
Probing the Brain–Body Connection Using Transcranial Magnetic Stimulation (TMS): Validating a Promising Tool to Provide Biomarkers of Neuroplasticity and Central Nervous System Function



Abstract
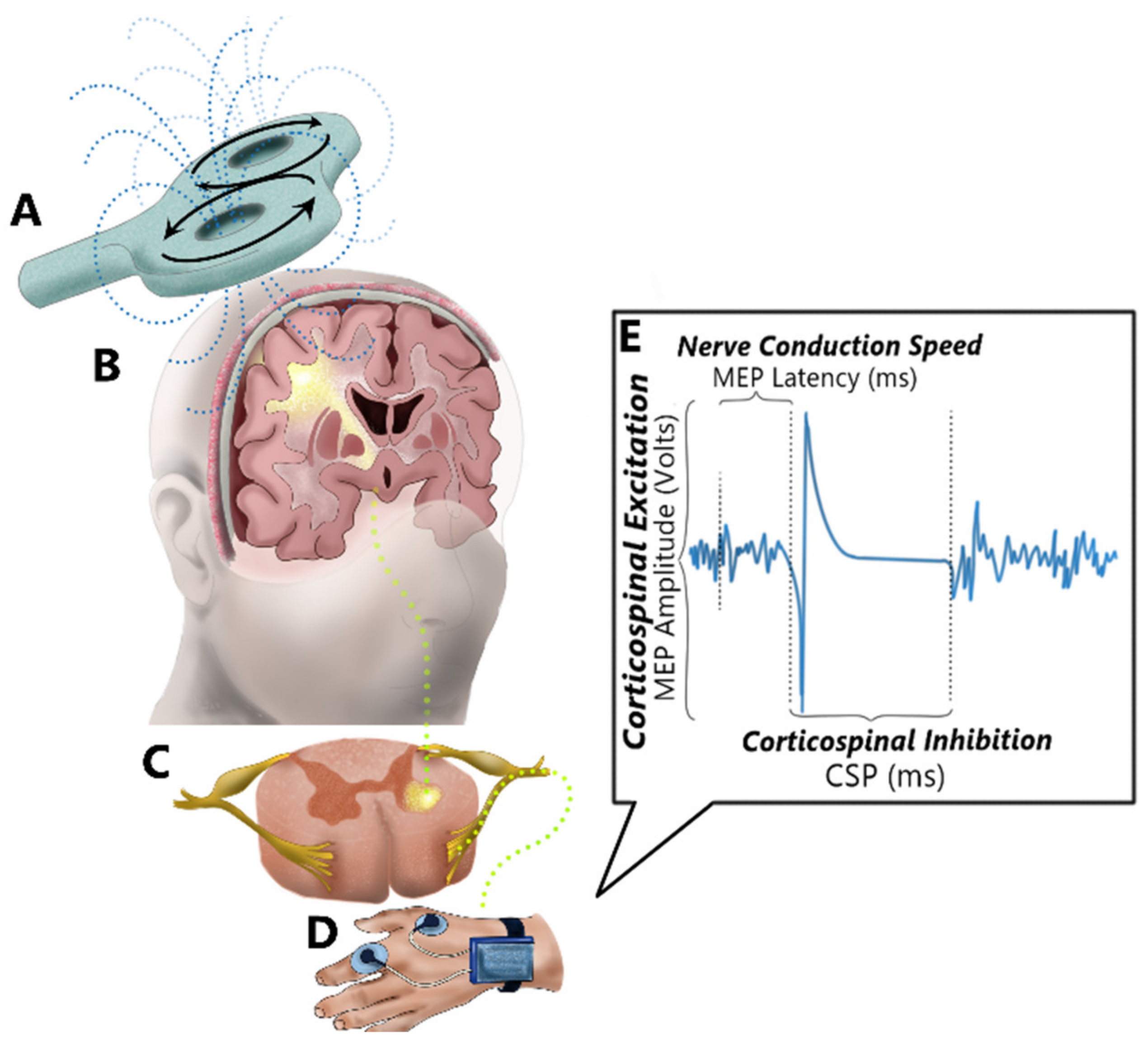
:1. Introduction
1.1. Overview of TMS Methods
1.1.1. Motor Thresholds
1.1.2. Excitatory Recruitment Curve
1.1.3. Cortical Silent Period (CSP)
1.1.4. Inhibitory Recruitment Curve
1.1.5. Transcallosal Inhibition and the Ipsilateral Silent Period
- As administering TMS in both brain hemispheres doubles the testing time, is it necessary to collect TMS variables bilaterally?
- To what extent do TMS variables correlate with clinical outcomes, specifically, motor (i.e., walking speed and nine-hole peg test (9HPT)) and non-motor function (i.e., fatigue and cognitive status)?
- Of the more than 25 variables derived from this TMS protocol, which variables are most strongly associated with severity of MS symptoms (controlling for confounding variables) and should be considered as part of a core-set?
2. Materials and Methods
2.1. Participants
2.2. Clinical Testing Procedures
2.3. TMS Testing Procedures
2.3.1. Electrode Placement and Skin Preparation
2.3.2. TMS System
2.3.3. Calibration
2.3.4. Obtaining the Motor Hotspot
2.3.5. Motor Thresholds
2.3.6. Recruitment Curves
2.3.7. Transcallosal Inhibition
2.4. TMS Post-Processing and Data Reduction
2.4.1. Motor Thresholds, MEP Latency, and AMT Symmetry
2.4.2. MEP Amplitudes and eREC Parameters
2.4.3. CSP Duration and iREC Parameters
2.4.4. Transcallosal Inhibition
2.5. Statistical Analysis
2.5.1. Differences in TMS Variables between Hemispheres
2.5.2. Examining Relationships between TMS Variables and Clinical Outcomes
2.5.3. Determining Strongest Predictors of Clinical Outcomes
2.5.4. Use of CNS-Modulating Drugs
3. Results
3.1. Participants
3.2. Excitability Differences between Hemispheres
3.3. Inhibitory Differences between Hemispheres (iREC)
3.4. MEP Latencies Not Significantly Different between Hemispheres
3.5. Transcallosal Inhibition Differences between Hemispheres
3.6. Relationships between Clinical Outcomes and TMS Variables
3.7. Best Predictors of Walking Speed
3.8. Best Predictors of Upper Extremity Dexterity (9HPT)
3.9. Best Predictors of Fatigue
3.10. Best Predictors of Cognition (SDMT)
3.11. Use of CSN-Modulating Drugs
3.12. The Core-Set
- Studies should consider assessing TMS bilaterally in order to index brain excitability asymmetries. However, the hemisphere corresponding to the weaker, or the more affected body side, should be prioritized.
- When investigating motor outcomes, studies should prioritize:
- a.
- Biomarkers of contra- and ipsilateral conduction latency.
- b.
- MEP amplitudes assessed at suprathreshold mid-high range TMS intensities (e.g., ≥125% to ≤145% of motor threshold).
- c.
- CSP assessed at lower-mid range TMS intensities (e.g., >100% to ≤115% of motor threshold).
- When investigating symptoms of fatigue, studies should prioritize:
- CSP assessed at higher range TMS intensities (e.g., ≥145% of motor threshold).
- When investigating cognition, studies should prioritize:
- a.
- RMT.
- b.
- MEP amplitudes assessed at suprathreshold mid-high range TMS intensities (e.g., ≥135% to 155% of motor threshold).
4. Discussion
4.1. Asymmetry as a TMS Biomarker
4.2. Robustness of TMS Variables from Hemisphere Corresponding to Weaker Hand
4.3. Significance of Transcallosal Inhibition and Elements of the iSP
4.4. Single Pulse TMS to Investigate Potential for Neuroplasticity
4.5. How TMS Probes GABAergic-Mediated Corticospinal Inhibition
4.6. Using the Recruitment Curve to Examine Glutamatergic-Mediated Corticospinal Excitation
4.7. TMS Variables That Did Not Stand Up to Scrutiny
4.8. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Group, B.D.W. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin Pharm. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Boyd, L.A.; Hayward, K.S.; Ward, N.S.; Stinear, C.M.; Rosso, C.; Fisher, R.J.; Carter, A.R.; Leff, A.P.; Copland, D.A.; Carey, L.M.; et al. Biomarkers of stroke recovery: Consensus-based core recommendations from the Stroke Recovery and Rehabilitation Roundtable. Int. J. Stroke 2017, 12, 480–493. [Google Scholar] [CrossRef] [PubMed]
- Bielekova, B.; Martin, R. Development of biomarkers in multiple sclerosis. Brain 2004, 127, 1463–1478. [Google Scholar] [CrossRef] [Green Version]
- Dalgas, U.; Hvid, L.G.; Kwakkel, G.; Motl, R.W.; de Groot, V.; Feys, P.; Op’t Eijnde, B.; Coote, S.; Beckerman, H.; Pfeifer, K.; et al. Moving exercise research in multiple sclerosis forward (the MoXFo initiative): Developing consensus statements for research. Mult. Scler. 2020, 26, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, A.; Iodice, R.; Petracca, M.; Inglese, M.; Cerillo, I.; Cocozza, S.; Saiote, C.; Brunetti, A.; Tedeschi, E.; Manganelli, F.; et al. Upper motor neuron evaluation in multiple sclerosis patients treated with Sativex(®). Acta Neurol. Scand. 2017, 135, 442–448. [Google Scholar] [CrossRef]
- Ploughman, M.; Austin, M.W.; Glynn, L.; Corbett, D. The effects of poststroke aerobic exercise on neuroplasticity: A systematic review of animal and clinical studies. Transl. Stroke Res. 2015, 6, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Ziemssen, T.; Akgun, K.; Bruck, W. Molecular biomarkers in multiple sclerosis. J. Neuroinflamm. 2019, 16, 272. [Google Scholar] [CrossRef] [Green Version]
- Barnett, Y.; Garber, J.Y.; Barnett, M.H. MRI biomarkers of disease progression in multiple sclerosis: Old dog, new tricks? Quant. Imaging Med. Surg. 2020, 10, 527–532. [Google Scholar] [CrossRef]
- Filippi, M.; Preziosa, P.; Banwell, B.L.; Barkhof, F.; Ciccarelli, O.; De Stefano, N.; Geurts, J.J.G.; Paul, F.; Reich, D.S.; Toosy, A.T.; et al. Assessment of lesions on magnetic resonance imaging in multiple sclerosis: Practical guidelines. Brain 2019, 142, 1858–1875. [Google Scholar] [CrossRef] [Green Version]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Ziemann, U.; Reis, J.; Schwenkreis, P.; Rosanova, M.; Strafella, A.; Badawy, R.; Muller-Dahlhaus, F. TMS and drugs revisited 2014. Clin. Neurophysiol. 2015, 126, 1847–1868. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.; Mooney, R.A.; Ackerley, S.J.; Barber, P.A.; Borges, V.M.; Clarkson, A.N.; Mangold, C.; Ren, A.; Smith, M.C.; Stinear, C.M.; et al. Neurochemical balance and inhibition at the subacute stage after stroke. J. Neurophysiol. 2020, 123, 1775–1790. [Google Scholar] [CrossRef]
- Groppa, S.; Oliviero, A.; Eisen, A.; Quartarone, A.; Cohen, L.G.; Mall, V.; Kaelin-Lang, A.; Mima, T.; Rossi, S.; Thickbroom, G.W.; et al. A practical guide to diagnostic transcranial magnetic stimulation: Report of an IFCN committee. Clin. Neurophysiol. 2012, 123, 858–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridding, M.C.; Rothwell, J.C. Stimulus/response curves as a method of measuring motor cortical excitability in man. Electroencephalogr. Clin. Neurophysiol. 1997, 105, 340–344. [Google Scholar] [CrossRef]
- Devanne, H.; Lavoie, B.A.; Capaday, C. Input-output properties and gain changes in the human corticospinal pathway. Exp. Brain Res. 1997, 114, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Capaday, C. Neurophysiological methods for studies of the motor system in freely moving human subjects. J. Neurosci. Methods 1997, 74, 201–218. [Google Scholar] [CrossRef]
- Carson, R.G.; Nelson, B.D.; Buick, A.R.; Carroll, T.J.; Kennedy, N.C.; Cann, R.M. Characterizing changes in the excitability of corticospinal projections to proximal muscles of the upper limb. Brain Stimul. 2013, 6, 760–768. [Google Scholar] [CrossRef]
- Talelli, P.; Ewas, A.; Waddingham, W.; Rothwell, J.C.; Ward, N.S. Neural correlates of age-related changes in cortical neurophysiology. NeuroImage 2008, 40, 1772–1781. [Google Scholar] [CrossRef] [Green Version]
- Potter-Baker, K.A.; Varnerin, N.M.; Cunningham, D.A.; Roelle, S.M.; Sankarasubramanian, V.; Bonnett, C.E.; Machado, A.G.; Conforto, A.B.; Sakaie, K.; Plow, E.B. Influence of Corticospinal Tracts from Higher Order Motor Cortices on Recruitment Curve Properties in Stroke. Front. Neurosci. 2016, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Kemlin, C.; Moulton, E.; Leder, S.; Houot, M.; Meunier, S.; Rosso, C.; Lamy, J.C. Redundancy among parameters describing the input-output relation of motor evoked potentials in healthy subjects and stroke patients. Front. Neurol. 2019, 10, 535. [Google Scholar] [CrossRef]
- Kukke, S.N.; Paine, R.W.; Chao, C.C.; de Campos, A.C.; Hallett, M. Efficient and reliable characterization of the corticospinal system using transcranial magnetic stimulation. J. Clin. Neurophysiol. 2014, 31, 246–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, L.; Koshy, P.J.; Steel, A.; Bageac, D.; Schintu, S.; Wassermann, E.M. Motor cortex inhibition by TMS reduces cognitive non-motor procedural learning when immediate incentives are present. Cortex 2017, 97, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, A.N.; Huang, B.S.; Macisaac, S.E.; Mody, I.; Carmichael, S.T. Reducing excessive GABA-mediated tonic inhibition promotes functional recovery after stroke. Nature 2010, 468, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yacyshyn, A.F.; Woo, E.J.; Price, M.C.; McNeil, C.J. Motoneuron responsiveness to corticospinal tract stimulation during the silent period induced by transcranial magnetic stimulation. Exp. Brain Res. 2016, 234, 3457–3463. [Google Scholar] [CrossRef]
- Hupfeld, K.E.; Swanson, C.W.; Fling, B.W.; Seidler, R.D. TMS-induced silent periods: A review of methods and call for consistency. J. Neurosci. Methods 2020, 346, 108950. [Google Scholar] [CrossRef]
- Škarabot, J.; Mesquita, R.N.O.; Brownstein, C.G.; Ansdell, P. Myths and methodologies: How loud is the story told by the transcranial magnetic stimulation-evoked silent period? Exp. Physiol. 2019, 104, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Snow, N.J.; Wadden, K.P.; Chaves, A.R.; Ploughman, M. Transcranial Magnetic Stimulation as a Potential Biomarker in Multiple Sclerosis: A Systematic Review with Recommendations for Future Research. Neural Plast. 2019, 2019, 6430596. [Google Scholar] [CrossRef] [Green Version]
- Wassermann, E.M.; Fuhr, P.; Cohen, L.G.; Hallett, M. Effects of transcranial magnetic stimulation on ipsilateral muscles. Neurology 1991, 41, 1795–1799. [Google Scholar] [CrossRef]
- Ferbert, A.; Priori, A.; Rothwell, J.C.; Day, B.L.; Colebatch, J.G.; Marsden, C.D. Interhemispheric inhibition of the human motor cortex. J. Physiol. 1992, 453, 525–546. [Google Scholar] [CrossRef]
- Di Pino, G.; Pellegrino, G.; Assenza, G.; Capone, F.; Ferreri, F.; Formica, D.; Ranieri, F.; Tombini, M.; Ziemann, U.; Rothwell, J.C.; et al. Modulation of brain plasticity in stroke: A novel model for neurorehabilitation. Nat. Rev. Neurol. 2014, 10, 597–608. [Google Scholar] [CrossRef]
- Llufriu, S.; Blanco, Y.; Martinez-Heras, E.; Casanova-Molla, J.; Gabilondo, I.; Sepulveda, M.; Falcon, C.; Berenguer, J.; Bargallo, N.; Villoslada, P.; et al. Influence of corpus callosum damage on cognition and physical disability in multiple sclerosis: A multimodal study. PLoS ONE 2012, 7, e37167. [Google Scholar] [CrossRef] [PubMed]
- Neva, J.L.; Lakhani, B.; Brown, K.E.; Wadden, K.P.; Mang, C.S.; Ledwell, N.H.; Borich, M.R.; Vavasour, I.M.; Laule, C.; Traboulsee, A.L.; et al. Multiple measures of corticospinal excitability are associated with clinical features of multiple sclerosis. Behav. Brain Res. 2016, 297, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, L.D.; Milot, M.H. Changes in transcranial magnetic stimulation outcome measures in response to upper-limb physical training in stroke: A systematic review of randomized controlled trials. Ann. Phys. Rehabil. Med. 2018, 61, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Hayward, K.S.; Neva, J.L.; Mang, C.S.; Peters, S.; Wadden, K.P.; Ferris, J.K.; Boyd, L.A. Interhemispheric Pathways Are Important for Motor Outcome in Individuals with Chronic and Severe Upper Limb Impairment Post Stroke. Neural Plast. 2017, 2017, 4281532. [Google Scholar] [CrossRef] [PubMed]
- Iodice, R.; Manganelli, F.; Dubbioso, R. The therapeutic use of non-invasive brain stimulation in multiple sclerosis—A review. Restor. Neurol. Neurosci. 2017, 35, 497–509. [Google Scholar] [CrossRef]
- Criste, G.; Trapp, B.; Dutta, R. Axonal loss in multiple sclerosis: Causes and mechanisms. Handb. Clin. Neurol. 2014, 122, 101–113. [Google Scholar] [CrossRef]
- Dutta, R.; Trapp, B.D. Relapsing and progressive forms of multiple sclerosis: Insights from pathology. Curr. Opin. Neurol. 2014, 27, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahad, D.H.; Trapp, B.D.; Lassmann, H. Pathological mechanisms in progressive multiple sclerosis. Lancet Neurol. 2015, 14, 183–193. [Google Scholar] [CrossRef]
- Trapp, B.D.; Vignos, M.; Dudman, J.; Chang, A.; Fisher, E.; Staugaitis, S.M.; Battapady, H.; Mork, S.; Ontaneda, D.; Jones, S.E.; et al. Cortical neuronal densities and cerebral white matter demyelination in multiple sclerosis: A retrospective study. Lancet Neurol. 2018, 17, 870–884. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.H.; Leary, S.M. Primary-progressive multiple sclerosis. Lancet Neurol. 2007, 6, 903–912. [Google Scholar] [CrossRef]
- Lublin, F.D. New multiple sclerosis phenotypic classification. Eur. Neurol. 2014, 72 (Suppl. 1), 1–5. [Google Scholar] [CrossRef]
- Kappos, L.; De Stefano, N.; Freedman, M.S.; Cree, B.A.C.; Radue, E.W.; Sprenger, T.; Sormani, M.P.; Smith, T.; Haring, D.A.; Meier, D.P.; et al. Inclusion of brain volume loss in a revised measure of ‘no evidence of disease activity’ (NEDA-4) in relapsing-remitting multiple sclerosis. Mult. Scler. 2016, 22, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.L.; Wilson, T.E.; White, A.T.; Frohman, E.M. Thermoregulation in multiple sclerosis. J. Appl. Physiol. 2010, 109, 1531–1537. [Google Scholar] [CrossRef]
- Bollaert, R.E.; Motl, R.W. Physical and Cognitive Functions, Physical Activity, and Sedentary Behavior in Older Adults With Multiple Sclerosis. J. Geriatr. Phys. Ther. 2019, 42, 304–312. [Google Scholar] [CrossRef]
- Motl, R.W.; Gappmaier, E.; Nelson, K.; Benedict, R.H. Physical activity and cognitive function in multiple sclerosis. J. Sport Exerc. Psychol. 2011, 33, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, L.; Sgaramella, T.M.; Bortolon, F.; Stenta, G.; Fornaro, L.; Cracco, A.; Perini, F.; Soresi, S. Determinants of on-the-job-barriers in employed persons with multiple sclerosis: The role of disability severity and cognitive indices. Work 2014, 47, 509–520. [Google Scholar] [CrossRef]
- Ben Ari Shevil, E.; Johansson, S.; Ytterberg, C.; Bergstrom, J.; von Koch, L. How are cognitive impairment, fatigue and signs of depression related to participation in daily life among persons with multiple sclerosis? Disabil. Rehabil. 2014, 36, 2012–2018. [Google Scholar] [CrossRef]
- Heesen, C.; Böhm, J.; Reich, C.; Kasper, J.; Goebel, M.; Gold, S.M. Patient perception of bodily functions in multiple sclerosis: Gait and visual function are the most valuable. Mult. Scler. 2008, 14, 988–991. [Google Scholar] [CrossRef]
- Paltamaa, J.; Sarasoja, T.; Leskinen, E.; Wikström, J.; Mälkiä, E. Measures of physical functioning predict self-reported performance in self-care, mobility, and domestic life in ambulatory persons with multiple sclerosis. Arch. Phys. Med. Rehabil. 2007, 88, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.R.; Devasahayam, A.J.; Riemenschneider, M.; Pretty, R.W.; Ploughman, M. Walking Training Enhances Corticospinal Excitability in Progressive Multiple Sclerosis—A Pilot Study. Front. Neurol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Benedict, R.H.; DeLuca, J.; Phillips, G.; LaRocca, N.; Hudson, L.D.; Rudick, R.; Multiple Sclerosis Outcome Assessments, C. Validity of the Symbol Digit Modalities Test as a cognition performance outcome measure for multiple sclerosis. Mult. Scler. 2017, 23, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Drake, A.S.; Weinstock-Guttman, B.; Morrow, S.A.; Hojnacki, D.; Munschauer, F.E.; Benedict, R.H. Psychometrics and normative data for the Multiple Sclerosis Functional Composite: Replacing the PASAT with the Symbol Digit Modalities Test. Mult. Scler. 2010, 16, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Mathiowetz, V.; Weber, K.; Kashman, N.; Volland, G. Adult norms for the Nine Hole Peg Test of finger dexterity. Occup. Ther. J. Res. 1985, 5, 24–38. [Google Scholar] [CrossRef]
- Feys, P.; Lamers, I.; Francis, G.; Benedict, R.; Phillips, G.; LaRocca, N.; Hudson, L.D.; Rudick, R.; Outcome, M.S. The Nine-Hole Peg Test as a manual dexterity performance measure for multiple sclerosis. Mult. Scler. 2017, 23, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Chaves, A.R.; Wallack, E.M.; Kelly, L.P.; Pretty, R.W.; Wiseman, H.D.; Chen, A.; Moore, C.S.; Stefanelli, M.; Ploughman, M. Asymmetry of Brain Excitability: A New Biomarker that Predicts Objective and Subjective Symptoms in Multiple Sclerosis. Behav. Brain Res. 2019, 359, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Chaves, A.R.; Devasahayam, A.J.; Kelly, L.P.; Pretty, R.W.; Ploughman, M. Exercise-Induced Brain Excitability Changes in Progressive Multiple Sclerosis: A Pilot Study. J. Neurol. Phys. Ther. 2020, 44, 132–144. [Google Scholar] [CrossRef]
- Bohannon, R.W. Comfortable and maximum walking speed of adults aged 20–79 years: Reference values and determinants. Age Ageing 1997, 26, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Palmer, E.; Ashby, P. Corticospinal projections to upper limb motoneurones in humans. J. Physiol. 1992, 448, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Yao, B.; Klein, C.S.; Hu, H.; Li, S.; Zhou, P. Motor Unit Properties of the First Dorsal Interosseous in Chronic Stroke Subjects: Concentric Needle and Single Fiber EMG Analysis. Front. Physiol. 2018, 9, 1587. [Google Scholar] [CrossRef] [Green Version]
- Fritz, N.E.; Keller, J.; Calabresi, P.A.; Zackowski, K.M. Quantitative measures of walking and strength provide insight into brain corticospinal tract pathology in multiple sclerosis. Neuroimage. Clin. 2017, 14, 490–498. [Google Scholar] [CrossRef]
- DeLuca, G.C.; Ebers, G.C.; Esiri, M.M. Axonal loss in multiple sclerosis: A pathological survey of the corticospinal and sensory tracts. Brain 2004, 127, 1009–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thickbroom, G.W.; Byrnes, M.L.; Archer, S.A.; Kermode, A.G.; Mastaglia, F.L. Corticomotor organisation and motor function in multiple sclerosis. J. Neurol. 2005, 252, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Holobar, A.; Gazzoni, M.; Merletti, R.; Rymer, W.Z.; Zhou, P. Examination of Poststroke Alteration in Motor Unit Firing Behavior Using High-Density Surface EMG Decomposition. IEEE Trans. Bio-Med. Eng. 2015, 62, 1242–1252. [Google Scholar] [CrossRef]
- Singh, A.M.; Duncan, R.E.; Neva, J.L.; Staines, W.R. Aerobic exercise modulates intracortical inhibition and facilitation in a nonexercised upper limb muscle. BMC Sports Sci. Med. Rehabil. 2014, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.A.; Stapley, P.J.; Shemmell, J.B.H.; Lepers, R.; McAndrew, D.J. Global Corticospinal Excitability as Assessed in A Non-Exercised Upper Limb Muscle Compared Between Concentric and Eccentric Modes of Leg Cycling. Sci. Rep. 2019, 9, 19212. [Google Scholar] [CrossRef]
- Vucic, S.; Kiernan, M.C. Transcranial Magnetic Stimulation for the Assessment of Neurodegenerative Disease. Neurotherapeutics 2017, 14, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Chen, R. Transcranial magnetic stimulation to understand pathophysiology and as potential treatment for neurodegenerative diseases. Transl. Neurodegener. 2015, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Di Lazzaro, V.; Oliviero, A.; Saturno, E.; Pilato, F.; Insola, A.; Mazzone, P.; Profice, P.; Tonali, P.; Rothwell, J.C. The effect on corticospinal volleys of reversing the direction of current induced in the motor cortex by transcranial magnetic stimulation. Exp. Brain Res. 2001, 138, 268–273. [Google Scholar] [CrossRef]
- Hannah, R.; Rothwell, J.C. Pulse Duration as Well as Current Direction Determines the Specificity of Transcranial Magnetic Stimulation of Motor Cortex during Contraction. Brain Stimul. 2017, 10, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Fonov, V.; Evans, A.C.; Botteron, K.; Almli, C.R.; McKinstry, R.C.; Collins, D.L.; Brain Development Cooperative, G. Unbiased average age-appropriate atlases for pediatric studies. NeuroImage 2011, 54, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Collins, D.L.; Neelin, P.; Peters, T.M.; Evans, A.C. Automatic 3D intersubject registration of MR volumetric data in standardized Talairach space. J. Comput. Assist. Tomogr. 1994, 18, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Jonker, Z.D.; van der Vliet, R.; Hauwert, C.M.; Gaiser, C.; Tulen, J.H.M.; van der Geest, J.N.; Donchin, O.; Ribbers, G.M.; Frens, M.A.; Selles, R.W. TMS motor mapping: Comparing the absolute reliability of digital reconstruction methods to the golden standard. Brain Stimul. 2019, 12, 309–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, C.; Arai, N.; Lücke, J.; Ziemann, U. Hysteresis effects on the input-output curve of motor evoked potentials. Clin. Neurophysiol. 2009, 120, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Centeno, C.; Medeiros, D.; Beck, M.M.; Lugassy, L.; Gonzalez, D.F.; Nepveu, J.F.; Roig, M. The effects of aging on cortico-spinal excitability and motor memory consolidation. Neurobiol. Aging 2018, 70, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Kidgell, D.J.; Pearce, A.J. What has transcranial magnetic stimulation taught us about neural adaptations to strength training? A brief review. J. Strength Cond. Res. Natl. Strength Cond. Assoc. 2011, 25, 3208–3217. [Google Scholar] [CrossRef]
- Bailey, A.Z.; Mi, Y.P.; Nelson, A.J. Short-latency afferent inhibition in chronic spinal cord injury. Transl. Neurosci. 2015, 6, 235–243. [Google Scholar] [CrossRef]
- Leon-Sarmiento, F.E.; Rizzo-Sierra, C.V.; Bayona, E.A.; Bayona-Prieto, J.; Doty, R.L.; Bara-Jimenez, W. Novel mechanisms underlying inhibitory and facilitatory transcranial magnetic stimulation abnormalities in Parkinson’s disease. Arch. Med Res. 2013, 44, 221–228. [Google Scholar] [CrossRef]
- Werhahn, K.J.; Behrang-Nia, M.; Bott, M.C.; Klimpe, S. Does the recruitment of excitation and inhibition in the motor cortex differ? J. Clin. Neurophysiol. 2007, 24, 419–423. [Google Scholar] [CrossRef]
- Lenzi, D.; Conte, A.; Mainero, C.; Frasca, V.; Fubelli, F.; Totaro, P.; Caramia, F.; Inghilleri, M.; Pozzilli, C.; Pantano, P. Effect of corpus callosum damage on ipsilateral motor activation in patients with multiple sclerosis: A functional and anatomical study. Hum. Brain Mapp. 2007, 28, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Davidson, T.; Tremblay, F. Age and hemispheric differences in transcallosal inhibition between motor cortices: An ispsilateral silent period study. BMC Neurosci. 2013, 14, 62. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.-L.; Dubuc, T.; Boufadel, D.F.; Fisher, B.E. Measuring ipsilateral silent period: Effects of muscle contraction levels and quantification methods. Brain Res. 2017, 1674, 77–83. [Google Scholar] [CrossRef]
- Meyer, B.-U.; Röricht, S.; Gräfin von Einsiedel, H.; Kruggel, F.; Weindl, A. Inhibitory and excitatory interhemispheric transfers between motor cortical areas in normal humans and patients with abnormalities of the corpus callosum. Brain 1995, 118, 429–440. [Google Scholar] [CrossRef]
- Kesar, T.M.; Eicholtz, S.; Lin, B.J.; Wolf, S.L.; Borich, M.R. Effects of posture and coactivation on corticomotor excitability of ankle muscles. Restor. Neurol. Neurosci. 2018, 36, 131–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finisguerra, A.; Maffongelli, L.; Bassolino, M.; Jacono, M.; Pozzo, T.; D’Ausilio, A. Generalization of motor resonance during the observation of hand, mouth, and eye movements. J. Neurophysiol. 2015, 114, 2295–2304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobimatsu, S.; Sun, S.J.; Fukui, R.; Kato, M. Effects of sex, height and age on motor evoked potentials with magnetic stimulation. J. Neurol. 1998, 245, 256–261. [Google Scholar] [CrossRef]
- Livingston, S.C.; Friedlander, D.L.; Gibson, B.C.; Melvin, J.R. Motor evoked potential response latencies demonstrate moderate correlations with height and limb length in healthy young adults. Neurodiagn. J. 2013, 53, 63–78. [Google Scholar] [PubMed]
- Schättin, A.; Gennaro, F.; Egloff, M.; Vogt, S.; de Bruin, E.D. Physical Activity, Nutrition, Cognition, Neurophysiology, and Short-Time Synaptic Plasticity in Healthy Older Adults: A Cross-Sectional Study. Front. Aging Neurosci. 2018, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Mang, C.S.; Snow, N.J.; Campbell, K.L.; Ross, C.J.; Boyd, L.A. A single bout of high-intensity aerobic exercise facilitates response to paired associative stimulation and promotes sequence-specific implicit motor learning. J. Appl. Physiol. 2014, 117, 1325–1336. [Google Scholar] [CrossRef]
- Rosenkranz, K.; Kacar, A.; Rothwell, J.C. Differential modulation of motor cortical plasticity and excitability in early and late phases of human motor learning. J. Neurosci. 2007, 27, 12058–12066. [Google Scholar] [CrossRef] [Green Version]
- Fleming, M.K.; Newham, D.J. Reliability of Transcallosal Inhibition in Healthy Adults. Front. Hum. Neurosci. 2016, 10, 681. [Google Scholar] [CrossRef] [Green Version]
- Daskalakis, Z.J.; Molnar, G.F.; Christensen, B.K.; Sailer, A.; Fitzgerald, P.B.; Chen, R. An automated method to determine the transcranial magnetic stimulation-induced contralateral silent period. Clin. Neurophysiol. 2003, 114, 938–944. [Google Scholar] [CrossRef]
- Schmierer, K.; Irlbacher, K.; Grosse, P.; Roricht, S.; Meyer, B.U. Correlates of disability in multiple sclerosis detected by transcranial magnetic stimulation. Neurology 2002, 59, 1218–1224. [Google Scholar] [CrossRef]
- Giovannelli, F.; Borgheresi, A.; Balestrieri, F.; Zaccara, G.; Viggiano, M.P.; Cincotta, M.; Ziemann, U. Modulation of interhemispheric inhibition by volitional motor activity: An ipsilateral silent period study. J. Physiol. 2009, 587, 5393–5410. [Google Scholar] [CrossRef]
- Jung, P.; Ziemann, U. Differences of the ipsilateral silent period in small hand muscles. Muscle Nerve 2006, 34, 431–436. [Google Scholar] [CrossRef]
- Harris-Love, M.L.; Morton, S.M.; Perez, M.A.; Cohen, L.G. Mechanisms of short-term training-induced reaching improvement in severely hemiparetic stroke patients: A TMS study. Neurorehabilit. Neural Repair 2011, 25, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Sheskin, D.J. Handbook of Parametric and Nonparametric Statistical Procedures, 3rd ed.; CRC Press: New York, NY, USA, 2003. [Google Scholar]
- Kirkland, M.C.; Wallack, E.M.; Rancourt, S.N.; Ploughman, M. Comparing three dual-task methods and the relationship to physical and cognitive impairment in people with multiple sclerosis and controls. Mult. Scler. Int. 2015, 2015, 650645. [Google Scholar] [CrossRef] [Green Version]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Tabachnick, B.G.; Fidell, L.S. Using Multivariate Statistics; Pearson Education: Boston, MA, USA, 2013. [Google Scholar]
- Vatcheva, K.P.; Lee, M.; McCormick, J.B.; Rahbar, M.H. Multicollinearity in Regression Analyses Conducted in Epidemiologic Studies. Epidemiology 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Salamon, S.J.; Hansen, H.J.; Abbott, D. How real are observed trends in small correlated datasets? R. Soc. Open Sci. 2019, 6, 181089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, J.; Cohen, P.; West, S.G.; Aiken, L.S. Applied Multiple Regression/Correlation Analysis for the Behavioral Sciences, 3rd ed.; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 2003; pp. xxviii, 703–xxviii, 703. [Google Scholar]
- Moore, D.S.; Notz, W.I.; Flinger, M.A. The Basic Practice of Statistics, 6th ed.; W. H. Freeman and Company: New York, NY, USA, 2013. [Google Scholar]
- Cook, R.D.; Weisberg, S. Residuals and Influence in Regression; Chapman and Hall: New York, NY, USA, 1982. [Google Scholar]
- Bollen, K.A.; Jackman, R.W. Regression diagnostics: An expository treatment of outliers and influential cases. Sociol. Methods Res. 1985, 13, 510–542. [Google Scholar] [CrossRef]
- Burke, S. Missing values, outliers, robust statistics & non-parametric methods. Sci. Data Manag. 1998, 1, 32–38. [Google Scholar]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Li, C.; Liu, F.; Peng, H.; Huang, Y.; Song, X.; Xie, Q.; Li, Y.; Liu, Y. The positive effect of venlafaxine on central motor conduction. Clin. Neurol. Neurosurg. 2018, 167, 65–69. [Google Scholar] [CrossRef]
- Santarnecchi, E.; Rossi, S.; Bartalini, S.; Cincotta, M.; Giovannelli, F.; Tatti, E.; Ulivelli, M. Neurophysiological Correlates of Central Fatigue in Healthy Subjects and Multiple Sclerosis Patients before and after Treatment with Amantadine. Neural Plast. 2015, 2015, 616242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priori, A.; Berardelli, A.; Inghilleri, M.; Accornero, N.; Manfredi, M. Motor cortical inhibition and the dopaminergic system: Pharmacological changes in the silent period after transcranial brain stimulation in normal subjects, patients with Parkinson’s disease and drug-induced parkinsonism. Brain 1994, 117, 317–323. [Google Scholar] [CrossRef]
- Rizzo, V.; Crupi, D.; Bagnato, S.; Quartarone, A.; Benvenga, S.; Bartolone, L.; Ghilardi, M.F.; Trimarchi, F.; Girlanda, P.; Battaglia, F. Neural response to transcranial magnetic stimulation in adult hypothyroidism and effect of replacement treatment. J. Neurol. Sci. 2008, 266, 38–43. [Google Scholar] [CrossRef]
- Münchau, A.; Langosch, J.M.; Gerschlager, W.; Rothwell, J.C.; Orth, M.; Trimble, M.R. Mirtazapine increases cortical excitability in healthy controls and epilepsy patients with major depression. J. Neurol. Neurosurg. Psychiatry 2005, 76, 527. [Google Scholar] [CrossRef]
- Mufti, M.A.; Holtzheimer, P.E., III; Epstein, C.M.; Quinn, S.C.; Vito, N.; McDonald, W.M. Bupropion decreases resting motor threshold: A case report. Brain Stimul. 2010, 3, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Monte-Silva, K.; Ruge, D.; Teo, J.T.; Paulus, W.; Rothwell, J.C.; Nitsche, M.A. D2 Receptor Block Abolishes Theta Burst Stimulation-Induced Neuroplasticity in the Human Motor Cortex. Neuropsychopharmacology 2011, 36, 2097–2102. [Google Scholar] [CrossRef] [PubMed]
- Manganotti, P.; Bortolomasi, M.; Zanette, G.; Pawelzik, T.; Giacopuzzi, M.; Fiaschi, A. Intravenous clomipramine decreases excitability of human motor cortex. A study with paired magnetic stimulation. J. Neurol. Sci. 2001, 184, 27–32. [Google Scholar] [CrossRef]
- Schoonheim, M.M.; Filippi, M. Functional plasticity in MS: Friend or foe? Neurology 2012, 79, 1418–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajda, C.; Pukoli, D.; Bende, Z.; Majlath, Z.; Vecsei, L. Excitotoxins, Mitochondrial and Redox Disturbances in Multiple Sclerosis. Int. J. Mol. Sci. 2017, 18, 353. [Google Scholar] [CrossRef] [Green Version]
- Murase, N.; Duque, J.; Mazzocchio, R.; Cohen, L.G. Influence of interhemispheric interactions on motor function in chronic stroke. Ann. Neurol. 2004, 55, 400–409. [Google Scholar] [CrossRef]
- Zipser, C.M.; Premoli, I.; Belardinelli, P.; Castellanos, N.; Rivolta, D.; Heidegger, T.; Müller-Dahlhaus, F.; Ziemann, U. Cortical excitability and interhemispheric connectivity in early relapsing-remitting multiple sclerosis studied with TMS-EEG. Front. Neurosci. 2018, 12, 393. [Google Scholar] [CrossRef] [Green Version]
- Nantes, J.C.; Zhong, J.; Holmes, S.A.; Whatley, B.; Narayanan, S.; Lapierre, Y.; Arnold, D.L.; Koski, L. Intracortical inhibition abnormality during the remission phase of multiple sclerosis is related to upper limb dexterity and lesions. Clin. Neurophysiol. 2016, 127, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Vucic, S.; Burke, T.; Lenton, K.; Ramanathan, S.; Gomes, L.; Yannikas, C.; Kiernan, M.C. Cortical dysfunction underlies disability in multiple sclerosis. Mult. Scler. 2012, 18, 425–432. [Google Scholar] [CrossRef]
- Zeller, D.; Dang, S.Y.; Stefan, K.; Biller, A.; Bartsch, A.; Saur, D.; Bendszus, M.; Rieckmann, P.; Toyka, K.V.; Classen, J. Functional role of ipsilateral motor areas in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2011, 82, 578–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codecà, C.; Mori, F.; Kusayanagi, H.; Monteleone, F.; Boffa, L.; Paolillo, A.; Bernardi, G.; Koch, G.; Centonze, D. Differential patterns of interhemispheric functional disconnection in mild and advanced multiple sclerosis. Mult. Scler. 2010, 16, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.; Hübers, A.; Lauterbach-Soon, B.; Hattingen, E.; Jung, P.; Cohen, L.G.; Ziemann, U. Motor callosal disconnection in early relapsing-remitting multiple sclerosis. Hum. Brain Mapp. 2011, 32, 846–855. [Google Scholar] [CrossRef]
- Luscher, C.; Malenka, R.C. NMDA receptor-dependent long-term potentiation and long-term depression (LTP/LTD). Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliss, T.V.; Cooke, S.F. Long-term potentiation and long-term depression: A clinical perspective. Clinics 2011, 66 (Suppl. 1), 3–17. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, U.; Ilic, T.V.; Pauli, C.; Meintzschel, F.; Ruge, D. Learning modifies subsequent induction of long-term potentiation-like and long-term depression-like plasticity in human motor cortex. J. Neurosci. 2004, 24, 1666–1672. [Google Scholar] [CrossRef]
- Sale, A.; Berardi, N.; Maffei, L. Environment and brain plasticity: Towards an endogenous pharmacotherapy. Physiol. Rev. 2014, 94, 189–234. [Google Scholar] [CrossRef]
- Mang, C.S.; Campbell, K.L.; Ross, C.J.; Boyd, L.A. Promoting neuroplasticity for motor rehabilitation after stroke: Considering the effects of aerobic exercise and genetic variation on brain-derived neurotrophic factor. Phys. Ther. 2013, 93, 1707–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mott, D.D.; Lewis, D.V. The pharmacology and function of central GABAB receptors. Int. Rev. Neurobiol. 1994, 36, 97–223. [Google Scholar] [PubMed]
- Kullmann, D.M.; Moreau, A.W.; Bakiri, Y.; Nicholson, E. Plasticity of inhibition. Neuron 2012, 75, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Epstein, C.M.; Wassermann, E.M.; Ziemann, U.; Wolters, A.; Ziemann, U.; Benecke, R. The Cortical Silent Period; Oxford University Press: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Classen, J.; Schnitzler, A.; Binkofski, F.; Werhahn, K.J.; Kim, Y.S.; Kessler, K.R.; Benecke, R. The motor syndrome associated with exaggerated inhibition within the primary motor cortex of patients with hemiparetic. Brain 1997, 120 Pt 4, 605–619. [Google Scholar] [CrossRef] [Green Version]
- Sale, M.V.; Ridding, M.C.; Nordstrom, M.A. Factors influencing the magnitude and reproducibility of corticomotor excitability changes induced by paired associative stimulation. Exp. Brain Res. 2007, 181, 615–626. [Google Scholar] [CrossRef]
- Klein, P.A.; Petitjean, C.; Olivier, E.; Duque, J. Top-down suppression of incompatible motor activations during response selection under conflict. NeuroImage 2014, 86, 138–149. [Google Scholar] [CrossRef]
- Liepert, J.; Mingers, D.; Heesen, C.; Baumer, T.; Weiller, C. Motor cortex excitability and fatigue in multiple sclerosis: A transcranial magnetic stimulation study. Mult. Scler. 2005, 11, 316–321. [Google Scholar] [CrossRef]
- Stagg, C.J.; Bestmann, S.; Constantinescu, A.O.; Moreno, L.M.; Allman, C.; Mekle, R.; Woolrich, M.; Near, J.; Johansen-Berg, H.; Rothwell, J.C. Relationship between physiological measures of excitability and levels of glutamate and GABA in the human motor cortex. J. Physiol. 2011, 589, 5845–5855. [Google Scholar] [CrossRef] [PubMed]
- Davey, N.J.; Smith, H.C.; Savic, G.; Maskill, D.W.; Ellaway, P.H.; Frankel, H.L. Comparison of input-output patterns in the corticospinal system of normal subjects and incomplete spinal cord injured patients. Exp. Brain Res. 1999, 127, 382–390. [Google Scholar] [CrossRef]
- Magistris, M.R.; Rosler, K.M.; Truffert, A.; Myers, J.P. Transcranial stimulation excites virtually all motor neurons supplying the target muscle. A demonstration and a method improving the study of motor evoked potentials. Brain 1998, 121, 437–450. [Google Scholar] [CrossRef] [Green Version]
- Tataroglu, C.; Genc, A.; Idiman, E.; Cakmur, R.; Idiman, F. Cortical relay time for long latency reflexes in patients with definite multiple sclerosis. Can. J. Neurol. Sci. 2004, 31, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Tataroglu, C.; Genc, A.; Idiman, E.; Cakmur, R.; Idiman, F. Cortical silent period and motor evoked potentials in patients with multiple sclerosis. Clin. Neurol. Neurosurg. 2003, 105, 105–110. [Google Scholar] [CrossRef]
- Zeller, D.; Aufm Kampe, K.; Biller, A.; Stefan, K.; Gentner, R.; Schutz, A.; Bartsch, A.; Bendszus, M.; Toyka, K.V.; Rieckmann, P.; et al. Rapid-onset central motor plasticity in multiple sclerosis. Neurology 2010, 74, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Stampanoni Bassi, M.; Buttari, F.; Gilio, L.; De Paolis, N.; Fresegna, D.; Centonze, D.; Iezzi, E. Inflammation and Corticospinal Functioning in Multiple Sclerosis: A TMS Perspective. Front. Neurol. 2020, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Caramia, M.D.; Desiato, M.T.; Cicinelli, P.; Iani, C.; Rossini, P.M. Latency jump of “relaxed” versus “contracted” motor evoked potentials as a marker of cortico-spinal maturation. Electroencephalogr. Clin. Neurophysiol. 1993, 89, 61–66. [Google Scholar] [CrossRef]
- Lemon, R.N.; Baker, S.N.; Davis, J.A.; Kirkwood, P.A.; Maier, M.A.; Yang, H.S. The importance of the cortico-motoneuronal system for control of grasp. Novartis Found. Symp. 1998, 218, 202–215, discussion 215–208. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Participants (n = 110) | Higher Levels of Disability Group (n = 34) | Lower Levels of Disability Group (n = 76) | ||
|---|---|---|---|---|
| Sex (Female/Male) | 80/30 | 23/11 | 57/19 | |
| MS Type (n) | RRMS | 91 | 16 | 75 |
| SPMS | 14 | 13 | 1 | |
| PPMS | 5 | 5 | - | |
| Age (years) | 48.4 ± 10.5 | 51.5 ± 11.3 | 47.0 ± 9.9 | |
| Disease Duration (years) | 14.0 ± 8.4 | 17.18 ± 9.34 | 12.6 ± 7.5 | |
| MS Severity [EDSS 0–10; median (range)] | 2.0 (0–7.0) | 6.0 (3.0–7.0) | 1.0 (0–2.5) | |
| Functional Tests | ||||
| Walking Speed | cm/s | 100.25 ± 28.73 | 74.36 ± 29.09 | 111.83 ± 19.68 |
| cm/s/heightcm | 0.59 ± 0.17 | 0.44 ± 0.17 | 0.66 ± 0.12 | |
| Upper Extremity [9HPT (seconds)] | Weaker | 24.42 ± 8.13 | 30.20 ± 11.04 | 22.52 ± 5.87 |
| Stronger | 22.27 ± 4.84 | 26.83 ± 6.17 | 20.71 ± 3.02 | |
| Fatigue [Low (0 mm) to High (100 mm)] | 39.5 ± 30.1 | 37.8 ± 11.0 | 40.2 ± 31.4 | |
| Cognitive Processing Speed (SDMT score) | 48.39 ± 10.82 | 41.21 ± 10.61 | 50.69 ± 9.90 | |
| All Participants (n = 110) | Higher Level of Disability (EDSS 4.9 ± 1.5, n = 34) | Lower Level of Disability (EDSS 1.2 ± 0.9, n = 76) | ||||
|---|---|---|---|---|---|---|
| TMS Variables | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand |
| RMT (MSO%) | 41 ± 11 (n = 99) | 43 ± 14 (n = 96) | 44 ± 14 (n = 29) | 50 ± 20 (n = 30) | 39 ± 9 (n = 70) | 40 ± 8 (n = 66) |
| AMT (MSO%) | 34 ± 8 (n = 103) | 37 ± 13 (n = 103) | 36 ± 10 (n = 30) | 44 ± 17 (n = 30) | 33 ± 7 (n = 73) | 34 ± 10 (n = 73) |
| Resting MEP Latency (ms; ms/heightcm) | 24.50 ± 3.70; 0.144 ± 0.022 (n = 96) | 25.11 ± 4.11; 0.148 ± 0.023 (n = 92) | 26.88 ± 5.63; 0.159 ± 0.34 (n = 29) | 27.45 ± 5.75; 0.160 ± 0.33 (n = 29) | 23.48 ± 1.6; 0.138 ± 0.01 (n = 67) | 24.03 ± 2.5; 0.141 ± 0.13 (n =60) |
| Active MEP Latency (ms; ms/heightcm) | 23.60 ± 3.41; 0.139 ± 0.019 (n = 101) | 24.20 ± 4.18; 0.143 ± 0.023 (n = 102) | 26.14 ± 4.82; 0.154 ± 0.28 (n = 30) | 26.89 ± 5.97; 0.158 ± 0.33 (n = 30) | 22.52 ± 1.7; 0.133 ± 0.01 (n = 71) | 23.02 ± 2.3; 0.136 ± 0.012 (n = 71) |
| MEP Amplitude 105% AMT (µV) | 366.47 ± 199.88 (n = 84) | 352.49 ± 184.99 (n = 82) | 313.95 ± 150.34 (n = 22) | 297.97 ± 97.99 (n = 21) | 385.11 ± 212.68 (n = 62) | 371.26 ± 203.96 (n = 61) |
| MEP Amplitude 115% AMT (µV) | 695.69 ± 504.93 (n = 84) | 626.97 ± 384.11 (n = 82) | 611.07 ± 530.65 (n = 22) | 437.40 ± 250.03 (n = 21) | 725.71 ± 496.46 (n = 62) | 692.23 ± 401.79 (n = 61) |
| MEP Amplitude 125% AMT (µV) | 1072.55 ± 697.87 (n = 84) | 920.43 ± 554.91 (n = 82) | 962.64 ± 788.99 (n = 22) | 623.63 ± 408.55 (n = 21) | 1111.55 ± 665.18 (n = 62) | 1022.61 ± 564.46 (n = 61) |
| MEP Amplitude 135% AMT (µV) | 1442.97 ± 870.71 (n = 84) | 1246.99 ± 773.34 (n = 80) | 1204.49 ± 944.31 (n = 22) | 787.22 ± 482.55 (n = 21) | 1527.60 ± 834.74 (n = 62) | 1410.63 ± 794.03 (n = 59) |
| MEP Amplitude 145% AMT (µV) | 1757.19 ± 985.46 (n = 84) | 1507.81 ± 941.55 (n = 80) | 1465.85 ± 1019.74 (n = 22) | 819.45 ± 556.27 (n = 21) | 1860.57 ± 960.16 (n = 62) | 1752.82 ± 931.75 (n = 59) |
| MEP Amplitude 155% AMT (µV) | 1986.81 ± 1034.14 (n = 84) | 1768.90 ± 1044.18 (n = 79) | 1713.77 ± 1112.91 (n = 22) | 1084.56 ± 883.46 (n = 20) | 2083.70 ± 996.15 (n = 62) | 2000.88 ± 997.18 (n = 59) |
| eREC AUC | 61,450.39 ± 33,185.46 (n = 84) | 54,323.52 ± 28,516.35 (n = 79) | 52,579.10 ± 35,594.71 (n = 22) | 34,558.14 ± 19,665.86 (n = 20) | 64,598.27 ± 31,997.28 (n = 62) | 61,023.65 ± 28,044.92 (n = 59) |
| eREC Slope | 33.31 ± 20.69 (n = 84) | 28.64 ± 21.35 (n = 79) | 28.02 ± 21.76 (n = 22) | 15.04 ± 15.69 (n = 20) | 35.18 ± 20.14 (n = 62) | 33.25 ± 21.14 (n = 59) |
| eREC R2 | 0.86 ± 0.10 (n = 84) | 0.77 ± 0.22 (n = 79) | 0.80 ± 0.13 (n = 22) | 0.61 ± 0.32 (n = 20) | 0.88 ± 0.8 (n = 62) | 0.82 ± 0.15 (n = 59) |
| CSP Duration 105% AMT (ms) | 61.34 ± 29.24 (n = 83) | 74.62 ± 43.96 (n = 80) | 75.13 ± 36.66 (n = 22) | 105.09 ± 61.64 (n = 21) | 56.37 ± 24.56 (n = 61) | 63.78 ± 29.44 (n = 59) |
| CSP Duration 115% AMT (ms) | 80.46 ± 37.93 (n = 82) | 90.97 ± 46.47 (n = 79) | 94.71 ± 42.92 (n = 22) | 119.98 ± 55.75 (n = 21) | 75.24 ± 34.88 (n = 60) | 80.46 ± 37.98 (n = 58) |
| CSP Duration 125% AMT (ms) | 97.16 ± 41.50 (n = 82) | 112.16 ± 52.39 (n = 79) | 117.24 ± 43.01 (n = 22) | 145.07 ± 63.00 (n = 21) | 89.80 ± 38.74 (n = 60) | 100.25 ± 42.65 (n = 58) |
| CSP Duration 135% AMT (ms) | 116.64 ± 44.03 (n = 82) | 126.03 ± 52.42 (n = 77) | 136.43 ± 43.82 (n = 22) | 158.89 ± 59.92 (n = 21) | 109.39 ± 42.17 (n = 60) | 113.71 ± 43.86 (n = 58) |
| CSP Duration 145% AMT (ms) | 130.82 ± 43.49 (n = 82) | 138.34 ± 54.00 (n = 75) | 151.15 ± 46.81 (n = 22) | 168.51 ± 70.46 (n = 20) | 123.37 ± 40.07 (n = 60) | 127.36 ± 42.30 (n = 58) |
| CSP Duration 155% AMT (ms) | 142.20 ± 44.42 (n = 81) | 148.45 ± 50.08 (n = 77) | 167.50 ± 45.83 (n = 22) | 175.39 ± 57.57 (n = 20) | 132.92 ± 40.43 (n = 60) | 139.00 ± 43.92 (n = 58) |
| iREC AUC | 5278.49 ± 1881.28 (n = 81) | 5738.80 ± 2266.99 (n = 75) | 6208.30 ± 2028.57 (n = 22) | 7150.18 ± 2727.68 (n = 20) | 4931.78 ± 1715.08 (n = 59) | 5225.58 ± 1848.99 (n = 55) |
| iSP Onset Latency (ms) | 35.83 ± 7.70 (n = 55) | 38.33 ± 9.19 (n = 55) | 41.47 ± 9.25 (n = 15) | 45.60 ± 10.13 (n = 15) | 33.71 ± 5.88 (n = 40) | 35.61 ± 7.21 (n = 40) |
| iSP Duration (ms) | 30.10 ± 12.67 (n = 55) | 26.37 ± 13.36 (n = 55) | 36.42 ± 15.53 (n = 15) | 25.74 ± 15.85 (n = 15) | 27.73 ± 10.71 (n = 40) | 26.61 ± 12.52 (n = 40) |
| iSP AUC | 49.50 ± 12.48 (n = 55) | 47.62 ± 14.81 (n = 55) | 42.91 ± 11.31 (n = 15) | 40.79 ± 18.66 (n = 15) | 51.97 ± 12.11 (n = 40) | 50.18 ± 12.41 (n = 40) |
| iSP Depth | 50.77 ± 11.73 (n = 55) | 53.46 ± 12.02 (n = 55) | 55.60 ± 13.03 (n = 15) | 58.11 ± 14.32 (n = 15) | 48.97 ± 10.83 (n = 40) | 51.71 ± 10.72 (n = 40) |
| iSP Depth × Duration | 1484.66 ± 632.71 (n = 55) | 1363.79 ± 723.69 (n = 55) | 1897.03 ± 639.95 (n = 15) | 1442.37 ± 940.06 (n = 15) | 1330.03 ± 563.33 (n = 40) | 1334.32 ± 636.13 (n = 40) |
| Statistically significant findings (sig.) are highlighted as: | Sig. ≤ 0.001 | Sig. ≤ 0.010 | Sig. ≤ 0.050 | |||
| Walking Speed | 9HPT Stronger Hand | 9HPT Weaker Hand | Fatigue | SDMT | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TMS Variables | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand | Stronger Hand | Weaker Hand |
| RMT (MSO%) | r = 0.138 | r = 0.286 | r = 0.247 | r = 0.333 | r = 0.212 | r = 0.333 | r = 0.048 | r = 0.179 | r = 0.167 | r = 0.358 |
| AMT (MSO%) | r = 0.211 | r = 0.275 | r = 0.146 | r = 0.340 | r = 0.203 | r = 0.391 | r = 0.076 | r = 0.161 | r = 0.095 | r = 0.272 |
| RMT Asymmetry | r = 0.214 | r = 0.115 | r = 0.183 | r = 0.204 | r = 0.253 | |||||
| AMT Asymmetry | r = 0.274 | r = 0.290 | r = 0.329 | r = 0.284 | r = 0.311 | |||||
| Resting MEP Latency (ms; ms/heightcm) | r = 0.234 | r = 0.314 | r = 0.242 | r = 0.463 | r = 0.309 | r = 0.422 | r = 0.033 | r = 0.092 | r = 0.160 | r = 0.217 |
| Active MEP Latency (ms; ms/heightcm) | r = 0.424 | r = 0.504 | r = 0.401 | r = 0.421 | r = 0.390 | r = 0.449 | r = 0.097 | r = 0.050 | r = 0.280 | r = 0.342 |
| MEP Amplitude 105% AMT (µV) | r = 0.172 | r = 0.146 | r = 0.149 | r = 0.037 | r = 0.243 | r = 0.048 | r = 0.019 | r = 0.220 | r = 0.207 | r = 0.109 |
| MEP Amplitude 115% AMT (µV) | r = 0.268 | r = 0.157 | r = 0.080 | r = 0.291 | r = 0.183 | r = 0.294 | r = 0.069 | r < 0.001 | r = 0.197 | r = 0.272 |
| MEP Amplitude 125% AMT (µV) | r = 0.352 | r = 0.228 | r = 0.121 | r = 0.243 | r = 0.190 | r = 0.333 | r = 0.012 | r = 0.010 | r = 0.188 | r = 0.252 |
| MEP Amplitude 135% AMT (µV) | r = 0.379 | r = 0.332 | r = 0.233 | r = 0.331 | r = 0.342 | r = 0.405 | r = 0.014 | r = 0.109 | r = 0.268 | r = 0.324 |
| MEP Amplitude 145% AMT (µV) | r = 0.259 | r = 0.338 | r = 0.214 | r = 0.300 | r = 0.292 | r = 0.402 | r = 0.066 | r = 0.019 | r = 0.287 | r = 0.371 |
| MEP Amplitude 155% AMT (µV) | r = 0.264 | r = 0.286 | r = 0.200 | r = 0.390 | r = 0.304 | r = 0.369 | r = 0.006 | r = 0.117 | r = 0.255 | r = 0.388 |
| eREC AUC | r = 0.360 | r = 0.317 | r = 0.221 | r = 0.388 | r = 0.328 | r = 0.403 | r = 0.030 | r = 0.067 | r = 0.300 | r = 0.367 |
| eREC Slope | r = 0.246 | r = 0.246 | r = 0.211 | r = 0.388 | r = 0.274 | r = 0.399 | r = 0.031 | r = 0.082 | r = 0.240 | r = 0.384 |
| eREC R2 | r = 0.116 | r = 0.344 | r = 0.049 | r = 0.185 | r = 0.004 | r = 0.242 | r = 0.033 | r = 0.047 | r = 0.148 | r = 0.204 |
| CSP Duration 105% AMT (ms) | r = 0.298 | r = 0.282 | r = 0.120 | r = 0.324 | r = 0.262 | r = 0.396 | r = 0.113 | r = 0.176 | r = 0.166 | r = 0.284 |
| CSP Duration 115% AMT (ms) | r = 0.259 | r = 0.364 | r = 0.136 | r = 0.288 | r = 0.222 | r = 0.287 | r = 0.070 | r = 0.135 | r = 0.111 | r = 0.20 |
| CSP Duration 125% AMT (ms) | r = 0.273 | r = 0.235 | r = 0.116 | r = 0.235 | r = 0.267 | r = 0.295 | r = 0.087 | r = 0.186 | r = 0.098 | r = 0.209 |
| CSP Duration 135% AMT (ms) | r = 0.297 | r = 0.234 | r = 0.015 | r = 0.213 | r = 0.159 | r = 0.272 | r = 0.084 | r = 0.244 | r = 0.008 | r = 0.176 |
| CSP Duration 145% AMT (ms) | r = 0.362 | r = 0.248 | r = 0.060 | r = 0.127 | r = 0.221 | r = 0.215 | r = 0.142 | r = 0.251 | r = 0.039 | r = 0.058 |
| CSP Duration 155% AMT (ms) | r = 0.324 | r = 0.239 | r = 0.051 | r = 0.121 | r = 0.222 | r = 0.244 | r = 0.201 | r = 0.340 | r = 0.001 | r = 0.021 |
| iREC AUC | r = 0.313 | r = 0.279 | r = 0.088 | r = 0.198 | r = 0.228 | r = 0.260 | r = 0.104 | r = 0.229 | r = 0.056 | r = 0.013 |
| iSP Onset Latency (ms) | r = 0.552 | r = 0.526 | r = 0.237 | r = 0.506 | r = 0.425 | r = 0.471 | r = 0.004 | r = 0.009 | r = 0.194 | r = 0.290 |
| iSP Duration (ms) | r = 0.330 | r = 0.082 | r = 0.169 | r = 0.065 | r = 0.111 | r = 0.248 | r = 0.077 | r = 0.022 | r = 0.058 | r = 0.062 |
| iSP AUC | r = 0.356 | r = 0.281 | r = 0.289 | r = 0.065 | r = 0.296 | r = 0.173 | r = 0.030 | r = 0.158 | r = 0.034 | r = 0.164 |
| iSP Depth | r = 0.263 | r = 0.270 | r = 0.219 | r = 0.272 | r = 0.270 | r = 0.288 | r = 0.039 | r = 0.215 | r = 0.267 | r = 0.297 |
| iSP Depth×Duration | r = 0.451 | r = 0.014 | r = 0.248 | r = 0.192 | r = 0.220 | r = 0.368 | r = 0.076 | r = 0.135 | r = 0.187 | r = 0.201 |
| Statistically significant findings (sig.) are highlighted as: | Sig. ≤ 0.001 | Sig. ≤ 0.010 | Sig. ≤ 0.050 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, A.R.; Snow, N.J.; Alcock, L.R.; Ploughman, M. Probing the Brain–Body Connection Using Transcranial Magnetic Stimulation (TMS): Validating a Promising Tool to Provide Biomarkers of Neuroplasticity and Central Nervous System Function. Brain Sci. 2021, 11, 384. https://doi.org/10.3390/brainsci11030384
Chaves AR, Snow NJ, Alcock LR, Ploughman M. Probing the Brain–Body Connection Using Transcranial Magnetic Stimulation (TMS): Validating a Promising Tool to Provide Biomarkers of Neuroplasticity and Central Nervous System Function. Brain Sciences. 2021; 11(3):384. https://doi.org/10.3390/brainsci11030384
Chicago/Turabian StyleChaves, Arthur R., Nicholas J. Snow, Lynsey R. Alcock, and Michelle Ploughman. 2021. "Probing the Brain–Body Connection Using Transcranial Magnetic Stimulation (TMS): Validating a Promising Tool to Provide Biomarkers of Neuroplasticity and Central Nervous System Function" Brain Sciences 11, no. 3: 384. https://doi.org/10.3390/brainsci11030384
APA StyleChaves, A. R., Snow, N. J., Alcock, L. R., & Ploughman, M. (2021). Probing the Brain–Body Connection Using Transcranial Magnetic Stimulation (TMS): Validating a Promising Tool to Provide Biomarkers of Neuroplasticity and Central Nervous System Function. Brain Sciences, 11(3), 384. https://doi.org/10.3390/brainsci11030384