.; Fernandez, R.; Logan, D.; Duffy, C.J. Neurophysiological and perceptual correlates of navigational impairment in Alzheimer’s disease. Brain 2006, 129, 736–746. [Google Scholar] [CrossRef] [Green Version]
- Mapstone, M.; Dickerson, K.; Duffy, C.J. Distinct mechanisms of impairment in cognitive ageing and Alzheimer’s disease. Brain 2008, 131, 1618–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, H.L.; Tetewsky, S.J.; Avery, L.M.; Cushman, L.A.; Makous, W.; Duffy, C.J. Visual mechanisms of spatial disorientation in Alzheimer’s disease. Cereb. Cortex 2001, 11, 1083–1092. [Google Scholar] [CrossRef] [Green Version]
- Tetewsky, S.J.; Duffy, C.J. Visual loss and getting lost in Alzheimer’s disease. Neurology 1999, 52, 958. [Google Scholar] [CrossRef]
- Warren, W.H., Jr.; Blackwell, A.W.; Morris, M.W. Age differences in perceiving the direction of self-motion from optical flow. J. Gerontol. 1989, 44, P147–P153. [Google Scholar] [CrossRef] [Green Version]
- Antonova, E.; Parslow, D.; Brammer, M.; Dawson, G.R.; Jackson, S.H.D.; Morris, R.G. Age-related neural activity during allocentric spatial memory. Memory 2009, 17, 125–143. [Google Scholar] [CrossRef] [PubMed]
- Meulenbroek, O.; Petersson, K.M.; Voermans, N.; Weber, B.; Fernandez, G. Age differences in neural correlates of route encoding and route recognition. NeuroImage 2004, 22, 1503–1514. [Google Scholar] [CrossRef] [Green Version]
- Moffat, S.D.; Elkins, W.; Resnick, S.M. Age differences in the neural systems supporting human allocentric spatial navigation. Neurobiol. Aging 2006, 27, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Konishi, K.; Etchamendy, N.; Roy, S.; Marighetto, A.; Rajah, N.; Bohbot, V.D. Decreased functional magnetic resonance imaging activity in the hippocampus in favor of the caudate nucleus in older adults tested in a virtual navigation task. Hippocampus 2013, 23, 1005–1014. [Google Scholar] [CrossRef]
- Schuck, N.W.; Doeller, C.F.; Polk, T.A.; Lindenberger, U.; Li, S.-C. Human aging alters the neural computation and representation of space. NeuroImage 2015, 117, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Stangl, M.; Achtzehn, J.; Huber, K.; Dietrich, C.; Tempelmann, C.; Wolbers, T. Compromised grid-cell-like representations in old age as a key mechanism to explain age-related navigational deficits. Curr. Biol. 2018, 28, 1108–1115.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajac, L.; Burte, H.; Taylor, H.A.; Killiany, R.J. Self-reported navigation ability is associated with optic flow-sensitive regions’ functional connectivity patterns during visual path integration. Brain Behav. 2019, 9, e01236. [Google Scholar] [CrossRef] [PubMed]
- Knopman, D.S.; Roberts, R.O.; Geda, Y.E.; Pankratz, V.S.; Christianson, T.J.H.; Petersen, R.C.; Rocca, W.A. Validation of the telephone interview for cognitive status-modified in subjects with normal cognition, mild cognitive impairment, or dementia. Neuroepidemiology 2010, 34, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Nasreddine, Z.S.; Phillips, N.A.; Bedirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Luis, C.A.; Keegan, A.P.; Mullan, M. Cross validation of the Montreal Cognitive Assessment in community dwelling older adults residing in the Southeastern US. Int. J. Geriatr. Psychiatry 2009, 24, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, H.C.; Lacritz, L.H.; Cullum, C.M.; Weiner, M.F. Normative data for the Montreal Cognitive Assessment (MoCA) in a population-based sample. Neurology 2011, 77, 1272–1275. [Google Scholar] [CrossRef]
- Waldron-Perrine, B.; Axelrod, B.N. Determining an appropriate cutting score for indication of impairment on the Montreal Cognitive Assessment. Int. J. Geriatr. Psychiatry 2012, 27, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Pitzalis, S.; Sereno, M.I.; Committeri, G.; Fattori, P.; Galati, G.; Patria, F.; Galletti, C. Human V6: The medial motion area. Cereb. Cortex 2009, 20, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Pérez, M.A. Forced-choice staircases with fixed step sizes: Asymptotic and small-sample properties. Vision Res. 1998, 38, 1861–1881. [Google Scholar] [CrossRef] [Green Version]
- Jenkinson, M.; Beckmann, C.F.; Behrens, T.E.J.; Woolrich, M.W.; Smith, S.M. FSL. NeuroImage 2012, 62, 782–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salimi-Khorshidi, G.; Douaud, G.; Beckmann, C.F.; Glasser, M.F.; Griffanti, L.; Smith, S.M. Automatic denoising of functional MRI data: Combining independent component analysis and hierarchical fusion of classifiers. NeuroImage 2014, 90, 449–468. [Google Scholar] [CrossRef] [Green Version]
- Greve, D.N.; Fischl, B. Accurate and robust brain image alignment using boundary-based registration. NeuroImage 2009, 48, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S.M. Improved optimization for the robust and accurate linear registration and motion correction of brain images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef]
- Jenkinson, M.; Smith, S. A global optimisation method for robust affine registration of brain images. Med. Image Anal. 2001, 5, 143–156. [Google Scholar] [CrossRef]
- Andersson, J.L.; Jenkinson, M.; Smith, S. Non-Linear Registration aka Spatial Normalisation FMRIB Technical Report TR07JA2; FMRIB Analysis Group of the University of Oxford; FMRIB Centre: Oxford, UK, 2007. [Google Scholar]
- Xia, M.; Wang, J.; He, Y. BrainNet viewer: A network visualization tool for human brain connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischl, B.; Rajendran, N.; Busa, E.; Augustinack, J.; Hinds, O.; Yeo, B.T.; Mohlberg, H.; Amunts, K.; Zilles, K. Cortical folding patterns and predicting cytoarchitecture. Cereb. Cortex 2007, 18, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Kimmig, H.; Greenlee, M.W.; Gondan, M.; Schira, M.M.; Kassubek, J.; Mergner, T. Relationship between saccadic eye movements and cortical activity as measured by fMRI: Quantitative and qualitative aspects. Exp. Brain Res. 2001, 141, 184–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, S.J.; Killiany, R.J. Hippocampal subregions are differentially affected in the progression to Alzheimer’s disease. Anat. Rec. 2012, 295, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Hartley, T.; Maguire, E.A.; Spiers, H.J.; Burgess, N. The well-worn route and the path less traveled: Distinct neural bases of route following and wayfinding in humans. Neuron 2003, 37, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Maguire, E.A.; Gadian, D.G.; Johnsrude, I.S.; Good, C.D.; Ashburner, J.; Frackowiak, R.S.J.; Frith, C.D. Navigation-related structural change in the hippocampi of taxi drivers. Proc. Natl. Acad. Sci. USA 2000, 97, 4398–4403. [Google Scholar] [CrossRef] [Green Version]
- Maguire, E.A.; Woollett, K.; Spiers, H.J. London taxi drivers and bus drivers: A structural MRI and neuropsychological analysis. Hippocampus 2006, 16, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Iaria, G.; Chen, J.-K.; Guariglia, C.; Ptito, A.; Petrides, M. Retrosplenial and hippocampal brain regions in human navigation: Complementary functional contributions to the formation and use of cognitive maps. Eur. J. Neurosci. 2007, 25, 890–899. [Google Scholar] [CrossRef]
- Epstein, R.A. Parahippocampal and retrosplenial contributions to human spatial navigation. Trends Cogn. Sci. 2008, 12, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Epstein, R.A.; Patai, E.Z.; Julian, J.B.; Spiers, H.J. The cognitive map in humans: Spatial navigation and beyond. Nat. Neurosci. 2017, 20, 1504–1513. [Google Scholar] [CrossRef]
- Ino, T.; Doi, T.; Hirose, S.; Kimura, T.; Ito, J.; Fukuyama, H. Directional disorientation following left retrosplenial hemorrhage: A case report with FMRI studies. Cortex 2007, 43, 248–254. [Google Scholar] [CrossRef]
- Marchette, S.A.; Vass, L.K.; Ryan, J.P.; Epstein, R.A. Anchoring the neural compass: Coding of local spatial reference frames in human medial parietal lobe. Nat. Neurosci. 2014, 17, 1598–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, N.; Kawamura, M.; Shiota, J.; Kasahata, N.; Hirayama, K. Pure topographic disorientation due to right retrosplenial lesion. Neurology 1997, 49, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Vann, S.D.; Aggleton, J.P.; Maguire, E.A. What does the retrosplenial cortex do? Nat. Rev. Neurosci. 2009, 10, 792–802. [Google Scholar] [CrossRef]
- Wolbers, T.; Buchel, C. Dissociable retrosplenial and hippocampal contributions to successful formation of survey representations. J. Neurosci. 2005, 25, 3333–3340. [Google Scholar] [CrossRef] [Green Version]
- Helfrich, R.F.; Becker, H.G.; Haarmeier, T. Processing of coherent visual motion in topographically organized visual areas in human cerebral cortex. Brain Topogr. 2013, 26, 247–263. [Google Scholar] [CrossRef]
- Orban, G.; Van Essen, D.; Vanduffel, W. Comparative mapping of higher visual areas in monkeys and humans. Trends Cogn. Sci. 2004, 8, 315–324. [Google Scholar] [CrossRef]
- Vanduffel, W.; Fize, D.; Peuskens, H.; Denys, K.; Sunaert, S.; Todd, J.T.; Orban, G.A. Extracting 3D from motion: Differences in human and monkey intraparietal cortex. Science 2002, 298, 413–415. [Google Scholar] [CrossRef] [Green Version]
- Fischer, E.; Bülthoff, H.H.; Logothetis, N.K.; Bartels, A. Visual motion responses in the posterior cingulate sulcus: A comparison to V5/MT and MST. Cereb. Cortex 2011, 22, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Biehl, S.C.; Andersen, M.; Waiter, G.D.; Pilz, K.S. Neural changes related to motion processing in healthy aging. Neurobiol. Aging 2017, 57, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Yang, Y.; Li, G.; Zhang, J.; Wang, Y.; Zhou, Y.; Leventhal, A.G. Aging affects the direction selectivity of MT cells in rhesus monkeys. Neurobiol. Aging 2010, 31, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liang, Z.; Li, G.; Wang, Y.; Zhou, Y. Aging affects response variability of V1 and MT neurons in rhesus monkeys. Brain Res. 2009, 1274, 21–27. [Google Scholar] [CrossRef]
- Leventhal, A.G. GABA and its agonists improved visual cortical function in senescent monkeys. Science 2003, 300, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, M.S.; Cox, R.W.; DeYoe, E.A. Graded effects of spatial and featural attention on human area MT and associated motion processing areas. J. Neurophysiol. 1997, 78, 516–520. [Google Scholar] [CrossRef]
- O’Craven, K.M.; Rosen, B.R.; Kwong, K.K.; Treisman, A.; Savoy, R.L. Voluntary attention modulates fMRI activity in human MT–MST. Neuron 1997, 18, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Treue, S.; Maunsell, J.H.R. Attentional modulation of visual motion processing in cortical areas MT and MST. Nature 1996, 382, 539–541. [Google Scholar] [CrossRef]
- Moffat, S.D. Aging and spatial navigation: What do we know and where do we go? Neuropsychol. Rev. 2009, 19, 478–489. [Google Scholar] [CrossRef]
- Della-Justina, H.M.; Pastorello, B.F.; Santos, T.E.G.; Pontes-Neto, O.M.; Santos, T.E.G.; Baffa, O.; Colafêmina, J.F.; Leite, J.P.; Araujo, D.B. Human variability of fMRI brain activation in response to oculomotor stimuli. Brain Topogr. 2008, 20, 113–121. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OF-Sensitive Region | MNI152 Coordinates (mm) |
|---|---|
| LCSv | −12, −20, 42 |
| RCSv | −12, −22, 44 |
| LMT+ | −38, −64, 2 |
| RMT+ | 44, −54, 2 |
| LPIVC | −40, −34, 20 |
| RPIVC | 40, −30, 20 |
| RPc | 14, −42, 56 |
| LpV6 | −12, −78, 34 |
| RpV6 | 20, −74, 36 |
| LpVIP | −22, −62, 62 |
| RpVIP | 22, −60, 62 |
| Measure | Young (n = 29) | Aged (n = 22) |
|---|---|---|
| Age (years) | 25.2 ± 3.42 (20–34) | 70 ± 4.87 (62–80) *** |
| Sex (F/M) | 17 F/12 M | 16 F/6 M |
| Education (years completed) | 17.5 ± 2.16 (14–24) | 18.4 ± 2.81 (12–24) |
| Handedness (R/L) | 24 R/5 L | 22 R/0 L * |
| MoCA(education-adjusted total score) | n/a | 27.8 ± 1.54 (25–30) |
| VPI Accuracy (% Correct) | 97.0% (±5.44) (75%−100%) | 80.4% (±22.8) (31.3%−100%) ** |
| TC Accuracy (% Correct) | 96.8% (±5.69) (81.3%−100%) | 91.8% (±13.3) (50%−100%) |
| Perceived VPI Accuracy | 1.76 (±1.15) (1–6) | 2.77 (±1.60) (1–7) ** |
| Perceived VPI Difficulty | 2.24 (±1.24) (1–5) | 3.96 (±1.50) (2–6) ** |
| Effort Exerted VPI | 2.69 (±1.23) (1–5) | 4.36 (±1.47) (2–7) ** |
| Perceived TC Accuracy | 1.90 (±0.860) (1–4) | 2.32 (±1.59) (1–6) |
| Perceived TC Difficulty | 2.00 (±0.802) (1–4) | 2.50 (±1.26) (1–5) |
| Effort Exerted TC | 2.28 (±1.07) (1–4) | 2.59 (±1.50) (1–5) |
| Neighborhood Familiarity (% Familiar) | 58.6% | 50% |
| Familiarity Not Helpful (% Reporting Not Helpful) | 76.5% | 90.9% |
| Updating Strategy (% Endorsing) | 82.8% | 81.8% |
| Whole Model Statistics | ||
|---|---|---|
| VPI Accuracy, VPI Activity Model | VPI Accuracy, TC Activity Model | |
| R2 | 0.421 | 0.139 |
| p value | 0.0055 ** | 0.241 |
| RMSE | 18.2 | 22.2 |
| Predictor-Specific Statistics: VPI Accuracy, VPI Activity Model | ||
| LMT+ Activity VPI | RpVIP Activity VPI | |
| Parameter estimate (PE) | 0.78 | 0.297 |
| Standard Error | 0.274 | 0.132 |
| FDR-corrected p value | 0.0205 * | 0.0368 * |
| PE Confidence Interval (lower 95%, upper 95%) | 0.207, 1.35 | 0.0201, 0.573 |
| Predictor-Specific Statistics: VPI Accuracy, TC Activity Model | ||
| LMT+ Activity TC | RpVIP Activity TC | |
| Parameter estimate (PE) | 0.602 | 0.076 |
| Standard Error | 0.366 | 0.173 |
| FDR-corrected p value | 0.233 | 0.665 |
| PE Confidence Interval (lower 95%, upper 95%) | −0.164, 1.37 | −0.286, 0.438 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajac, L.; Killiany, R. Activity Strength within Optic Flow-Sensitive Cortical Regions Is Associated with Visual Path Integration Accuracy in Aged Adults. Brain Sci. 2021, 11, 245. https://doi.org/10.3390/brainsci11020245
Zajac L, Killiany R. Activity Strength within Optic Flow-Sensitive Cortical Regions Is Associated with Visual Path Integration Accuracy in Aged Adults. Brain Sciences. 2021; 11(2):245. https://doi.org/10.3390/brainsci11020245
Chicago/Turabian StyleZajac, Lauren, and Ronald Killiany. 2021. "Activity Strength within Optic Flow-Sensitive Cortical Regions Is Associated with Visual Path Integration Accuracy in Aged Adults" Brain Sciences 11, no. 2: 245. https://doi.org/10.3390/brainsci11020245
APA StyleZajac, L., & Killiany, R. (2021). Activity Strength within Optic Flow-Sensitive Cortical Regions Is Associated with Visual Path Integration Accuracy in Aged Adults. Brain Sciences, 11(2), 245. https://doi.org/10.3390/brainsci11020245