
Simultaneous fMRI and tDCS for Enhancing Training of Flight Tasks


Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Procedure
2.3. Flight Landing Task
2.4. Auditory Task
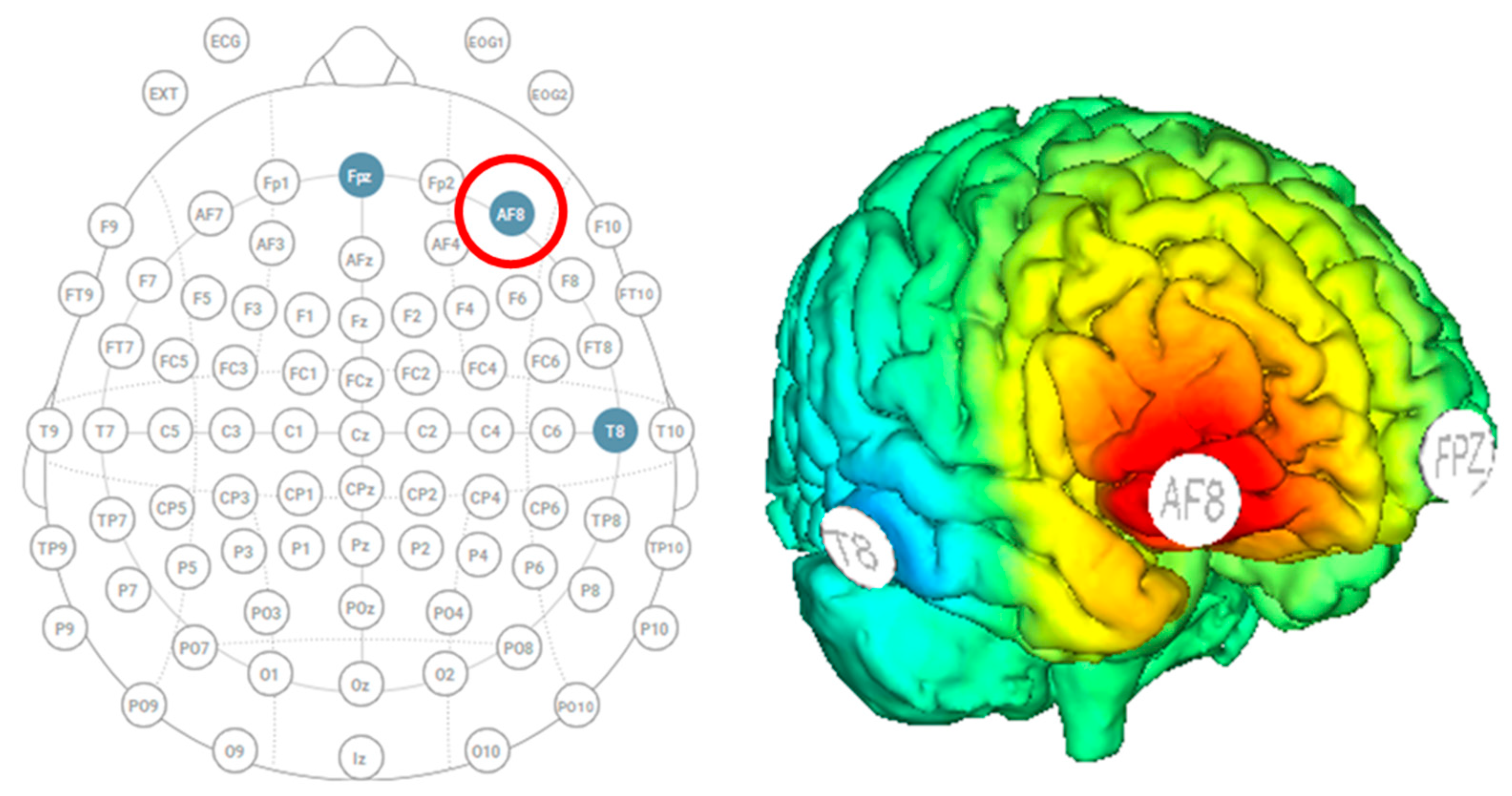
2.5. tDCS Neurostimulation
2.6. fMRI Neuroimaging
2.6.1. Scanning
2.6.2. Preprocessing
2.6.3. Analysis
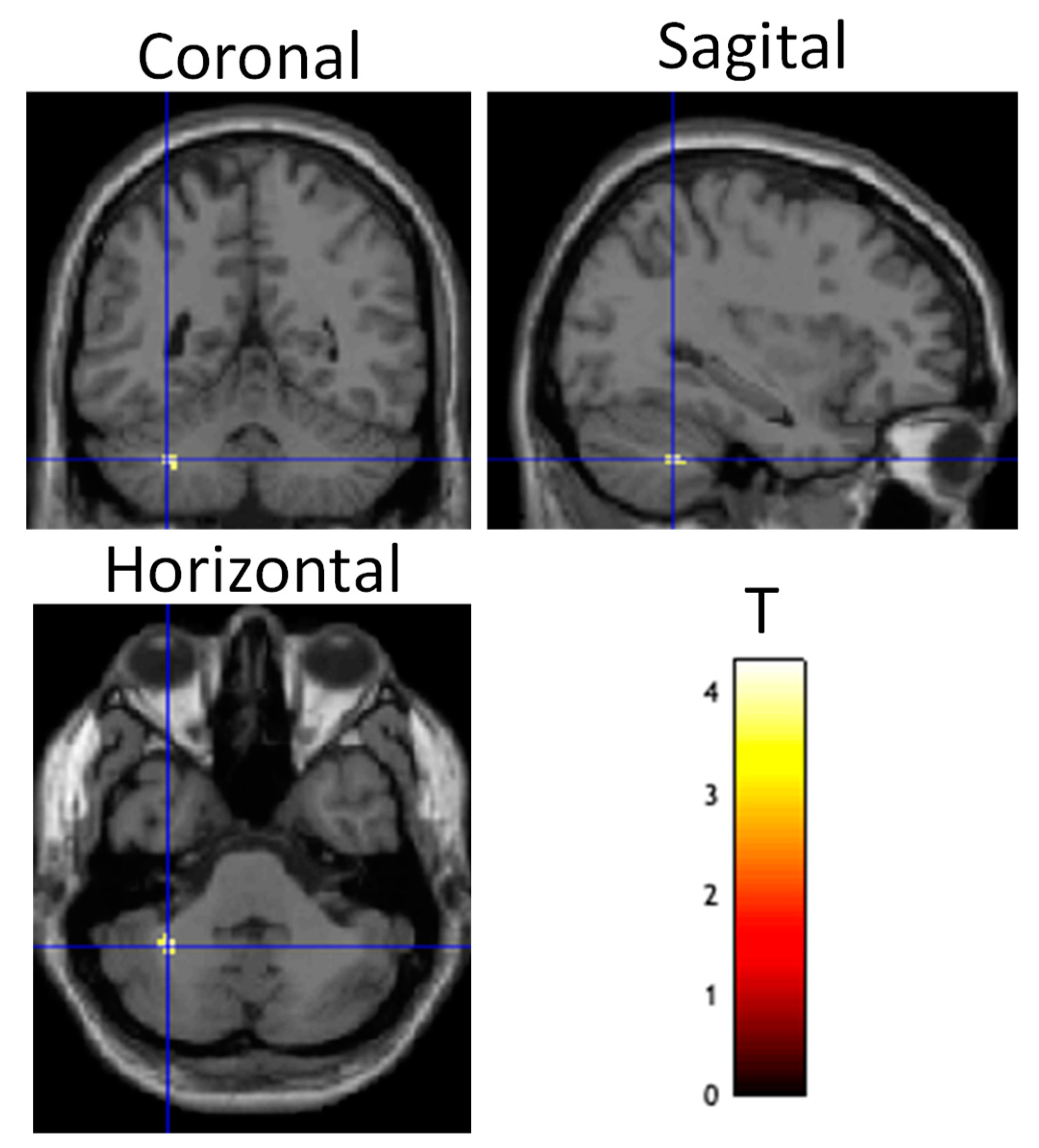
- Post((Fly_Listen_Hard—Fly_Listen_Easy)—(Fly_NoListen_Hard—Fly_NoLiten_Easy))—Pre((Fly_Listen_Hard—Fly_Listen_Easy)—(Fly_NoListen_Hard—Fly_NoListen_Easy)). This contrast is essentially looking at differences in brain activity post relative to pretraining for the most difficult flying condition requiring dual-task attention controlling for task and stimulus variables. It is predicted that this contrast will be most sensitive to learning effects induced by tDCS stimulation. In particular, brain regions involved with perceptual-motor control related to piloting during landing are expected to show greater differential activity between the tDCS stim and sham groups. These brain regions include the cerebellum, basal ganglia, and premotor cortex. Region-of-interest (ROI) analyses were conducted using masks constructed from the WFU PickAtlas Tool in SPM12 for the left and right cerebellum, as well as the left and right caudate of the basal ganglia. The mask used for the premotor cortex included both ventral and rostral maps given in Neubert et al. [67];
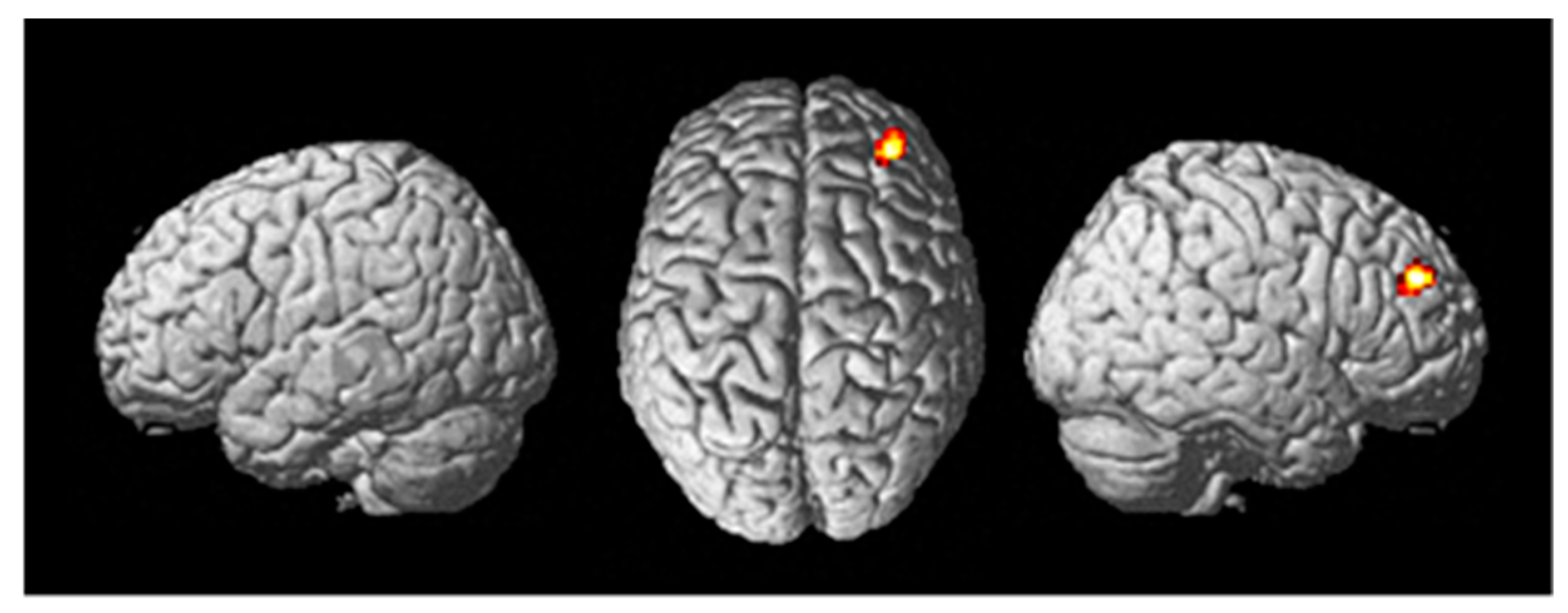
- Training((Fly_Listen_Hard—Fly_Listen_Easy)—(Fly_NoListen_Hard—Fly_NoListen_Easy)). This contrast looks at brain activity during the training session for the most difficult flying condition requiring dual-task attention controlling for task and stimulus variables. It is predicted that differences in brain regions activated to a greater extent during tDCS stimulation will be identified. In particular, the DLPFC, the site of tDCS stimulation, is predicted to show significant differential activity. The ROI map used for the DLPFC was from Sallet et al. [68] including both area 9/46 dorsal and area 9/46 ventral;
- Psychophysiological Interaction Analysis (PPI) for the Training((Fly_Listen_Hard—Fly_Listen_Easy)—(Fly_NoListen_Hard—Fly_NoListen_Easy)) Contrast. Standard procedures in SPM12 were used for the PPI analysis. This included first extracting the neural activity of the seed voxel (VOI) in the DLPFC (MNI 34,46,28) by deconvolution with the hemodynamic response for the contrast listed above using the PPI function in SPM. The results of this analysis are three files: PPI.ppi (the interaction term created by the PPI analysis), PPI.Y (The time-series extracted from the VOI), and PPI.P (the convolved onset times). These three files together with the 6 realignment parameters (variables of noninterest) are then used as regressors in an SPM fixed-effect analysis separately for each participant. The resultant PPI contrast image for each participant is used in the between groups’ (tDCS stim vs. sham) random-effects analysis. ROI analyses were conducted assessing the connectivity from the site of stimulation, DLPFC, to brain regions predicted to be involved with perceptual-motor control of airplane piloting during landing (cerebellum, basal ganglia, and premotor cortex);
- Training(Aud_Listen_Hard—Aud_Listen_Easy). It is hypothesized that tDCS will act to focus brain processes involved with the primary training task (in this case the airplane landing task) and suppress brain processes involved with other tasks (e.g., the auditory response task). It is therefore predicted that when the flying task is more difficult there will be greater suppression of auditory (BA41 and 42) and speech (premotor cortex, Broca’s area) processing regions in the brain for the active stimulation over the sham group. The ROI for the auditory cortex was made from the WFU PickAtlas Tool in SPM12 including Brodmann Area 41 and 42 with a dilation of 1. The ROI map for the dorsal premotor cortex PMd was from Sallet et.al. [68]. The ROI map for Broca’s area was defined as BA44 ventral by Neubert et al. [67].
2.7. Behavioral Performance Measures and Analysis
3. Results
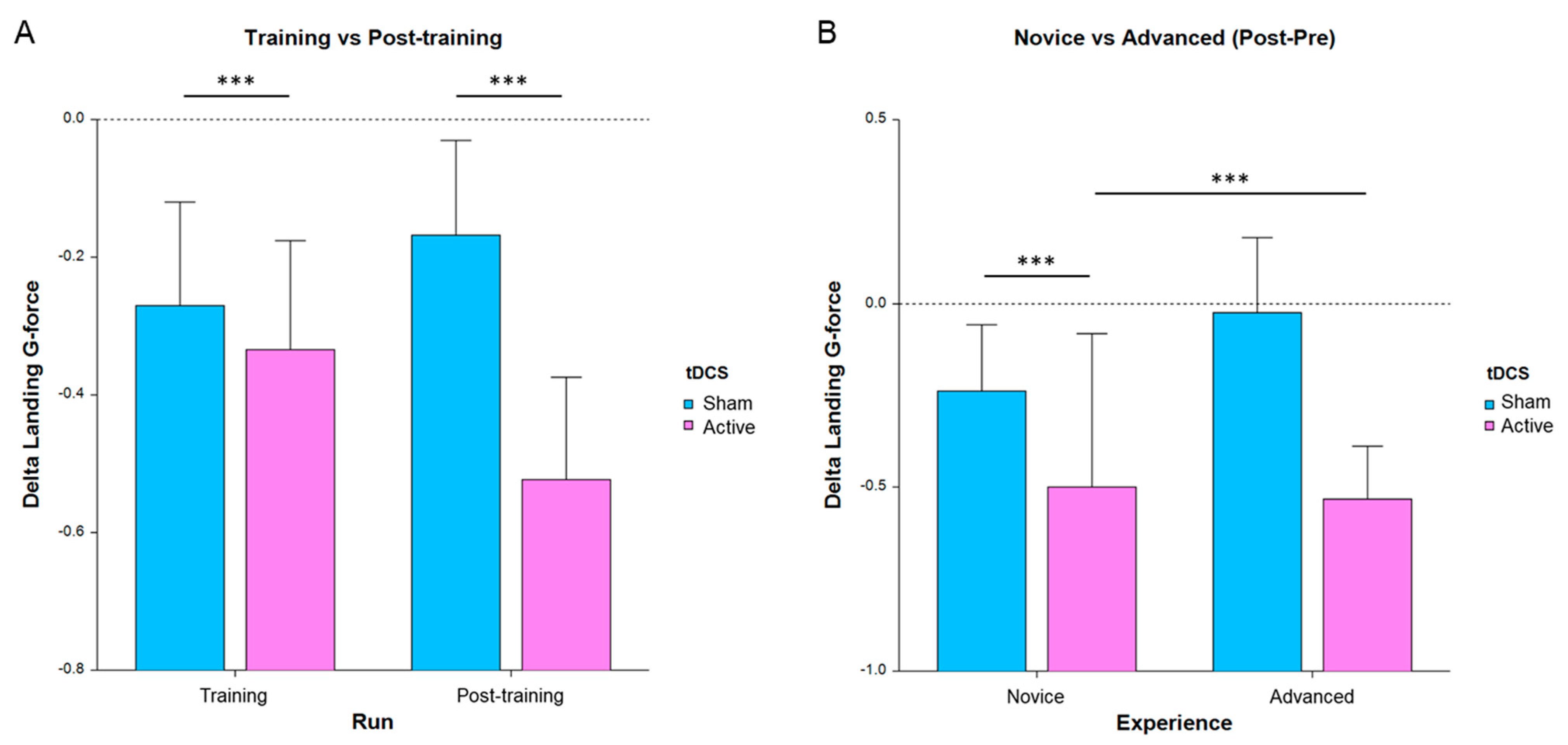
3.1. Behavioral Performance Measures
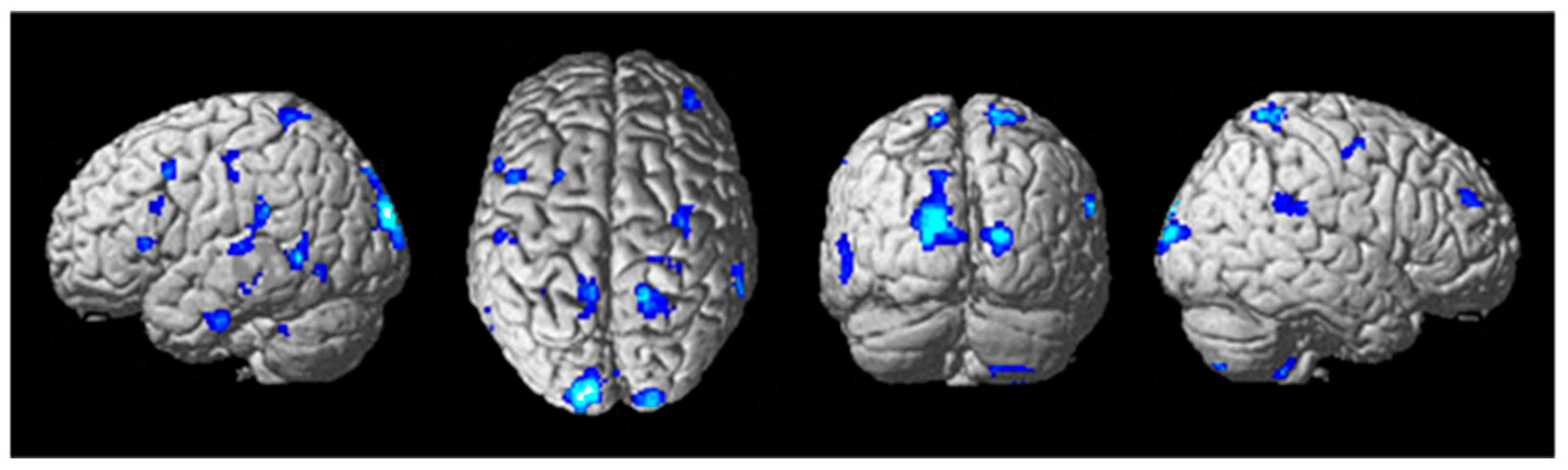
3.2. Brain Function Measures
4. Discussion
4.1. Effects on Behavioral Performance and Expertise
4.2. Brain Activity and Connectivity Modulation
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Federal Aviation Administration, United States Department of Transportation. Hourly Requirements. Available online: https://www.faa.gov/faq (accessed on 9 June 2023).
- American College of Surgeons. How Many Years of Postgraduate Training Do Surgical Residents Undergo? Available online: https://www.facs.org/for-medical-professionals/education (accessed on 9 June 2023).
- Murray, G.; Green, J. After COVID-19, Aviation Faces A Pilot Shortage. Available online: https://www.oliverwyman.com/our-expertise/insights/2021/mar/after-covid-19-aviation-faces-a-pilot-shortage.html (accessed on 9 June 2023).
- Robeznieks, A. Doctor Shortages Are Here—And They’ll Get Worse if We Don’t Act Fast. Available online: https://www.ama-assn.org/practice-management/sustainability/doctor-shortages-are-here-and-they-ll-get-worse-if-we-don-t-act (accessed on 9 June 2023).
- International Civil Aviation Organization. Multi-Pronged Approach to Enhancing Pilot Training and Competency During a Period of Anticipated Growth and Complexity; A40-WP/122; 2/8/2019. 2019. Available online: https://www.icao.int/Meetings/a40/Documents/WP/wp_122_en.pdf (accessed on 9 June 2023).
- Parasuraman, R.; Christensen, J.; Grafton, S. Neuroergonomics: The brain in action and at work. Neuroimage 2012, 59, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Dehais, F.; Karwowski, W.; Ayaz, H. Brain at Work and in Everyday Life as the Next Frontier: Grand Field Challenges for Neuroergonomics. Front. Neuroergonomics 2020, 1, 583733. [Google Scholar] [CrossRef]
- Ayaz, H.; Baker, W.B.; Blaney, G.; Boas, D.A.; Bortfeld, H.; Brady, K.; Brake, J.; Brigadoi, S.; Buckley, E.M.; Carp, S.A.; et al. Optical imaging and spectroscopy for the study of the human brain: Status report. Neurophotonics 2022, 9, S24001. [Google Scholar] [CrossRef] [PubMed]
- Tinga, A.M.; de Back, T.T.; Louwerse, M.M. Non-invasive neurophysiological measures of learning: A meta-analysis. Neurosci. Biobehav. Rev. 2019, 99, 59–89. [Google Scholar] [CrossRef] [PubMed]
- Clark, V.P.; Parasuraman, R. Neuroenhancement: Enhancing brain and mind in health and in disease. Neuroimage 2014, 85, 889–894. [Google Scholar] [CrossRef] [Green Version]
- de Souza, A.C.S.; Yehia, H.C.; Sato, M.-a.; Callan, D. Brain activity underlying auditory perceptual learning during short period training: Simultaneous fMRI and EEG recording. BMC Neurosci. 2013, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Callan, D.E.; Schweighofer, N. Positive and negative modulation of word learning by reward anticipation. Hum. Brain Mapp. 2008, 29, 237–249. [Google Scholar] [CrossRef]
- Callan, D.E.; Schweighofer, N. Neural correlates of the spacing effect in explicit verbal semantic encoding support the deficient-processing theory. Hum. Brain Mapp. 2010, 31, 645–659. [Google Scholar] [CrossRef]
- Callan, A.M.; Callan, D.E.; Masaki, S. When meaningless symbols become letters: Neural activity change in learning new phonograms. Neuroimage 2005, 28, 553–562. [Google Scholar] [CrossRef]
- Callan, D.E.; Tajima, K.; Callan, A.M.; Kubo, R.; Masaki, S.; Akahane-Yamada, R. Learning-induced neural plasticity associated with improved identification performance after training of a difficult second-language phonetic contrast. Neuroimage 2003, 19, 113–124. [Google Scholar] [CrossRef]
- Callan, D.; Perrey, S. The Use of tDCS and rTMS Methods in Neuroergonomics. In Neuroergonomics; Elsevier: Amsterdam, The Netherlands, 2019; pp. 31–33. [Google Scholar]
- Falcone, B.; Wada, A.; Parasuraman, R.; Callan, D.E. Individual differences in learning correlate with modulation of brain activity induced by transcranial direct current stimulation. PLoS ONE 2018, 13, e0197192. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Woodman, G.F. Converging Evidence That Neural Plasticity Underlies Transcranial Direct-Current Stimulation. J. Cogn. Neurosci. 2021, 33, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Chase, H.W.; Boudewyn, M.A.; Carter, C.S.; Phillips, M.L. Transcranial direct current stimulation: A roadmap for research, from mechanism of action to clinical implementation. Mol. Psychiatry 2020, 25, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, L.; Koslowsky, M.; Lavidor, M. tDCS polarity effects in motor and cognitive domains: A meta-analytical review. Exp. Brain Res. 2012, 216, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, R. Neuroergonomics: Brain, cognition, and performance at work. Curr. Dir. Psychol. Sci. 2011, 20, 181–186. [Google Scholar] [CrossRef]
- Gateau, T.; Ayaz, H.; Dehais, F. In silico vs. Over the Clouds: On-the-Fly Mental State Estimation of Aircraft Pilots, Using a Functional Near Infrared Spectroscopy Based Passive-BCI. Front. Hum. Neurosci. 2018, 12, 187. [Google Scholar] [CrossRef] [Green Version]
- Parasuraman, R.; McKinley, R.A. Using noninvasive brain stimulation to accelerate learning and enhance human performance. Hum. Factors 2014, 56, 816–824. [Google Scholar] [CrossRef] [Green Version]
- Cerreta, A.G.B.; Mruczek, R.E.B.; Berryhill, M.E. Predicting Working Memory Training Benefits From Transcranial Direct Current Stimulation Using Resting-State fMRI. Front. Psychol. 2020, 11, 570030. [Google Scholar] [CrossRef]
- Callan, D.E.; Falcone, B.; Wada, A.; Parasuraman, R. Simultaneous tDCS-fMRI Identifies Resting State Networks Correlated with Visual Search Enhancement. Front. Hum. Neurosci. 2016, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- McKendrick, R.; Falcone, B.; Scheldrup, M.; Ayaz, H. Effects of Transcranial Direct Current Stimulation on Baseline and Slope of Prefrontal Cortex Hemodynamics During a Spatial Working Memory Task. Front. Hum. Neurosci. 2020, 14, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Gong, Y.; Huang, P.; Zhang, Y.; Guo, Z.; Zhu, X.; You, X. Effect of Repeated Anodal HD-tDCS on Executive Functions: Evidence From a Pilot and Single-Blinded fNIRS Study. Front. Hum. Neurosci. 2021, 14, 583730. [Google Scholar] [CrossRef]
- Choe, J.; Coffman, B.A.; Bergstedt, D.T.; Ziegler, M.D.; Phillips, M.E. Transcranial Direct Current Stimulation Modulates Neuronal Activity and Learning in Pilot Training. Front. Hum. Neurosci 2016, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Y.; Wang, N.; Du, J.; Kong, L.; Liu, S.; Xu, M.; An, X.; Ming, D. The Effects of Transcranial Direct Current Stimulation (tDCS) on Working Memory Training in Healthy Young Adults. Front. Hum. Neurosci. 2019, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Dedoncker, J.; Brunoni, A.R.; Baeken, C.; Vanderhasselt, M.-A. A systematic review and meta-analysis of the effects of transcranial direct current stimulation (tDCS) over the dorsolateral prefrontal cortex in healthy and neuropsychiatric samples: Influence of stimulation parameters. Brain Stimul. 2016, 9, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Dong, S.; Jeong, J.; Yoon, B. Effects of Transcranial Direct Current Stimulation Over the Dorsolateral Prefrontal Cortex (PFC) on Cognitive-Motor Dual Control Skills. Percept. Mot. Ski. 2020, 127, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Ljubisavljevic, M.R.; Oommen, J.; Filipovic, S.; Bjekic, J.; Szolics, M.; Nagelkerke, N. Effects of tDCS of Dorsolateral Prefrontal Cortex on Dual-Task Performance Involving Manual Dexterity and Cognitive Task in Healthy Older Adults. Front. Aging Neurosci. 2019, 11, 144. [Google Scholar] [CrossRef]
- Jeon, S.Y.; Han, S.J. Improvement of the working memory and naming by transcranial direct current stimulation. Ann. Rehabil. Med. 2012, 36, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.-J.; Tseng, P.; Chang, C.-F.; Pai, M.-C.; Hsu, K.-S.; Lin, C.-C.; Juan, C.-H. Modulating the interference effect on spatial working memory by applying transcranial direct current stimulation over the right dorsolateral prefrontal cortex. Brain Cogn. 2014, 91, 87–94. [Google Scholar] [CrossRef]
- Figeys, M.; Loucks, T.M.; Leung, A.W.S.; Kim, E.S. Transcranial direct current stimulation over the right dorsolateral prefrontal cortex increases oxyhemoglobin concentration and cognitive performance dependent on cognitive load. Behav. Brain Res. 2023, 443, 114343. [Google Scholar] [CrossRef]
- Rocha, K.; Marinho, V.; Magalhães, F.; Carvalho, V.; Fernandes, T.; Ayres, M.; Crespo, E.; Velasques, B.; Ribeiro, P.; Cagy, M.; et al. Unskilled shooters improve both accuracy and grouping shot having as reference skilled shooters cortical area: An EEG and tDCS study. Physiol. Behav. 2020, 224, 113036. [Google Scholar] [CrossRef] [PubMed]
- Iyer, M.B.; Mattu, U.; Grafman, J.; Lomarev, M.; Sato, S.; Wassermann, E.M. Safety and cognitive effect of frontal DC brain polarization in healthy individuals. Neurology 2005, 64, 872–875. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Ugawa, Y. Adverse events of tDCS and tACS: A review. Clin. Neurophysiol. Pract. 2017, 2, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Bikson, M.; Grossman, P.; Thomas, C.; Zannou, A.L.; Jiang, J.; Adnan, T.; Mourdoukoutas, A.P.; Kronberg, G.; Truong, D.; Boggio, P. Safety of transcranial direct current stimulation: Evidence based update 2016. Brain Stimul. 2016, 9, 641–661. [Google Scholar] [CrossRef] [Green Version]
- Brunoni, A.R.; Nitsche, M.A.; Bolognini, N.; Bikson, M.; Wagner, T.; Merabet, L.; Edwards, D.J.; Valero-Cabre, A.; Rotenberg, A.; Pascual-Leone, A. Clinical research with transcranial direct current stimulation (tDCS): Challenges and future directions. Brain Stimul. 2012, 5, 175–195. [Google Scholar] [CrossRef] [Green Version]
- Antal, A.; Alekseichuk, I.; Bikson, M.; Brockmöller, J.; Brunoni, A.R.; Chen, R.; Cohen, L.; Dowthwaite, G.; Ellrich, J.; Flöel, A. Low intensity transcranial electric stimulation: Safety, ethical, legal regulatory and application guidelines. Clin. Neurophysiol. 2017, 128, 1774–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitsche, M.A.; Bikson, M. Extending the parameter range for tDCS: Safety and tolerability of 4 mA stimulation. Brain Stimul. Basic Transl. Clin. Res. Neuromodulation 2017, 10, 541–542. [Google Scholar] [CrossRef]
- Feldman, L.S.; Cao, J.; Andalib, A.; Fraser, S.; Fried, G.M. A method to characterize the learning curve for performance of a fundamental laparoscopic simulator task: Defining “learning plateau” and “learning rate”. Surgery 2009, 146, 381–386. [Google Scholar] [CrossRef]
- Bjekić, J.; Vulić, K.; Živanović, M.; Vujičić, J.; Ljubisavljević, M.; Filipović, S.R. The immediate and delayed effects of single tDCS session over posterior parietal cortex on face-word associative memory. Behav. Brain Res. 2019, 366, 88–95. [Google Scholar] [CrossRef]
- Toth, A.J.; Ramsbottom, N.; Constantin, C.; Milliet, A.; Campbell, M.J. The effect of expertise, training and neurostimulation on sensory-motor skill in esports. Comput. Hum. Behav. 2021, 121, 106782. [Google Scholar] [CrossRef]
- Ayaz, H.; Shewokis, P.A.; Bunce, S.; Izzetoglu, K.; Willems, B.; Onaral, B. Optical brain monitoring for operator training and mental workload assessment. Neuroimage 2012, 59, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Chua, Z.K.; Remy, F. Influences of age, mental workload, and flight experience on cognitive performance and prefrontal activity in private pilots: A fNIRS study. Sci. Rep. 2019, 9, 7688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayaz, H.; Onaral, B.; Izzetoglu, K.; Shewokis, P.A.; McKendrick, R.; Parasuraman, R. Continuous monitoring of brain dynamics with functional near infrared spectroscopy as a tool for neuroergonomic research: Empirical examples and a technological development. Front. Hum. Neurosci. 2013, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callan, D.E.; Naito, E. Neural Processes Distinguishing Elite from Expert and Novice Athletes. Cogn. Behav. Neurol. 2014, 27, 183–188. [Google Scholar] [CrossRef]
- Kim, W.; Chang, Y.; Kim, J.; Seo, J.; Ryu, K.; Lee, E.; Woo, M.; Janelle, C.M. An fMRI study of differences in brain activity among elite, expert, and novice archers at the moment of optimal aiming. Cogn. Behav. Neurol. 2014, 27, 173–182. [Google Scholar] [CrossRef]
- Esmaeilpour, Z.; Shereen, A.D.; Ghobadi-Azbari, P.; Datta, A.; Woods, A.J.; Ironside, M.; O’Shea, J.; Kirk, U.; Bikson, M.; Ekhtiari, H. Methodology for tDCS integration with fMRI. Hum. Brain Mapp. 2019, 41, 1950–1967. [Google Scholar] [CrossRef] [Green Version]
- van Weelden, E.; Alimardani, M.; Wiltshire, T.J.; Louwerse, M.M. Aviation and neurophysiology: A systematic review. Appl. Ergon. 2022, 105, 103838. [Google Scholar] [CrossRef]
- Callan, D.E.; Dehais, F. Neuroergonomics for Aviation. In Neuroergonomics; Elsevier: Amsterdam, The Netherlands, 2019; pp. 55–58. [Google Scholar]
- Ayaz, H.; Pakir, M.; Izzetoglu, K.; Curtin, A.; Shewokis, P.A.; Bunce, S.C.; Onaral, B. Monitoring Expertise Development during Simulated UAV Piloting Tasks Using Optical Brain Imaging. In Proceedings of the 2012 IEEE Aerospace Conference, Big Sky, MT, USA, 3–10 March 2012; IEEE: Piscataway, NJ, USA, 2012; pp. 1–11. [Google Scholar]
- Callan, D.E.; Gamez, M.; Cassel, D.B.; Terzibas, C.; Callan, A.; Kawato, M.; Sato, M.A. Dynamic visuomotor transformation involved with remote flying of a plane utilizes the ’Mirror Neuron’ system. PLoS ONE 2012, 7, e33873. [Google Scholar] [CrossRef]
- Mark, J.A.; Kraft, A.E.; Ziegler, M.D.; Ayaz, H. Neuroadaptive Training via fNIRS in Flight Simulators. Front. Neuroergonomics 2022, 3, 820523. [Google Scholar] [CrossRef]
- Gougelet, R.J.; Terzibas, C.; Callan, D.E. Cerebellum, Basal Ganglia, and Cortex Mediate Performance of an Aerial Pursuit Task. Front. Hum. Neurosci. 2020, 14, 29. [Google Scholar] [CrossRef]
- Barbey, A.K.; Koenigs, M.; Grafman, J. Dorsolateral prefrontal contributions to human working memory. Cortex 2013, 49, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.L.; Cummings, J. The Human Frontal Lobes: Functions and Disorders, 2nd ed.; Guilford: New York, NY, USA, 2007. [Google Scholar]
- Callan, D.E.; Fukada, T.; Dehais, F.; Ishii, S. The role of brain-localized gamma and alpha oscillations in inattentional deafness: Implications for understanding human attention. Front. Hum. Neurosci. 2023, 17, 1168108. [Google Scholar] [CrossRef]
- Callan, D.E.; Terzibas, C.; Cassel, D.B.; Callan, A.; Kawato, M.; Sato, M.-a. Differential activation of brain regions involved with error-feedback and imitation based motor simulation when observing self and an expert’s actions in pilots and non-pilots on a complex glider landing task. NeuroImage 2013, 72, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Curtin, A.; Ayaz, H.; Tang, Y.; Sun, J.; Wang, J.; Tong, S. Enhancing neural efficiency of cognitive processing speed via training and neurostimulation: An fNIRS and TMS study. Neuroimage 2019, 198, 73–82. [Google Scholar] [CrossRef] [PubMed]
- McKendrick, R.; Parasuraman, R.; Ayaz, H. Wearable functional Near Infrared Spectroscopy (fNIRS) and transcranial Direct Current Stimulation (tDCS): Expanding Vistas for Neurocognitive Augmentation. Front. Syst. Neurosci. 2015, 9, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, J.; Ayaz, H.; Callan, D. Developing a tDCS-Enhanced Dual-Task Flight Simulator for Evaluating Learning. In Advances in Neuroergonomics and Cognitive Engineering: Proceedings of the AHFE 2020 Virtual Conferences on Neuroergonomics and Cognitive Engineering, and Industrial Cognitive Ergonomics and Engineering Psychology, July 16–20, 2020, USA, 2021; Springer: Berlin/Heidelberg, Germany, 2020; pp. 149–155. [Google Scholar]
- Durantin, G.; Dehais, F.; Gonthier, N.; Terzibas, C.; Callan, D.E. Neural signature of inattentional deafness. Hum. Brain Mapp. 2017, 38, 5440–5455. [Google Scholar] [CrossRef] [Green Version]
- Neubert, F.-X.; Mars, R.B.; Thomas, A.G.; Sallet, J.; Rushworth, M.F. Comparison of human ventral frontal cortex areas for cognitive control and language with areas in monkey frontal cortex. Neuron 2014, 81, 700–713. [Google Scholar] [CrossRef] [Green Version]
- Sallet, J.; Mars, R.B.; Noonan, M.P.; Neubert, F.-X.; Jbabdi, S.; O’Reilly, J.X.; Filippini, N.; Thomas, A.G.; Rushworth, M.F. The organization of dorsal frontal cortex in humans and macaques. J. Neurosci. 2013, 33, 12255–12274. [Google Scholar] [CrossRef] [Green Version]
- Doya, K. What are the computations of the cerebellum, the basal ganglia and the cerebral cortex? Neural Netw. 1999, 12, 961–974. [Google Scholar] [CrossRef]
- Imamizu, H.; Kawato, M. Cerebellar internal models: Implications for the dexterous use of tools. Cerebellum 2012, 11, 325–335. [Google Scholar] [CrossRef]
- Higuchi, S.; Imamizu, H.; Kawato, M. Cerebellar activity evoked by common tool-use execution and imagery tasks: An fMRI study. Cortex 2007, 43, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Imamizu, H.; Kuroda, T.; Miyauchi, S.; Yoshioka, T.; Kawato, M. Modular organization of internal models of tools in the human cerebellum. Proc. Natl. Acad. Sci. USA 2003, 100, 5461–5466. [Google Scholar] [CrossRef] [PubMed]
- Imamizu, H.; Miyauchi, S.; Tamada, T.; Sasaki, Y.; Takino, R.; PuÈtz, B.; Yoshioka, T.; Kawato, M. Human cerebellar activity reflecting an acquired internal model of a new tool. Nature 2000, 403, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Kawato, M. Internal models for motor control and trajectory planning. Curr. Opin. Neurobiol. 1999, 9, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Caligiore, D.; Pezzulo, G.; Baldassarre, G.; Bostan, A.C.; Strick, P.L.; Doya, K.; Helmich, R.C.; Dirkx, M.; Houk, J.; Jörntell, H.; et al. Consensus Paper: Towards a Systems-Level View of Cerebellar Function: The Interplay Between Cerebellum, Basal Ganglia, and Cortex. Cerebellum 2017, 16, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Dehais, F.; Callan, D. A neuroergonomics Approach to Human Performance in Aviation. In Improving Aviation Performance through Applying Engineering Psychology; CRC Press: Boca Raton, FL, USA, 2019; pp. 123–141. [Google Scholar]
- Callan, D.E.; Terzibas, C.; Cassel, D.B.; Sato, M.-a.; Parasuraman, R. The Brain Is Faster than the Hand in Split-Second Intentions to Respond to an Impending Hazard: A Simulation of Neuroadaptive Automation to Speed Recovery to Perturbation in Flight Attitude. Front. Hum. Neurosci. 2016, 10, 187. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run 1 | Run 2 | Run 3 | |||
|---|---|---|---|---|---|
| Rest | Pre-Training | Rest | Training | Rest | Post-Training |
| 5 min | 20 min | 5 min | 20 min | 5 min | 20 min |
| No feedback | Performance feedback | No feedback | |||
| No tDCS | Active Stim/Sham tDCS | No tDCS | |||
| Wind Condition | Auditory Task | Runway Condition | |
|---|---|---|---|
| Type 1 | Absent | Ignore | Wide |
| Type 2 | Present | Perform | Narrow |
| Type 3 | Absent | Perform | Narrow |
| Type 4 | Present | Ignore | Wide |
| Type 5 | Absent | Perform | Wide |
| Type 6 | Present | Ignore | Narrow |
| Type 7 | Absent | Ignore | Narrow |
| Type 8 | Present | Perform | Wide |
| Term | Factor | F-Ratio | p-Value | Partial η2 | |
|---|---|---|---|---|---|
| Landing G-force Main Effects (Post-Pre) | |||||
| tDCS | 16.78 | 1/23.7 | 0.0004 *** | 0.415 | |
| Experience | 30 | 1/24.5 | 0.00001 *** | 0.550 | |
| Experience*tDCS | 14.6 | 1/21 | 0.0001 *** | 0.410 | |
| Runway | 3.02 | 1/537 | 0.08 | 0.006 | |
| Wind | 62.03 | 1/548 | 0.00000 *** | 0.102 | |
| Auditory | 0.53 | 1/537 | 0.47 | 0.001 | |
| Pretraining Performance | 270.6 | 1/489 | 0.00000 *** | 0.356 | |
| Landing G-force Post Hoc Comparisons (Post-Pre) | |||||
| Active stim, experience novice vs. advanced | 30.50 | 1/25.8 | 0.00002 *** | 0.542 | |
| Experience novice, active stim vs. sham | 20.30 | 1/25.2 | 0.0003 *** | 0.446 | |
| Landing G-force Main Effects (Training-Pre) | |||||
| tDCS | 31.9 | 1/23.3 | 0.00001 *** | 0.578 | |
| Experience | 27.4 | 1/24.1 | 0.00000 *** | 0.532 | |
| Experience*tDCS | 19.1 | 1/20.3 | 0.0003 *** | 0.485 | |
| Runway | 0.67 | 1/540 | 0.41 | 0.001 | |
| Wind | 46.1 | 1/552 | 0.00000 *** | 0.077 | |
| Auditory | 3.47 | 1/540 | 0.06 | 0.006 | |
| Pretraining Performance | 238 | 1/480 | 0.00000 *** | 0.331 | |
| Landing G-force Post Hoc Comparisons (Training-Pre) | |||||
| Active stim, experience novice vs. advanced | 55.9 | 1/24.3 | 0.00000 *** | 0.697 | |
| Experience novice, active stim vs. sham | 43.0 | 1/23.7 | 0.0000002 *** | 0.645 | |
| Landing G-force Main Effects (Training and Post-Pre) | |||||
| Run | 0.06 | 1/1102 | 0.8 | 0.000 | |
| tDCS | 34 | 1/22.3 | 0.00007 *** | 0.604 | |
| Experience | 54.7 | 1/22.9 | 0.00000 *** | 0.705 | |
| Run*tDCS | 1.67 | 1/1102 | 0.2 | 0.002 | |
| Experience*tDCS | 23.2 | 1/20.3 | 0.0001 *** | 0.533 | |
| Runway | 0.25 | 1/1103 | 0.61 | 0.000 | |
| Wind | 107.7 | 1/1118 | 0.00000 *** | 0.088 | |
| Auditory | 0.9 | 1/1103 | 0.34 | 0.001 | |
| Pretraining Performance | 501.7 | 1/948 | 0.00000 *** | 0.346 | |
| Landing G-force Post Hoc Comparisons (Training and Post-Pre) | |||||
| Run 2, tDCS stim vs. sham | 27.6 | 1/39.6 | 0.00001 *** | 0.411 | |
| Run 3, tDCS stim vs. sham | 16.6 | 1/39.6 | 0.0004 *** | 0.295 | |
| Active stim, experience novice vs. advanced | 60.4 | 1/24.3 | 0.00000 *** | 0.713 | |
| Experience novice, active stim vs. sham | 43.0 | 1/23.8 | 0.000002 *** | 0.644 | |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p |
|---|---|---|---|---|
| Cerebellum | 447 | −32, −50, −26 | 5.32 | 0.0000028 |
| Cerebellum | 77 | 8, −52, −52 | 4.52 | 0.000085 |
| Midbrain/Brain Stem | 69 | −8, −18, −20 | 4.42 | 0.00011 |
| Midbrain/Brain Stem 1 Hippocampus Thalamus | 272 | 12, −24, −10 24, −28, −10 16, −22, 8 | 5.94 4.48 4.39 | 0.0000027 0.000093 0.00012 |
| Hippocampus | 79 | −36, −18, −12 | 5.05 | 0.000024 |
| Thalamus | 24 | −8, −16, 0 | 4.27 | 0.00036 |
| Thalamus | 34 | 8, −6, 16 | 4.15 | 0.00021 |
| Caudate | 27 | −14, 14, 12 | 4.21 | 0.00018 |
| Caudate | 27 | 6, 20, −2 | 4.08 | 0.00025 |
| Frontal Operculum | 20 | 28, 32, 12 | 4.05 | 0.00046 |
| Orbital IFG | 29 | −42, 32, −8 | 4.46 | 0.000098 |
| Opercular IFG | 24 | 40, 8, 24 | 3.92 | 0.00092 |
| Medial Orbital Gyrus | 50 | 14, 6, −20 | 5.36 | 0.000011 |
| Premotor Cortex/Precentral Gyrus | 21 | −44, 2, 22 | 4.47 | 0.000095 |
| Postcentral Gyrus | 21 | 42, −10, 24 | 3.94 | 0.00035 |
| Parietal Operculum | 69 | 36, −32, 26 | 4.99 | 0.000027 |
| Precuneus/Posterior Cingulate Gyrus | 36 | 18, −40, 42 | 4.54 | 0.00008 |
| Posterior Cingulate Gyrus | 20 | −12, −40, 20 | 4.54 | 0.00008 |
| Superior Parietal Lobule Angular Gyrus | 145 | 32, −54, 44 38, −5, 34 | 4.5 3.98 | 0.00009 0.00031 |
| Middle Occipital Gyrus | 20 | 42, −66, 20 | 4.23 | 0.00017 |
| Lingual Gyrus | 21 | 6, −74, −8 | 4.12 | 0.00022 |
| Primary Auditory/Planum Temporal | 55 | 36, −28, 6 | 4.34 | 0.00013 |
| Planum Temporal | 85 | −42, −34, 4 | 5.21 | 0.000016 |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p | pFWE_corr |
|---|---|---|---|---|---|
| Cerebellum | 265 | −32, −50, −26 | 5.32 | 0.0000028 | 0.026 * |
| Cerebellum | 77 | 8, −52, −52 | 4.52 | 0.000085 | 0.110 |
| Caudate | 27 | −14, 14, 12 | 4.21 | 0.00018 | 0.025 * |
| Caudate | 23 | 6, 20, −2 | 4.08 | 0.00025 | 0.032 * |
| PMv | 2 | −44, 4, 20 | 3.75 | 0.00056 | 0.063 |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p |
|---|---|---|---|---|
| DLPFC | 131 | 34, 46, 28 | 5.71 | 0.0000049 |
| Middle Cingulate Gyrus | 67 | 8, 4, 40 | 4.56 | 0.000077 |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p | pFWE_corr |
|---|---|---|---|---|---|
| DLPFC | 94 | 34, 46, 28 | 5.71 | 0.0000049 | 0.004 * |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p |
|---|---|---|---|---|
| Cerebellum | 18 | −32, −50, −40 | 4.03 | 0.00028 |
| Brain Stem | 42 | −8, −28, −36 | 4.30 | 0.00014 |
| Occipital Pole | 26 | 20, −94, 2 | 3.84 | 0.00044 |
| MTG | 26 | 60, −40, −12 | 3.86 | 0.00042 |
| Brain ROI | Cluster Size | MNI Coordinate x, y, z | T | p | pFWE_corr |
|---|---|---|---|---|---|
| Cerebellum 1 | 18 | −32, −50, −40 | 4.03 | 0.00028 | 0.211 |
| Cerebellum 2 | 17 | −32, −50, −40 | 4.03 | 0.00028 | 0.042 * |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p |
|---|---|---|---|---|
| Cerebellum | 37 | 30, −74, −54 | 4.48 | 0.000093 |
| Cerebellum | 41 | −16, −56, −22 | 4.40 | 0.00011 |
| Cerebellum | 35 | 28, −42, −56 | 4.14 | 0.00022 |
| Cerebellum | 30 | −32, −50, −36 | 4.09 | 0.00024 |
| DLPFC | 42 | 40, 50, 30 | 4.43 | 0.00011 |
| Anterior Insula | 22 | 28, 18, 10 | 4.45 | 0.0001 |
| IFG | 33 | −50, 24, 8 | 4.27 | 0.00016 |
| IFG | 20 | −50, 16, 26 | 4.24 | 0.00017 |
| Middle Frontal Gyrus | 46 | −26, 10, 40 | 4.68 | 0.000057 |
| Premotor Cortex | 48 | −48, 10, 44 | 4.41 | 0.00041 |
| Precentral Gyrus | 91 | 34, −12, 50 | 5.39 | 0.00001 |
| Medial Precentral Gyrus SMA Middle Cingulate Gyrus | 145 | 10, −20, 50 10, −2, 46 8, −12, 44 | 6.16 4.85 4.18 | 0.0000017 0.000038 0.00019 |
| Postcentral Gyrus | 30 | −48, −20, 42 | 4.34 | 0.00013 |
| Postcentral Gyrus | 20 | 20, −34, 56 | 3.80 | 0.0005 |
| Postcentral Gyrus | 19 | 30, −34, 50 | 4.00 | 0.0003 |
| Medial Postcentral Gyrus Superior Parietal Lobule | 122 | −14, −42, 56 −10, −48, 70 | 4.56 4.42 | 0.000077 0.00011 |
| Superior Parietal Lobule Postcentral Gyrus | 174 | 18, −56, 72 14, −44, 72 | 4.48 4.05 | 0.00093 0.00027 |
| Superior Parietal Lobule | 15 | −34, −48, 44 | 3.93 | 0.00035 |
| SMG/AnG | 62 | 64, −46, 28 | 4.38 | 0.00012 |
| Lingual Gyrus/Calcarine Cortex | 22 | 18, −62, 2 | 4.16 | 0.0002 |
| Lingual Gyrus/Calcarine Cortex | 28 | −16, −66, −2 | 4.12 | 0.00023 |
| Superior Occipital Gyrus | 655 * | −14, −96, 22 | 6.64 ** | 0.00000056 |
| Occipital Pole | 171 | 20, −102, 10 | 5.05 | 0.000024 |
| ITG/MTG | 63 | −46, −12, −30 | 5.12 | 0.00002 |
| ITG/MTG | 72 | −46, −36, −16 | 4.81 | 0.000042 |
| MTG | 87 | −52, −52, 0 | 4.64 | 0.000063 |
| MTG | 18 | −58, −66, −6 | 3.94 | 0.00035 |
| Primary Auditory Cortex Parietal Operculum Planum Temporal | 152 | −48, −24, 6 −54, −38, 24 −52, −32, 16 | 4.66 4.42 4.09 | 0.00006 0.00011 0.00024 |
| Brain Region | Cluster Size | MNI Coordinate x, y, z | T | p | pFWE_corr |
|---|---|---|---|---|---|
| Primary Auditory Cortex | 51 | −48, −24, 6 | 4.66 | 0.00006 | 0.013 * |
| PMd | 19 | −26, 12, 46 | 4.26 | 0.00016 | 0.039 * |
| BA44v | 6 | −50, 22, 10 | 3.88 | 0.00041 | 0.030 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mark, J.A.; Ayaz, H.; Callan, D.E. Simultaneous fMRI and tDCS for Enhancing Training of Flight Tasks. Brain Sci. 2023, 13, 1024. https://doi.org/10.3390/brainsci13071024
Mark JA, Ayaz H, Callan DE. Simultaneous fMRI and tDCS for Enhancing Training of Flight Tasks. Brain Sciences. 2023; 13(7):1024. https://doi.org/10.3390/brainsci13071024
Chicago/Turabian StyleMark, Jesse A., Hasan Ayaz, and Daniel E. Callan. 2023. "Simultaneous fMRI and tDCS for Enhancing Training of Flight Tasks" Brain Sciences 13, no. 7: 1024. https://doi.org/10.3390/brainsci13071024
APA StyleMark, J. A., Ayaz, H., & Callan, D. E. (2023). Simultaneous fMRI and tDCS for Enhancing Training of Flight Tasks. Brain Sciences, 13(7), 1024. https://doi.org/10.3390/brainsci13071024