Sex Specific Alterations in α4*Nicotinic Receptor Expression in the Nucleus Accumbens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Varenicline and Yohimbine Administration
2.3. Maternal Separation Procedure
2.4. Restraint Stress
2.5. Elevated Plus Maze
2.6. Sample Collection and Processing
2.7. Corticosterone Measurements
2.8. Protein Measurements
2.9. Statistics
3. Results
3.1. Varenicline Attenuates Yohimbine-Induced Increases in Plasma Corticosterone
3.2. Maternal Separation Alters Body Weight in Female Mice Only
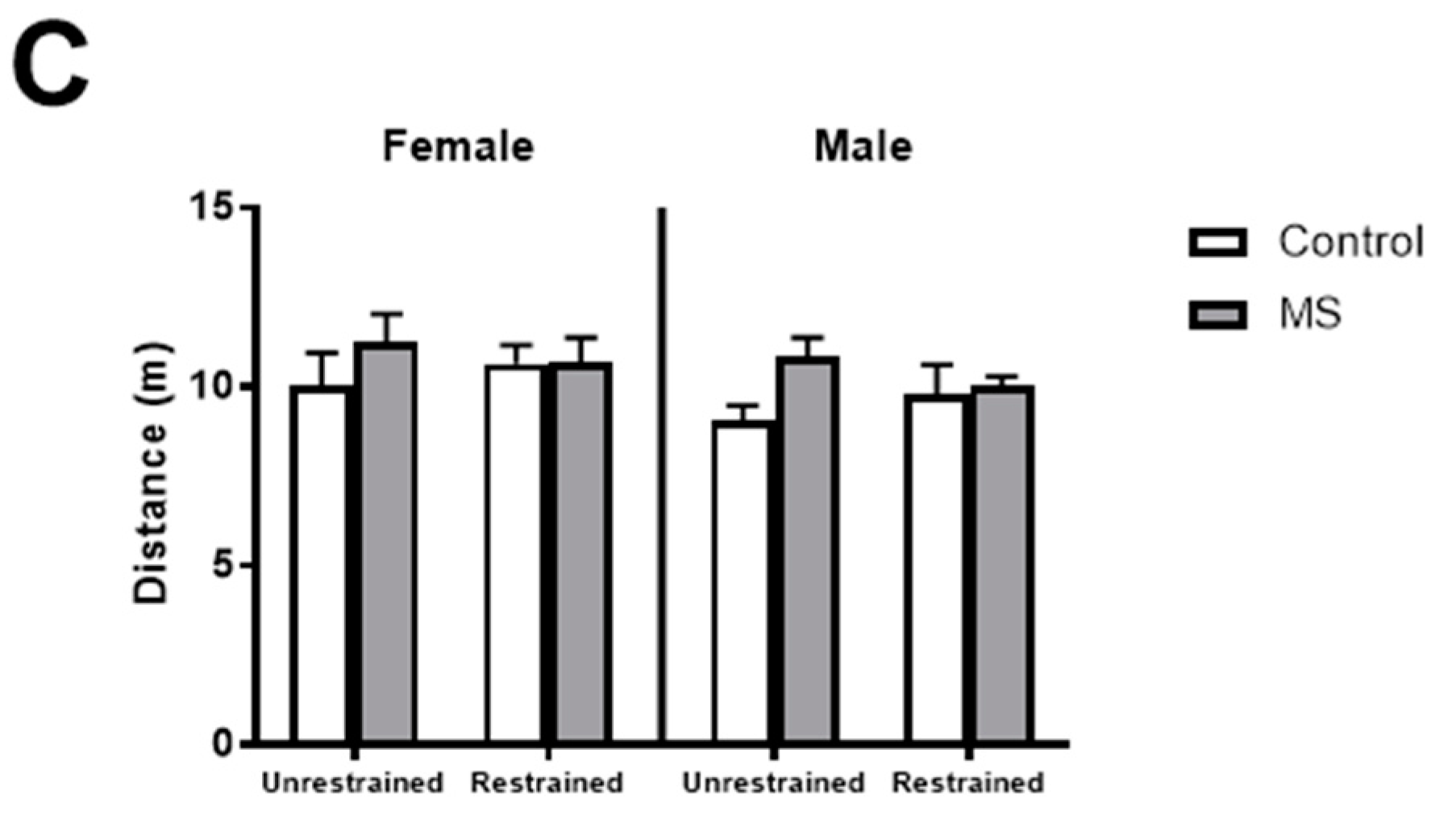
3.3. Restraint Stress Increases Anxiety-Like Behaviours in Female MS Mice
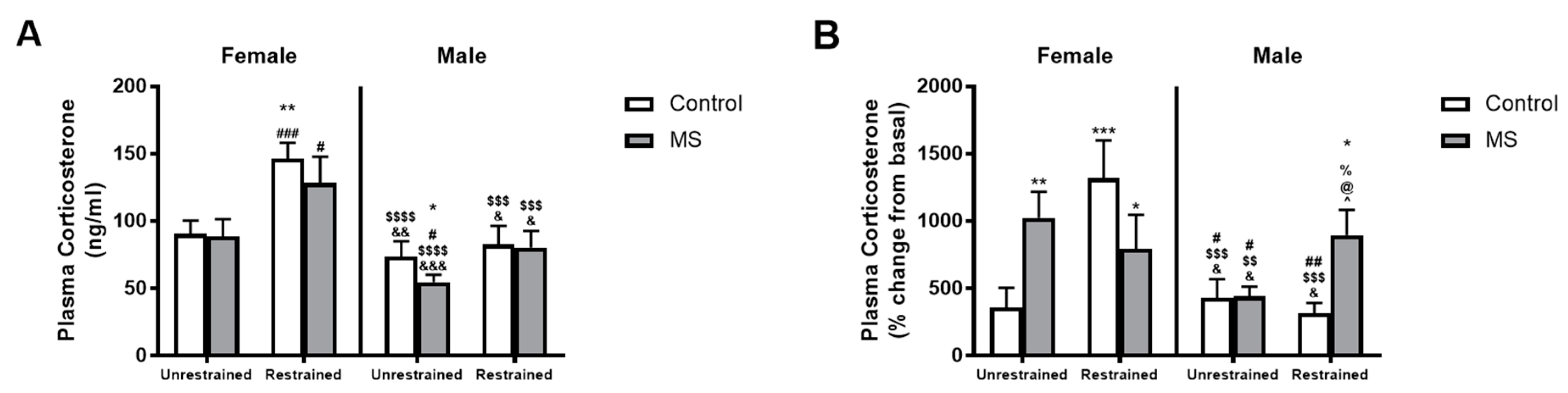
3.4. ELS Has Sex-Specific Effects on Plasma Corticosterone Levels in Response to Stress
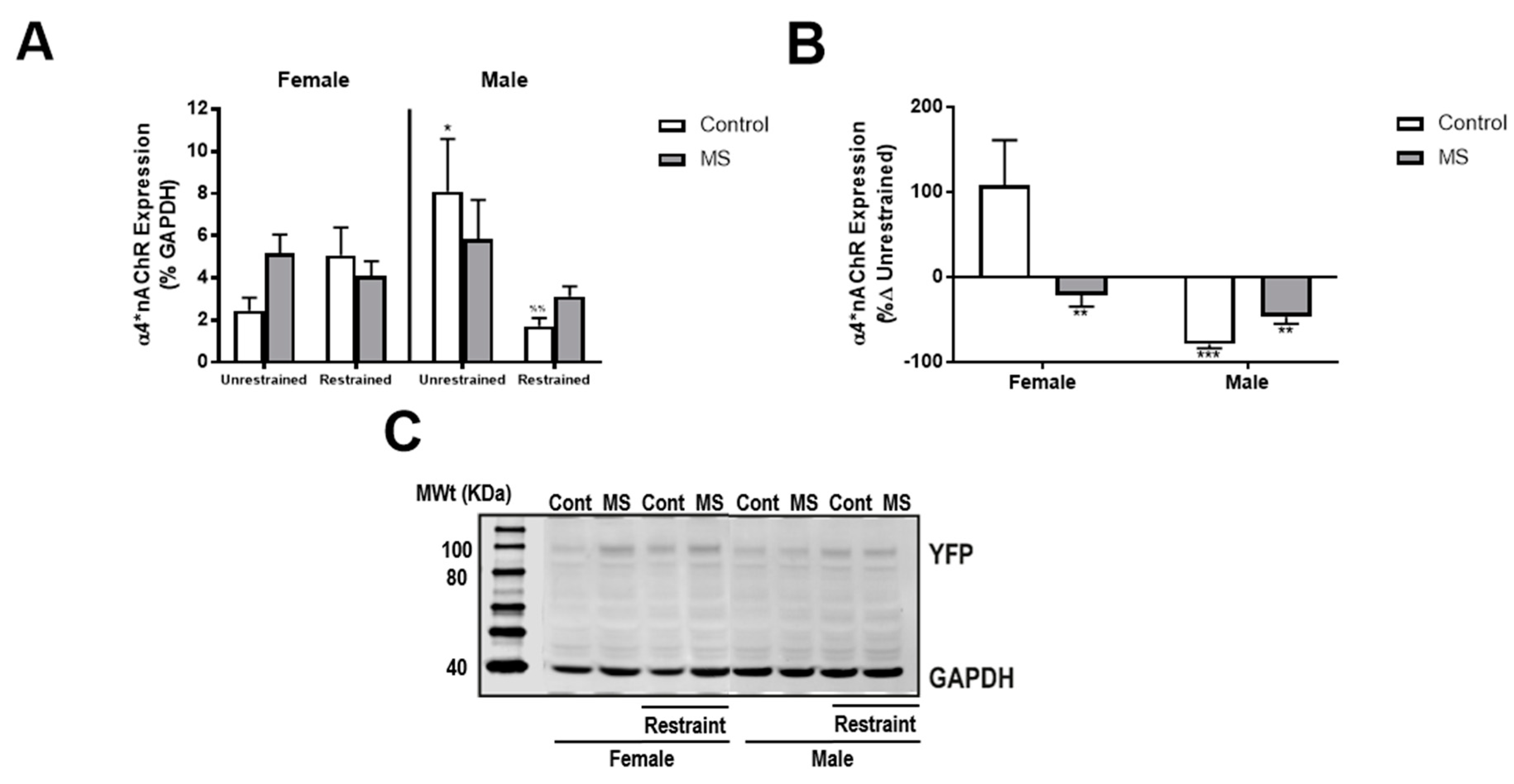
3.5. MS Attenuates Restraint-Induced Decreases in NAc α4*nAChR Expression in Male Mice
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviation
| ACh | acetylcholine |
| AChE | acetylcholinesterase |
| AUDs | alcohol use disorders |
| ELS | early life stress |
| GABA | gamma-aminobutyric acid |
| MS | maternal separation |
| nAChR | nicotinic acetylcholine receptor |
| NAc | nucleus accumbens |
| PTSD | post-traumatic stress disorder |
| PFC | prefrontal cortex |
| VTA | ventral tegmental area |
References
- McEwen, B.S. Central effects of stress hormones in health and disease: Understanding the protective and damaging effects of stress and stress mediators. Eur. J. Pharmacol. 2008, 583, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Putnam, F.W. The Impact of Trauma on Child Development. Juv. Fam. Court J. 2006, 57, 1–11. [Google Scholar] [CrossRef]
- Felitti, F.V.J.; Anda, M.S.R.F.; Nordenberg, D.; Williamson, P.D.F.; Spitz, M.P.H.A.M.; Edwards, V.; Koss, M.P.; Marks, M.P.H.J.S. Relationship of Childhood Abuse and Household Dysfunction to Many of the Leading Causes of Death in Adults: The Adverse Childhood Experiences (ACE) Study. Am. J. Prev. Med. 1998, 14, 245–258. [Google Scholar] [CrossRef]
- Andrews, M.M.; Meda, S.A.; Thomas, A.D.; Potenza, M.N.; Krystal, J.H.; Worhunsky, P.; Stevens, M.C.; O’malley, S.; Book, G.A.; Reynolds, B.; et al. Individuals family history positive for alcoholism show functional magnetic resonance imaging differences in reward sensitivity that are related to impulsivity factors. Biol. Psychiatry 2011, 69, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Cservenka, A.; Casimo, K.; Fair, D.A.; Nagel, B.J. Resting state functional connectivity of the nucleus accumbens in youth with a family history of alcoholism. Psychiatry Res. 2014, 221, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Cservenka, A.; Gillespie, A.J.; Michael, P.G.; Nagel, B.J. Family history density of alcoholism relates to left nucleus accumbens volume in adolescent girls. J. Stud. Alcohol Drugs 2015, 76, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Piao, C.; Deng, X.; Wang, X.; Yuan, Y.; Liu, Z.; Liang, J. Altered function in medial prefrontal cortex and nucleus accumbens links to stress-induced behavioral inflexibility. Behav. Brain Res. 2017, 317, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Kalon, E.; Hong, J.Y.; Tobin, C.; Schulte, T. Psychological and Neurobiological Correlates of Food Addiction. Int. Rev. Neurobiol. 2016, 129, 85–110. [Google Scholar] [PubMed]
- Tomoda, A. Preliminary Evidence for Impaired Brain Activity of Neural Reward Processing in Children and Adolescents with Reactive Attachment Disorder. Yakugak. Zasshi 2016, 136, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Vythilingam, M.; Nelson, E.E.; Scaramozza, M.; Waldeck, T.; Hazlett, G.; Southwick, S.M.; Pine, D.S.; Drevets, W.; Charney, D.S.; Ernst, M. Reward circuitry in resilience to severe trauma: An fMRI investigation of resilient special forces soldiers. Psychiatry Res. 2009, 172, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Sailer, U.; Robinson, S.; Fischmeister, F.P.; Konig, D.; Oppenauer, C.; Lueger-Schuster, B.; Moser, E.; Kryspin-Exner, I.; Bauer, H. Altered reward processing in the nucleus accumbens and mesial prefrontal cortex of patients with posttraumatic stress disorder. Neuropsychologia 2008, 46, 2836–2844. [Google Scholar] [CrossRef] [PubMed]
- Tolu, P.; Masi, F.; Leggio, B.; Scheggi, S.; Tagliamonte, A.; De Montis, M.G.; Gambarana, C. Effects of long-term acetyl-l-carnitine administration in rats: I. increased dopamine output in mesocorticolimbic areas and protection toward acute stress exposure. Neuropsychopharmacology 2002, 27, 410–420. [Google Scholar] [CrossRef]
- Sarter, M.; Nelson, C.L.; Bruno, J.P. Cortical cholinergic transmission and cortical information processing in schizophrenia. Schizophr. Bull. 2005, 31, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Jeon, W.K.; Bizon, J.L.; Han, J.S. Interaction of basal forebrain cholinergic neurons with the glucocorticoid system in stress regulation and cognitive impairment. Front. Aging Neurosci. 2015, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Adermark, L. Dopaminergic Regulation of Striatal Interneurons in Reward and Addiction: Focus on Alcohol. Neural Plast. 2015, 2015, 814567. [Google Scholar] [CrossRef] [PubMed]
- Heinz, A.; Beck, A.; Grusser, S.M.; Grace, A.A.; Wrase, J. Identifying the neural circuitry of alcohol craving and relapse vulnerability. Addict. Biol. 2009, 14, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Weiss, F.; Ciccocioppo, R.; Parsons, L.H.; Katner, S.; Liu, X.; Zorrilla, E.P.; Valdez, G.R.; Ben-Shahar, O.; Angeletti, S.; Richter, R.R. Compulsive drug-seeking behavior and relapse. Neuroadaptation, stress, and conditioning factors. Ann. N. Y. Acad. Sci. 2001, 937, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Feduccia, A.A.; Simms, J.A.; Mill, D.; Yi, H.Y.; Bartlett, S.E. Varenicline decreases ethanol intake and increases dopamine release via neuronal nicotinic acetylcholine receptors in the nucleus accumbens. Br. J. Pharmacol. 2014, 171, 3420–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanashi, K.; Miyamae, T.; Misu, Y.; Goshima, Y. Tonic function of nicotinic receptors in stress-induced release of L-DOPA from the nucleus accumbens in freely moving rats. Eur. J. Pharmacol. 2001, 424, 199–202. [Google Scholar] [CrossRef]
- Schulz, K.M.; Andrud, K.M.; Burke, M.B.; Pearson, J.N.; Kreisler, A.D.; Stevens, K.E.; Leonard, S.; Adams, C.E. The effects of prenatal stress on alpha4 beta2 and alpha7 hippocampal nicotinic acetylcholine receptor levels in adult offspring. Dev. Neurobiol. 2013, 73, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.J.; Liu, L.A.; Cheng, X.H.; Wang, C.A. Modulation of neuronal nicotinic acetylcholine receptors by glucocorticoids. Acta Pharmacol. Sin. 2002, 23, 237–242. [Google Scholar] [PubMed]
- Loomis, S.; Gilmour, G. Corticosterone urinalysis and nicotinic receptor modulation in rats. J. Neurosci. Methods 2010, 188, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Nashmi, R.; Xiao, C.; Deshpande, P.; Mckinney, S.; Grady, S.R.; Whiteaker, P.; Huang, Q.; Mcclure-Begley, T.; Lindstrom, J.M.; Labarca, C.; et al. Chronic nicotine cell specifically upregulates functional alpha 4* nicotinic receptors: Basis for both tolerance in midbrain and enhanced long-term potentiation in perforant path. J. Neurosci. 2007, 27, 8202–8218. [Google Scholar] [CrossRef] [PubMed]
- Nashmi, R.; Dickinson, M.E.; Mckinney, S.; Jareb, M.; Labarca, C.; Fraser, S.E.; Lester, H.A. Assembly of alpha4beta2 nicotinic acetylcholine receptors assessed with functional fluorescently labeled subunits: Effects of localization, trafficking, and nicotine-induced upregulation in clonal mammalian cells and in cultured midbrain neurons. J. Neurosci. 2003, 23, 11554–11567. [Google Scholar] [CrossRef] [PubMed]
- Solomon, M.B.; Loftspring, M.; De Kloet, A.D.; Ghosal, S.; Jankord, R.; Flak, J.N.; Wulsin, A.C.; Krause, E.G.; Zhang, R.; Rice, T.; et al. Neuroendocrine Function After Hypothalamic Depletion of Glucocorticoid Receptors in Male and Female Mice. Endocrinology 2015, 156, 2843–2853. [Google Scholar] [CrossRef] [PubMed]
- Van Den Buuse, M.; Martin, S.; Holgate, J.; Matthaei, K.; Hendry, I. Mice deficient in the alpha subunit of G(z) show changes in pre-pulse inhibition, anxiety and responses to 5-HT(1A) receptor stimulation, which are strongly dependent on the genetic background. Psychopharmacology 2007, 195, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Klenowski, P.M.; Shariff, M.R.; Belmer, A.; Fogarty, M.J.; Mu, E.W.; Bellingham, M.C.; Bartlett, S.E. Prolonged Consumption of Sucrose in a Binge-Like Manner, Alters the Morphology of Medium Spiny Neurons in the Nucleus Accumbens Shell. Front. Behav. Neurosci. 2016, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patkar, O.L.; Belmer, A.; Holgate, J.Y.; Tarren, J.R.; Shariff, M.R.; Morgan, M.; Fogarty, M.J.; Bellingham, M.C.; Bartlett, S.E.; Klenowski, P.M. The antihypertensive drug pindolol attenuates long-term but not short-term binge-like ethanol consumption in mice. Addict. Biol. 2017, 22, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Shariff, M.; Quik, M.; Holgate, J.; Morgan, M.; Patkar, O.L.; Tam, V.; Belmer, A.; Bartlett, S.E. Neuronal Nicotinic Acetylcholine Receptor Modulators Reduce Sugar Intake. PLoS ONE 2016, 11, e0150270. [Google Scholar] [CrossRef] [PubMed]
- Jaimes-Hoy, L.; Gutierrez-Mariscal, M.; Vargas, Y.; Perez-Maldonado, A.; Romero, F.; Sanchez-Jaramillo, E.; Charli, J.L.; Joseph-Bravo, P. Neonatal Maternal Separation Alters, in a Sex-Specific Manner, the Expression of TRH, of TRH-Degrading Ectoenzyme in the Rat Hypothalamus, and the Response of the Thyroid Axis to Starvation. Endocrinol. 2016, 157, 3253–3265. [Google Scholar] [CrossRef] [PubMed]
- Pautassi, R.M.; Nizhnikov, M.E.; Fabio, M.C.; Spear, N.E. Early maternal separation affects ethanol-induced conditioning in a nor-BNI insensitive manner, but does not alter ethanol-induced locomotor activity. Pharmacol. Biochem. Behav. 2012, 100, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Ryu, V.; Yoo, S.B.; Kang, D.W.; Lee, J.H.; Jahng, J.W. Post-weaning isolation promotes food intake and body weight gain in rats that experienced neonatal maternal separation. Brain Res. 2009, 1295, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.K.; Simms, J.A.; Bartlett, S.E. Conditioned cues and yohimbine induce reinstatement of beer and near-beer seeking in Long-Evans rats. Addict. Biol. 2009, 14, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Daniels, W.M.; Pietersen, C.Y.; Carstens, M.E.; Stein, D.J. Maternal separation in rats leads to anxiety-like behavior and a blunted ACTH response and altered neurotransmitter levels in response to a subsequent stressor. Metab. Brain Dis. 2004, 19, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Hulshof, H.J.; Novati, A.; Sgoifo, A.; Luiten, P.G.M.; Den Boer, J.A.; Meerlo, P. Maternal separation decreases adult hippocampal cell proliferation and impairs cognitive performance but has little effect on stress sensitivity and anxiety in adult Wistar rats. Behav. Brain Res. 2011, 216, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Rokutan, K. Molecular stress response in the stomach. Nihon Yakurigaku Zasshi 1999, 114, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.D.; Mueller, A.; Sisti, H.M.; Ogawa, S.; Mcewen, B.S.; Brake, W.G. Anxiety and fear behaviors in adult male and female C57BL/6 mice are modulated by maternal separation. Horm. Behav. 2003, 43, 561–567. [Google Scholar] [CrossRef]
- Chatterjee, S.; Steensland, P.; Simms, J.A.; Holgate, J.; Coe, J.W.; Hurst, R.S.; Shaffer, C.L.; Lowe, J.; Rollema, H.; Bartlett, S.E. Partial agonists of the alpha3beta4* neuronal nicotinic acetylcholine receptor reduce ethanol consumption and seeking in rats. Neuropsychopharmacology 2011, 36, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Yang, S.; Wen, D.; Sun, Q.; Li, Y.; Ma, C.; Cong, B. Stress-induced enhancement of fear conditioning activates the amygdalar cholecystokinin system in a rat model of post-traumatic stress disorder. Neuroreport 2014, 25, 1085–1090. [Google Scholar] [CrossRef] [PubMed]
- Golub, Y.; Kaltwasser, S.F.; Mauch, C.P.; Herrmann, L.; Schmidt, U.; Holsboer, F.; Czisch, M.; Wotjak, C.T. Reduced hippocampus volume in the mouse model of Posttraumatic Stress Disorder. J. Psychiatr. Res. 2011, 45, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Rácz, I.; Markert, A.; Mauer, D.; Stoffel-Wagner, B.; Zimmer, A. Long-term ethanol effects on acute stress responses: Modulation by dynorphin. Addict. Biol. 2013, 18, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Liu, T.; Kozlovsky, N.; Kaplan, Z.; Zohar, J.; Mathe, A.A. The neuropeptide Y (NPY)-ergic system is associated with behavioral resilience to stress exposure in an animal model of post-traumatic stress disorder. Neuropsychopharmacology 2012, 37, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Kavushansky, A.; Ben-Shachar, D.; Richter-Levin, G.; Klein, E. Physical stress differs from psychosocial stress in the pattern and time-course of behavioral responses, serum corticosterone and expression of plasticity-related genes in the rat. Stress 2009, 12, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Onnink, A.M.; Zwiers, M.P.; Hoogman, M.; Mostert, J.C.; Kan, C.C.; Buitelaar, J.; Franke, B. Brain alterations in adult ADHD: Effects of gender, treatment and comorbid depression. Eur. Neuropsychopharmacol. 2014, 24, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Kikusui, T.; Faccidomo, S.; Miczek, K.A. Repeated maternal separation: Differences in cocaine-induced behavioral sensitization in adult male and female mice. Psychopharmacology 2005, 178, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Diehl, L.A.; Silveira, P.P.; Leite, M.C.; Crema, L.M.; Portella, A.K.; Billodre, M.N.; Nunes, E.; Henriques, T.P.; Fidelix-Da-Silva, L.B.; Heis, M.D.; et al. Long lasting sex-specific effects upon behavior and S100b levels after maternal separation and exposure to a model of post-traumatic stress disorder in rats. Brain Res. 2007, 1144, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Cutajar, M.C.; Mullen, P.E.; Ogloff, J.R.; Thomas, S.D.; Wells, D.L.; Spataro, J. Psychopathology in a large cohort of sexually abused children followed up to 43 years. Child Abuse Negl. 2010, 34, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.T.; Back, S.E. Childhood trauma, posttraumatic stress disorder, and alcohol dependence. Alcohol Res. 2012, 34, 408–413. [Google Scholar] [PubMed]
- Farrugia, P.L.; Mills, K.L.; Barrett, E.; Back, S.E.; Teesson, M.; Baker, A.; Sannibale, C.; Hopwood, S.; Rosenfeld, J.; Merz, S.; et al. Childhood trauma among individuals with co-morbid substance use and post traumatic stress disorder. Ment. Health Subst. Use 2011, 4, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Chan, P.K.; Verduin, M.L.; Kemp, D.E.; Tolliver, B.K.; Ganocy, S.J.; Bilali, S.; Brady, K.T.; Findling, R.L.; Calabrese, J.R. Independent predictors for lifetime and recent substance use disorders in patients with rapid-cycling bipolar disorder: Focus on anxiety disorders. Am. J. Addict. 2010, 19, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Back, S.E.; Brady, K.T.; Waldrop, A.E.; Yeatts, S.D.; Mcrae, A.L.; Spratt, E. Early life trauma and sensitivity to current life stressors in individuals with and without cocaine dependence. Am. J. Drug Alcohol Abuse 2008, 34, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, A.E.; Santa Ana, E.J.; Saladin, M.E.; Mcrae, A.L.; Brady, K.T. Differences in Early Onset Alcohol Use and Heavy Drinking among Persons with Childhood and Adulthood Trauma. Am. J. Addict. 2007, 16, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Bidzan, L.; Ussorowska, D. Risk factors for dementia of the Alzheimer type. Psychiatr. Pol. 1995, 29, 297–306. [Google Scholar] [PubMed]
- Nylander, I.; Roman, E. Is the rodent maternal separation model a valid and effective model for studies on the early-life impact on ethanol consumption? Psychopharmacology 2013, 229, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Roman, E.; Nylander, I. The impact of emotional stress early in life on adult voluntary ethanol intake-results of maternal separation in rats. Stress 2005, 8, 157–174. [Google Scholar] [CrossRef] [PubMed]
- CDC. Fact Sheets—Excessive Alcohol Use and Risks to Men’s Health. Available online: https://www.cdc.gov/alcohol/fact-sheets/mens-health.htm (accessed on 10 November 2016).
- Martin, L.; Viljoen, M.; Kidd, M.; Seedat, S. Are childhood trauma exposures predictive of anxiety sensitivity in school attending youth? J. Affect. Disord. 2014, 168, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Mukerjee, D.; St George, D.; Knight, C.; Davar, J.; Wells, A.U.; Du Bois, R.M.; Black, C.M.; Coghlan, J.G. Echocardiography and pulmonary function as screening tests for pulmonary arterial hypertension in systemic sclerosis. Rheumatology 2004, 43, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Brioni, J.D.; Esbenshade, T.A.; Garrison, T.R.; Bitner, S.R.; Cowart, M.D. Discovery of histamine H3 antagonists for the treatment of cognitive disorders and Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2011, 336, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Weiss, F.; Zorrilla, E.P. Remission and resurgence of anxiety-like behavior across protracted withdrawal stages in ethanol-dependent rats. Alcohol. Clin. Exp. Res. 2007, 31, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Kushner, M.G.; Thuras, P.; Abrams, K.; Brekke, M.; Stritar, L. Anxiety mediates the association between anxiety sensitivity and coping-related drinking motives in alcoholism treatment patients. Addict. Behav. 2001, 26, 869–885. [Google Scholar] [CrossRef]
- Gilbertson, R.; Frye, R.F.; Nixon, S.J. Nicotine as a factor in stress responsiveness among detoxified alcoholics. Alcohol Alcohol. 2011, 46, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Tarren, J.R.; Lester, H.A.; Belmer, A.; Bartlett, S.E. Acute Ethanol Administration Upregulates Synaptic alpha4-Subunit of Neuronal Nicotinic Acetylcholine Receptors within the Nucleus Accumbens and Amygdala. Front. Mol. Neurosci. 2017, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Kim, J.W.; Song, J.Y.; Park, S.; Lee, H.J.; Chung, J.H. Association of polymorphisms in nicotinic acetylcholine receptor alpha 4 subunit gene (CHRNA4), mu-opioid receptor gene (OPRM1), and ethanol-metabolizing enzyme genes with alcoholism in Korean patients. Alcohol 2004, 34, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Reuter, M.; Markett, S.; Melchers, M.; Montag, C. Interaction of the cholinergic system and the hypothalamic-pituitary-adrenal axis as a risk factor for depression: Evidence from a genetic association study. Neuroreport 2012, 23, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, D.; Rennard, S.I.; Nides, M.; Oncken, C.; Azoulay, S.; Billing, C.B.; Watsky, E.J.; Gong, J.; Williams, K.E.; Reeves, K.R. Varenicline, an alpha4beta2 nicotinic acetylcholine receptor partial agonist, vs sustained-release bupropion and placebo for smoking cessation: A randomized controlled trial. JAMA 2006, 296, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.M.; Teague, C.H.; Kayser, A.S.; Bartlett, S.E.; Fields, H.L. Varenicline decreases alcohol consumption in heavy-drinking smokers. Psychopharmacology 2012, 223, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Steensland, P.; Simms, J.A.; Holgate, J.; Richards, J.K.; Bartlett, S.E. Varenicline, an alpha4beta2 nicotinic acetylcholine receptor partial agonist, selectively decreases ethanol consumption and seeking. Proc. Natl. Acad. Sci. USA 2007, 104, 12518–12523. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Control | MS |
|---|---|---|
| Female | 17.01 ± 0.333 | 17.97 ± 0.294 * |
| Male | 21.64 ± 0.446 | 22.17 ± 0.277 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holgate, J.Y.; Tarren, J.R.; Bartlett, S.E. Sex Specific Alterations in α4*Nicotinic Receptor Expression in the Nucleus Accumbens. Brain Sci. 2018, 8, 70. https://doi.org/10.3390/brainsci8040070
Holgate JY, Tarren JR, Bartlett SE. Sex Specific Alterations in α4*Nicotinic Receptor Expression in the Nucleus Accumbens. Brain Sciences. 2018; 8(4):70. https://doi.org/10.3390/brainsci8040070
Chicago/Turabian StyleHolgate, Joan Y., Josephine R. Tarren, and Selena E. Bartlett. 2018. "Sex Specific Alterations in α4*Nicotinic Receptor Expression in the Nucleus Accumbens" Brain Sciences 8, no. 4: 70. https://doi.org/10.3390/brainsci8040070
APA StyleHolgate, J. Y., Tarren, J. R., & Bartlett, S. E. (2018). Sex Specific Alterations in α4*Nicotinic Receptor Expression in the Nucleus Accumbens. Brain Sciences, 8(4), 70. https://doi.org/10.3390/brainsci8040070