

Increased Interpersonal Brain Synchronization in Romantic Couples Is Associated with Higher Honesty: An fNIRS Hyperscanning Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Task and Procedure
2.3. fNIRS Data Acquisition
2.4. Data Analysis
2.4.1. Behavior Performance
2.4.2. Interpersonal Brain Synchronization
2.4.3. Directional Coupling
3. Results
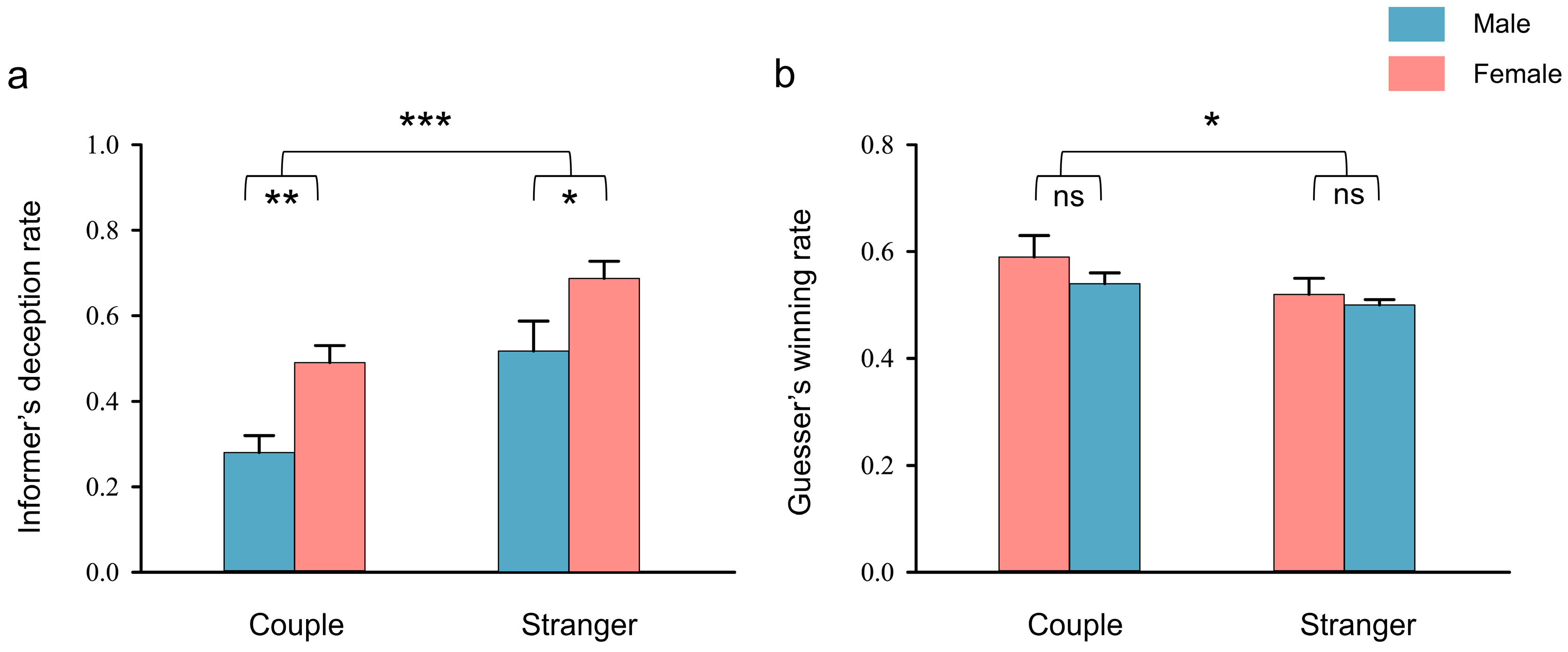
3.1. Behavioral Performance
3.2. Interbrain Synchronization
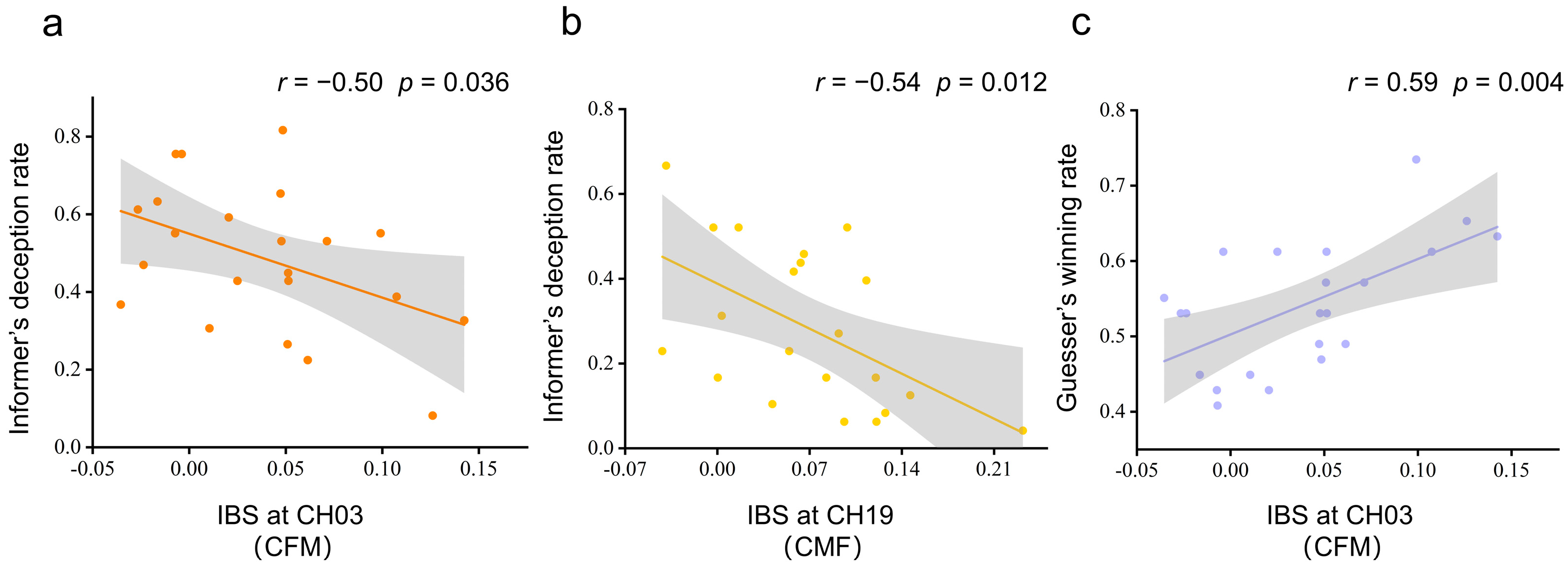
3.3. Interbrain Synchronization and Behavior Performance
3.4. Directional Coupling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DePaulo, B.M.; Lindsay, J.J.; Malone, B.E.; Muhlenbruck, L.; Charlton, K.; Cooper, H. Cues to deception. Psychol. Bull. 2003, 129, 74–118. [Google Scholar] [CrossRef]
- DePaulo, B.M.; Kashy, D.A.; Kirkendol, S.E.; Wyer, M.M.; Epstein, J.A. Lying in everyday life. J. Personal. Soc. Psychol. 1996, 70, 979–995. [Google Scholar] [CrossRef]
- Holm, H.J.; Kawagoe, T. Face-to-face lying—An experimental study in Sweden and Japan. J. Econ. Psychol. 2010, 31, 310–321. [Google Scholar] [CrossRef]
- Buroon, J.K.; Bulkr, D.B.; Ebesu, A.S.; White, C.H.; Rockwell, P.A. Testing Interpersonal Deception Theory: Effects of Suspicion on Communication Behaviors and Perceptions. Commun. Theory 2006, 6, 243–267. [Google Scholar] [CrossRef]
- Pinti, P.; Devoto, A.; Greenhalgh, I.; Tachtsidis, I.; Burgess, P.W.; Hamilton, A.F.d.C. The role of anterior prefrontal cortex (area 10) in face-to-face deception measured with fNIRS. Soc. Cogn. Affect. Neurosci. 2021, 16, 129–142. [Google Scholar] [CrossRef]
- Dreber, A.; Johannesson, M. Gender differences in deception. Econ. Lett. 2008, 99, 197–199. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Kray, L.J. Gender similarities and differences in dishonesty. Curr. Opin. Psychol. 2022, 48, 101461. [Google Scholar] [CrossRef]
- Vasconcellos, S.J.L.; Rizzatti, M.; Barbosa, T.P.; Schmitz, B.S.; Coelho, V.C.N.; Machado, A. Understanding Lies Based on Evolutionary Psychology: A Critical Review. Trends Psychol. 2019, 27, 141–153. [Google Scholar] [CrossRef]
- Buss, D.M.; Schmitt, D.P. Sexual strategies theory: An evolutionary perspective on human mating. Psychol. Rev. 1993, 100, 204–232. [Google Scholar] [CrossRef]
- de Vries, R.E.; Zettler, I.; Hilbig, B.E. Rethinking Trait Conceptions of Social Desirability Scales: Impression Management as an Expression of Honesty-Humility. Assessment 2014, 21, 286–299. [Google Scholar] [CrossRef]
- Mogilski, J.K.; Welling, L.L.M. Staying friends with an ex: Sex and dark personality traits predict motivations for post-relationship friendship. Personal. Individ. Differ. 2017, 115, 114–119. [Google Scholar] [CrossRef]
- Sutter, M.; Bosman, R.; Kocher, M.G.; van Winden, F. Gender pairing and bargaining—Beware the same sex! Exp. Econ. 2009, 12, 318–331. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, T.; Zhang, R.; Wang, N.; Yin, Q.; Li, Y.; Liu, J.; Liu, T.; Li, X. Neural alignment during face-to-face spontaneous deception: Does gender make a difference? Hum. Brain Mapp. 2020, 41, 4964–4981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, T.; Pelowski, M.; Yu, D. Gender difference in spontaneous deception: A hyperscanning study using functional near-infrared spectroscopy. Sci. Rep. 2017, 7, 7508. [Google Scholar] [CrossRef] [PubMed]
- Ennis, E.; Vrij, A.; Chance, C. Individual differences and lying in everyday life. J. Soc. Pers. Relatsh. 2008, 25, 105–118. [Google Scholar] [CrossRef]
- Le, B.M.; Chopik, W.J.; Shimshock, C.J.; Chee, P.X. When the truth helps and when it hurts: How honesty shapes well-being. Curr. Opin. Psychol. 2022, 46, 101397. [Google Scholar] [CrossRef] [PubMed]
- Lemay, E.P., Jr.; Bechis, M.A.; Martin, J.; Neal, A.M.; Coyne, C. Concealing negative evaluations of a romantic partner’s physical attractiveness. Pers. Relatsh. 2013, 20, 669–689. [Google Scholar] [CrossRef]
- Cote, M.; Dufour, M.; Tremblay, J. Integrative couple treatment for pathological gamblers with an emphasis on forgiveness processes: A case study with three couples. J. Marital Fam. Ther. 2022, 48, 1017–1039. [Google Scholar] [CrossRef]
- Haselton, M.G.; Buss, D.M.; Oubaid, V.; Angleitner, A. Sex, lies, and strategic interference: The psychology of deception between the sexes. Personal. Soc. Psychol. Bull. 2005, 31, 3–23. [Google Scholar] [CrossRef]
- Weiser, D.A.; Shrout, M.R.; Thomas, A.V.; Edwards, A.L.; Pickens, J.C. “I’ve been cheated, been mistreated, when will I be loved”: Two decades of infidelity research through an intersectional lens. J. Soc. Pers. Relatsh. 2022, 40, 856–898. [Google Scholar] [CrossRef]
- Wickham, R.E. Perceived authenticity in romantic partners. J. Exp. Soc. Psychol. 2013, 49, 878–887. [Google Scholar] [CrossRef]
- Feldman, R. The Neurobiology of Human Attachments. Trends Cogn. Sci. 2017, 21, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S. Oxytocin Pathways and the Evolution of Human Behavior. Annu. Rev. Psychol. 2014, 65, 17–39. [Google Scholar] [CrossRef]
- Goldstein, P.; Weissman-Fogel, I.; Dumas, G.; Shamay-Tsoory, S.G. Brain-to-brain coupling during handholding is associated with pain reduction. Proc. Natl. Acad. Sci. USA 2018, 115, E2528–E2537. [Google Scholar] [CrossRef]
- Long, Y.; Zheng, L.; Zhao, H.; Zhou, S.; Zhai, Y.; Lu, C. Interpersonal Neural Synchronization during Interpersonal Touch Underlies Affiliative Pair Bonding between Romantic Couples. Cereb. Cortex 2021, 31, 1647–1659. [Google Scholar] [CrossRef]
- Kinreich, S.; Djalovski, A.; Kraus, L.; Louzoun, Y.; Feldman, R. Brain-to-Brain Synchrony during Naturalistic Social Interactions. Sci. Rep. 2017, 7, 17060. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Chen, C.; Wu, K.; Zhou, S.; Zhou, F.; Zheng, L.; Zhao, H.; Zhai, Y.; Lu, C. Interpersonal Conflict Increases Interpersonal Neural Synchronization in Romantic Couples. Cereb. Cortex 2022, 32, 3254–3268. [Google Scholar] [CrossRef]
- Duan, H.; Yang, T.; Wang, X.; Kan, Y.; Zhao, H.; Li, Y.; Hu, W. Is the creativity of lovers better? A behavioral and functional near-infrared spectroscopy hyperscanning study. Curr. Psychol. 2022, 41, 41–54. [Google Scholar] [CrossRef]
- Pan, Y.; Cheng, X.; Zhang, Z.; Li, X.; Hu, Y. Cooperation in lovers: An fNIRS-based hyperscanning study. Hum. Brain Mapp. 2017, 38, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.R.; Mongeau, P.A.; Sleight, C. Fudging with Friends and Lying to Lovers: Deceptive Communication in Personal Relationships. J. Soc. Pers. Relatsh. 1986, 3, 495–512. [Google Scholar] [CrossRef]
- Lee, T.M.C.; Lee, T.M.Y.; Raine, A.; Chan, C.C.H. Lying about the Valence of Affective Pictures: An fMRI Study. PLoS ONE 2010, 5, e12291. [Google Scholar] [CrossRef] [PubMed]
- Gneezy, U. Deception: The Role of Consequences. Am. Econ. Rev. 2005, 95, 384–394. [Google Scholar] [CrossRef]
- Ding, X.P.; Gao, X.; Fu, G.; Lee, K. Neural correlates of spontaneous deception: A functional near-infrared spectroscopy (fNIRS) study. Neuropsychologia 2013, 51, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.P.; Sai, L.; Fu, G.; Liu, J.; Lee, K. Neural correlates of second-order verbal deception: A functional near-infrared spectroscopy (fNIRS) study. NeuroImage 2014, 87, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Zhang, S.; Jin, T.; Wu, H.; Su, S.; Liu, C. Brain activation and adaptation of deception processing during dyadic face-to-face interaction. Cortex 2019, 120, 326–339. [Google Scholar] [CrossRef]
- Abe, N.; Suzuki, M.; Tsukiura, T.; Mori, E.; Yamaguchi, K.; Itoh, M.; Fujii, T. Dissociable roles of prefrontal and anterior cingulate cortices in deception. Cereb. Cortex 2006, 16, 192–199. [Google Scholar] [CrossRef]
- Jenkins, A.C.; Zhu, L.; Hsu, M. Cognitive neuroscience of honesty and deception: A signaling framework. Curr. Opin. Behav. Sci. 2016, 11, 130–137. [Google Scholar] [CrossRef]
- Bhatt, M.A.; Lohrenz, T.; Camerer, C.F.; Montague, P.R. Neural signatures of strategic types in a two-person bargaining game. Proc. Natl. Acad. Sci. USA 2010, 107, 19720–19725. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Zhou, X.; Yu, R. Interpersonal brain synchronization under bluffing in strategic games. Soc. Cogn. Affect. Neurosci. 2020, 15, 1315–1324. [Google Scholar] [CrossRef]
- Hatfield, E.; Sprecher, S. Measuring passionate love in intimate relationships. J. Adolesc. 1986, 9, 383–410. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, X.; Feng, D.; Yuan, D.; Li, S.; Lu, C.; Li, X. Effects of acute psychosocial stress on interpersonal cooperation and competition in young women. Brain Cogn. 2021, 151, 105738. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Qiao, X.; Yun, Q.; Hao, N. Educational diversity and group creativity: Evidence from fNIRS hyperscanning. NeuroImage 2021, 243, 118564. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Xiao, W.; Lin, S.; Shao, Y.; Li, W.; Zhang, W. Cooperation with partners of differing social experience: An fNIRS-based hyperscanning study. Brain Cogn. 2021, 154, 105803. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y. Functional near-infrared spectroscopy: Current status and future prospects. J. Biomed. Opt. 2007, 12, 062106. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Bryant, D.M.; Reiss, A.L. NIRS-based hyperscanning reveals increased interpersonal coherence in superior frontal cortex during cooperation. NeuroImage 2012, 59, 2430–2437. [Google Scholar] [CrossRef]
- Tn, A.; Hsbc, D.; Eke, F.; Dm, F.; Pv, F.; Sha, F. The effects of interaction quality on neural synchrony during mother-child problem solving. Cortex 2020, 124, 235–249. [Google Scholar]
- Lu, K.; Yu, T.; Hao, N. Creating while taking turns, the choice to unlocking group creative potential. NeuroImage 2020, 219, 117025. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, Z.; Zhao, C.; Duan, L.; Gong, Y.; Li, Z.; Zhu, C. NIRS-KIT: A MATLAB toolbox for both resting-state and task fNIRS data analysis. Neurophotonics 2021, 8, 010802. [Google Scholar] [CrossRef]
- Cui, X.; Bray, S.; Reiss, A.L. Functional near infrared spectroscopy (NIRS) signal improvement based on negative correlation between oxygenated and deoxygenated hemoglobin dynamics. NeuroImage 2010, 49, 3039–3046. [Google Scholar] [CrossRef]
- Emberson, L.L.; Crosswhite, S.L.; Goodwin, J.R.; Berger, A.J.; Aslin, R.N. Isolating the effects of surface vasculature in infant neuroimaging using short-distance optical channels: A combination of local and global effects. Neurophotonics 2016, 3, 031406. [Google Scholar] [CrossRef]
- Grinsted, A.; Moore, J.C.; Jevrejeva, S. Application of the cross wavelet transform and wavelet coherence to geophysical time series. Nonlinear Process. Geophys. 2004, 11, 561–566. [Google Scholar] [CrossRef]
- Pan, Y.; Dikker, S.; Goldstein, P.; Zhu, Y.; Yang, C.; Hu, Y. Instructor-learner brain coupling discriminates between instructional approaches and predicts learning. NeuroImage 2020, 211, 116657. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Y.; Wang, X.; Kan, Y.; Xu, S.; Duan, H. Inter-brain neural mechanism underlying turn-based interaction under acute stress in women: A hyperscanning study using functional near-infrared spectroscopy. Soc. Cogn. Affect. Neurosci. 2022, 17, 850–863. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Lu, K.; Hao, N. Cooperation makes two less-creative individuals turn into a highly-creative pair. NeuroImage 2018, 172, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, T.; Sasaki, Y.; Sakaki, K.; Yokoyama, R.; Kawashima, R. Interpersonal frontopolar neural synchronization in group communication: An exploration toward fNIRS hyperscanning of natural interactions. NeuroImage 2016, 133, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A Network Visualization Tool for Human Brain Connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef]
- Liang, Z.; Li, S.; Zhou, S.; Chen, S.; Li, Y.; Chen, Y.; Zhao, Q.; Huang, F.; Lu, C.; Yu, Q.; et al. Increased or decreased? Interpersonal neural synchronization in group creation. NeuroImage 2022, 260, 119448. [Google Scholar] [CrossRef]
- Ono, Y.; Zhang, X.; Noah, J.A.; Dravida, S.; Hirsch, J. Bidirectional Connectivity Between Broca’s Area and Wernicke’s Area During Interactive Verbal Communication. Brain Connect. 2022, 12, 210–222. [Google Scholar] [CrossRef]
- Pan, Y.; Novembre, G.; Song, B.; Li, X.; Hu, Y. Interpersonal synchronization of inferior frontal cortices tracks social interactive learning of a song. NeuroImage 2018, 183, 280–290. [Google Scholar] [CrossRef]
- Niso, G.; Bruna, R.; Pereda, E.; Gutierrez, R.; Bajo, R.; Maestu, F.; del-Pozo, F. HERMES: Towards an Integrated Toolbox to Characterize Functional and Effective Brain Connectivity. Neuroinformatics 2013, 11, 405–434. [Google Scholar] [CrossRef]
- Capraro, V. Gender differences in lying in sender-receiver games: A meta-analysis. Judgm. Decis. Mak. 2018, 13, 345–355. [Google Scholar] [CrossRef]
- Lee, M.; Pitesa, M.; Pillutla, M.M.; Thau, S. Male immorality: An evolutionary account of sex differences in unethical negotiation behavior. Acad. Manag. J. 2017, 60, 2014–2044. [Google Scholar] [CrossRef]
- Buss, D.M.; Barnes, M. Preferences in human mate selection. J. Personal. Soc. Psychol. 1986, 50, 559–570. [Google Scholar] [CrossRef]
- Lippa, R.A. The preferred traits of mates in a cross-national study of heterosexual and homosexual men and women: An examination of biological and cultural influences. Arch. Sex. Behav. 2007, 36, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.; Marshall, T.; Fueloep, M.; Adonu, J.; Spiewak, S.; Neto, F.; Plaza, S.H. Mate Value and Self-Esteem: Evidence from Eight Cultural Groups. PLoS ONE 2012, 7, e36106. [Google Scholar] [CrossRef]
- Holden, C.J.; Zeigler-Hill, V.; Pham, M.N.; Shackelford, T.K. Personality features and mate retention strategies: Honesty-humility and the willingness to manipulate, deceive, and exploit romantic partners. Personal. Individ. Differ. 2014, 57, 31–36. [Google Scholar] [CrossRef]
- Weber, M.; Ruch, W. The role of character strengths in adolescent romantic relationships: An initial study on partner selection and mates’ life satisfaction. J. Adolesc. 2012, 35, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Garbinsky, E.N.; Gladstone, J.J.; Nikolova, H.; Olson, J.G. Love, Lies, and Money: Financial Infidelity in Romantic Relationships. J. Consum. Res. 2020, 47, 1–24. [Google Scholar] [CrossRef]
- Britt, S.; Grable, J.E.; Goff, B.S.N.; White, M. The influence of perceived spending behaviors on relationship satisfaction. Financ. Couns. Plan. 2008, 19, 31–43. [Google Scholar]
- Gao, M.; Yang, X.; Shi, J.; Lin, Y.; Chen, S. Does Gender Make a Difference in Deception? The Effect of Transcranial Direct Current Stimulation Over Dorsolateral Prefrontal Cortex. Front. Psychol. 2018, 9, 1321. [Google Scholar] [CrossRef]
- Lucas, I.; Balada, F.; Blanco, E.; Aluja, A. Prefrontal cortex activity triggered by affective faces exposure and its relationship with neuroticism. Neuropsychologia 2019, 132, 107146. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, A.; Fellows, L.K. Are You Upset? Distinct Roles for Orbitofrontal and Lateral Prefrontal Cortex in Detecting and Distinguishing Facial Expressions of Emotion. Cereb. Cortex 2012, 22, 2904–2912. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, J.; Stolarski, M.; Matthews, G. Emotional Intelligence and Mismatching Expressive and Verbal Messages: A Contribution to Detection of Deception. PLoS ONE 2014, 9, e92570. [Google Scholar] [CrossRef] [PubMed]
- Ueda, R.; Yanagisawa, K.; Ashida, H.; Abe, N. Executive control and faithfulness: Only long-term romantic relationships require prefrontal control. Exp. Brain Res. 2018, 236, 821–828. [Google Scholar] [CrossRef]
- Dogan, A.; Morishima, Y.; Heise, F.; Tanner, C.; Gibson, R.; Wagner, A.F.; Tobler, P.N. Prefrontal connections express individual differences in intrinsic resistance to trading off honesty values against economic benefits. Sci. Rep. 2016, 6, 33263. [Google Scholar] [CrossRef]
- Bellucci, G.; Molter, F.; Park, S.Q. Neural representations of honesty predict future trust behavior. Nat. Commun. 2019, 10, 5184. [Google Scholar] [CrossRef]
- Frith, C.D.; Frith, U. Interacting minds—A biological basis. Science 1999, 286, 1692–1695. [Google Scholar] [CrossRef]
- Vetter, N.C.; Altgassen, M.; Phillips, L.; Mahy, C.E.V.; Kliegel, M. Development of Affective Theory of Mind Across Adolescence: Disentangling the Role of Executive Functions. Develop. Neuropsychol. 2013, 38, 114–125. [Google Scholar] [CrossRef]
- Bahnemann, M.; Dziobek, I.; Prehn, K.; Wolf, I.; Heekeren, H.R. Sociotopy in the temporoparietal cortex: Common versus distinct processes. Soc. Cogn. Affect. Neurosci. 2010, 5, 48–58. [Google Scholar] [CrossRef]
- Bio, B.J.; Guterstam, A.; Pinsk, M.; Wilterson, A.I.; Graziano, M.S.A. Right temporoparietal junction encodes inferred visual knowledge of others. Neuropsychologia 2022, 171, 108243. [Google Scholar] [CrossRef]
- Ciaramidaro, A.; Adenzato, M.; Enrici, I.; Erk, S.; Pia, L.; Bara, B.G.; Walter, H. The intentional network: How the brain reads varieties of intentions. Neuropsychologia 2007, 45, 3105–3113. [Google Scholar] [CrossRef] [PubMed]
- Kana, R.K.; Libero, L.E.; Hu, C.P.; Deshpande, H.D.; Colburn, J.S. Functional Brain Networks and White Matter Underlying Theory-of-Mind in Autism. Soc. Cogn. Affect. Neurosci. 2014, 9, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Dodell-Feder, D.; Felix, S.; Yung, M.G.; Hooker, C.I. Theory-of-mind-related neural activity for one’s romantic partner predicts partner well-being. Soc. Cogn. Affect. Neurosci. 2016, 11, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Esmenio, S.; Soares, J.M.; Oliveira-Silva, P.; Goncalves, O.F.; Friston, K.; Fernandes Coutinho, J. Changes in the Effective Connectivity of the Social Brain When Making Inferences About Close Others vs. the Self. Front. Hum. Neurosci. 2020, 14, 151. [Google Scholar] [CrossRef]
- Zhang, W.H.; Qiu, L.T.; Tang, F.G.; Li, H. Affective or cognitive interpersonal emotion regulation in couples: An fNIRS hyperscanning study. Cereb. Cortex 2023, bhad091. [Google Scholar] [CrossRef]
- Yuan, D.; Zhang, R.; Liu, J.; Feng, D.; Hu, Y.; Li, X.; Wang, Y.; Zhou, X. Interpersonal neural synchronization could predict the outcome of mate choice. Neuropsychologia 2022, 165, 108112. [Google Scholar] [CrossRef]
- Li, L.; Huang, X.; Xiao, J.; Zheng, Q.; Shan, X.; He, C.; Liao, W.; Chen, H.; Menon, V.; Duan, X. Neural synchronization predicts marital satisfaction. Proc. Natl. Acad. Sci. USA 2022, 119, e2202515119. [Google Scholar] [CrossRef]
- Schippers, M.B.; Roebroeck, A.; Renken, R.; Nanetti, L.; Keysers, C. Mapping the information flow from one brain to another during gestural communication. Proc. Natl. Acad. Sci. USA 2010, 107, 9388–9393. [Google Scholar] [CrossRef]
- Chen, L.; Qu, Y.Z.; Cao, J.Y.; Liu, T.Y.; Gong, Y.L.; Tian, Z.L.; Xiong, J.; Lin, Z.F.; Yang, X.; Yin, T.; et al. The increased inter-brain neural synchronization in prefrontal cortex between simulated patient and acupuncturist during acupuncture stimulation: Evidence from functional near-infrared spectroscopy hyperscanning. Hum. Brain Mapp. 2023, 44, 980–988. [Google Scholar] [CrossRef]
- Cheng, X.J.; Pan, Y.F.; Hu, Y.Y.; Hu, Y. Coordination Elicits Synchronous Brain Activity Between Co-actors: Frequency Ratio Matters. Front. Neurosci. 2019, 13, 1071. [Google Scholar] [CrossRef]
- Kumar, A.; Chowdhary, S.; Capraro, V.; Perc, M. Evolution of honesty in higher-order social networks. Phys. Rev. E 2021, 104, 054308. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.J.; Weber, B. Can beneficial ends justify lying? Neural responses to the passive reception of lies and truth-telling with beneficial and harmful monetary outcomes. Soc. Cogn. Affect. Neurosci. 2016, 11, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, P.; Teodorescu, K.; Hertwig, R. The Truth About Lies: A Meta-Analysis on Dishonest Behavior. Psychol. Bull. 2019, 145, 1–44. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, C.; Zhang, X.; Wu, Y.; Zhang, W.; Sun, B. Increased Interpersonal Brain Synchronization in Romantic Couples Is Associated with Higher Honesty: An fNIRS Hyperscanning Study. Brain Sci. 2023, 13, 833. https://doi.org/10.3390/brainsci13050833
Shao C, Zhang X, Wu Y, Zhang W, Sun B. Increased Interpersonal Brain Synchronization in Romantic Couples Is Associated with Higher Honesty: An fNIRS Hyperscanning Study. Brain Sciences. 2023; 13(5):833. https://doi.org/10.3390/brainsci13050833
Chicago/Turabian StyleShao, Chong, Xuecheng Zhang, You Wu, Wenhai Zhang, and Binghai Sun. 2023. "Increased Interpersonal Brain Synchronization in Romantic Couples Is Associated with Higher Honesty: An fNIRS Hyperscanning Study" Brain Sciences 13, no. 5: 833. https://doi.org/10.3390/brainsci13050833
APA StyleShao, C., Zhang, X., Wu, Y., Zhang, W., & Sun, B. (2023). Increased Interpersonal Brain Synchronization in Romantic Couples Is Associated with Higher Honesty: An fNIRS Hyperscanning Study. Brain Sciences, 13(5), 833. https://doi.org/10.3390/brainsci13050833