On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior



Abstract
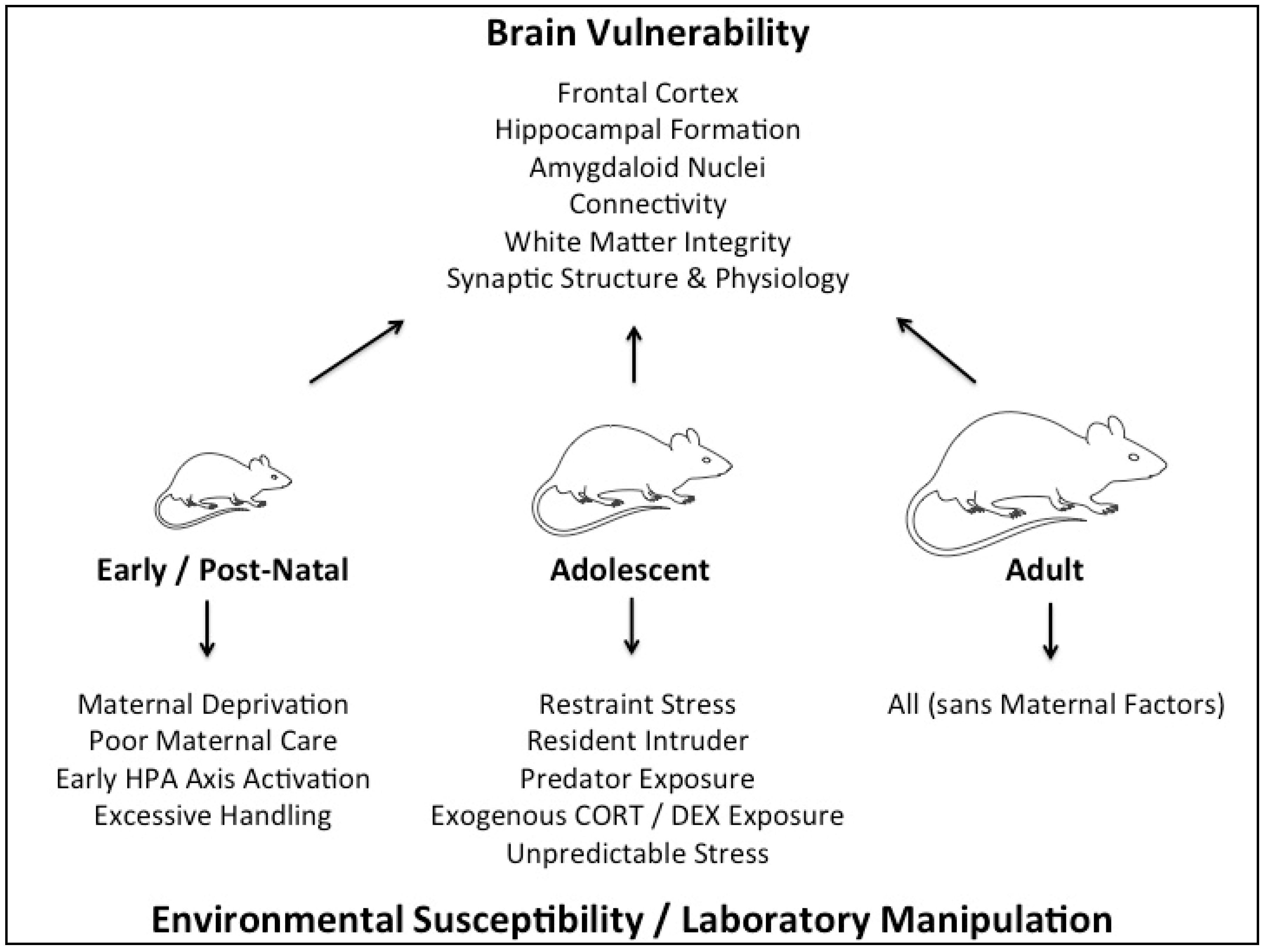
:1. Introduction
2. Early Life Stress Models
Psychological Stress Models in Early Life
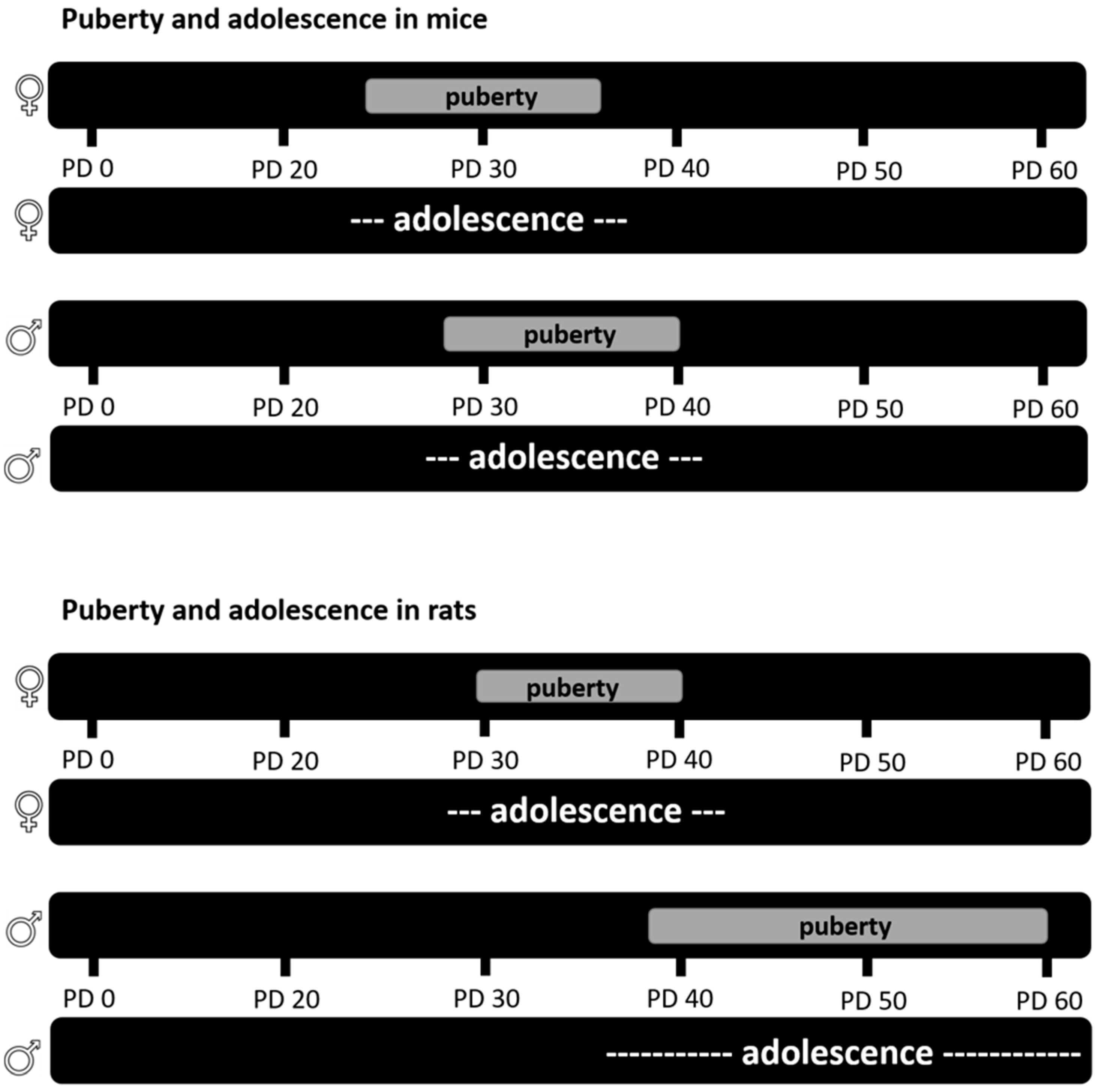
3. Adolescent Stress Models
3.1. Physical and Social Stress during Adolescence
3.2. Pharmacological Stress During Adolescence
4. Adult Stress Models
4.1. Physical and Social Stress in Adulthood
4.2. Pharmacological Stress in Adulthood
5. Genetically Modified Animal Models and the Study of Stress
5.1. Early Insight from Non-Specific Loss- and Gain-of-Function Models
5.2. Insight from Brain-Specific and Forebrain-Restricted Depletion of GR and MR
5.3. Insight from Deficient DNA Binding in GRDim Mice
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bao, A.M.; Meynen, G.; Swaab, D.F. The stress system in depression and neurodegeneration: Focus on the human hypothalamus. Brain Res. Rev. 2008, 57, 531–553. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R.; de Jong, I.E.M.; Oitzi, M.S. Neuropharmacology of glucocorticoids: Focus on emotion, cognition and cocaine. Eur. J. Pharmacol. 2008, 585, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, I.A.; Bennett, T.; Fellows, I.W. Catecholamines and the control of metabolism in man. Clin. Sci. 1985, 68, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Borodovitsyna, O.; Flamini, M.; Chandler, D. Noradrenergic modulation of cognition in health and disease. Neural Plast. 2017, 2017, 6031478. [Google Scholar] [CrossRef] [PubMed]
- McGaughy, J.; Ross, R.S.; Eichenbaum, H. Noradrenergic, but not cholinergic, deafferentation of prefrontal cortex impairs attentional set-shifting. Neuroscience 2008, 153, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.K.; Hillier, A.; Smith, R.M.; Tivarus, M.E.; Beversdorf, D.Q. Beta-adrenergic modulation of cognitive flexibility during stress. J. Cogn. Neurosci. 2007, 19, 468–478. [Google Scholar] [CrossRef] [PubMed]
- McCarty, R.; Horwatt, K.; Konarska, M. Chronic stress and sympathetic-adrenal medullary responsiveness. Soc. Sci. Med. 1988, 26, 333–341. [Google Scholar] [CrossRef]
- Konarska, M.; Stewart, R.E.; McCarty, R. Sensitization of sympathetic-adrenal medullary responses to a novel stressor in chronically stressed laboratory rats. Physiol. Behav. 1989, 46, 129–135. [Google Scholar] [CrossRef]
- Kvetnansky, R.; Pacak, K.; Fukuhara, K.; Viskupic, E.; Hiremagalur, B.; Nankova, B.; Goldstein, D.S.; Sabban, E.L.; Kopin, I.J. Sympathoadrenal system in stress. Interaction with the hypothalamic-pituitary-adrenocortical system. Ann. N. Y. Acad. Sci. 1995, 771, 131–158. [Google Scholar] [PubMed]
- De Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Workman, J.L.; Lee, T.T.; Innala, L.; Viau, V. Sex differences in the HPA axis. Compr. Physiol. 2014, 4, 1121–1155. [Google Scholar] [PubMed]
- Seale, J.V.; Wood, S.A.; Atkinson, H.C.; Bate, E.; Lightman, S.L.; Ingram, C.D.; Jessop, D.S.; Harbuz, M.S. Gonadectomy reverses the sexually diergic patterns of circadian and stress-induced hypothalamic-pituitary-adrenal axis activity in male and female rats. J. Neuroendocrinol. 2004, 16, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.E.; Beck, K.D.; Luine, V.N. Chronic stress effects on memory: Sex differences in performance and monoaminergic activity. Horm. Behav. 2003, 43, 48–59. [Google Scholar] [CrossRef]
- Bekker, M.H.J.; van Mens-Verhulst, J. Anxiety disorders: Sex differences in prevalence, degree, and background, but gender-neutral treatment. Gend. Med. 2007, 4, S178–S193. [Google Scholar] [CrossRef]
- Breslau, N.; Davis, G.C.; Andreski, P.; Peterson, E.L.; Schultz, L.R. Sex differences in posttraumatic stress disorder. Arch. Gen. Psychiatry 1997, 54, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Tolin, D.F.; Foa, E.B. Sex differences in trauma and posttraumatic stress disorder: A quantitative review of 25 years of research. Psychol. Bull. 2006, 132, 959–992. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.C.; Waddell, B.J. Circadian variation in basal plasma corticosterone and adrenocorticotropin in the rat: Sexual dimorphism and changes across the estrous cycle. Endocrinology 1997, 138, 3842–3848. [Google Scholar] [CrossRef] [PubMed]
- Bourke, C.H.; Harrell, C.S.; Neigh, G.N. Stress-induced sex differences: Adaptations mediated by the glucocorticoid receptor. Horm. Behav. 2012, 62, 210–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radley, J.J.; Rocher, A.B.; Miller, M.; Janssen, W.G.; Liston, C.; Hof, P.R.; McEwen, B.S.; Morrison, J.H. Repeated stress induces dendritic spine loss in the rat medial prefrontal cortex. Cereb. Cortex 2006, 16, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, K.A.; van den Hove, D.L.; Schmitz, C.; Segers, O.; Prickaerts, J.; Steinbusch, H.W. Prenatal stress and subsequent exposure to chronic mild stress influence dendritic spine density and morphology in the rat medial prefrontal cortex. BMC Neurosci. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Magariños, A.M.A.; McEwen, B.S.; Flügge, G.; Fuchs, E. Chronic psychosocial stress causes apical dendritic atrophy of hippocampal CA3 pyramidal neurons in subordinate tree shrews. J. Neurosci. 1996, 16, 3534–3540. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Jadhav, S.; McEwen, B.S.; Vyas, A.; Chattarji, S. Stress duration modulates the spatiotemporal patterns of spine formation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA 2005, 102, 9371–9376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, J.; Murmu, M.S.; Biala, Y.; Weinstock, M.; Braun, K. Prenatal stress and neonatal handling induce sex-specific changes in dendritic complexity and dendritic spine density in hippocampal subregions of prepubertal rats. Neuroscience 2011, 193, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Popoli, M.; Yan, Z.; McEwen, B.S.; Sanacora, G. The stressed synapse: The impact of stress and glucocorticoids on glutamate transmission. Nat. Rev. Neurosci. 2011, 13, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Scheggi, S.; Leggio, B.; Masi, F.; Grappi, S.; Gambarana, C.; Nanni, G.; Rauggi, R.; De Montis, M.G. Selective modifications in the nucleus accumbens of dopamine synaptic transmission in rats exposed to chronic stress. J. Neurochem. 2002, 83, 895–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marmigère, F.; Givalois, L.; Rage, F.; Arancibia, S.; Tapia-Arancibia, L. Rapid induction of bdnf expression in the hippocampus during immobilization stress challenge in adult rats. Hippocampus 2003, 13, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Makino, S.; Kvetnansky, R.; Post, R.M. Stress and glucocorticoids affect the expression of brain-derived neurotrophic factor and neurotrophin-3 mRNAs in the hippocampus. J. Neurosci. 1995, 15, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Choy, K.H.C.; de Visser, Y.; Nichols, N.R.; van den Buuse, M. Combined neonatal stress and young-adult glucocorticoid stimulation in rats reduce bdnf expression in hippocampus: Effects on learning and memory. Hippocampus 2008, 18, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Imbe, H.; Morikawa, Y.; Kubo, C.; Senba, E. Chronic stress, as well as acute stress, reduces BDNF mRNA expression in the rat hippocampus but less robustly. Neurosci. Res. 2005, 53, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Brummelte, S.; Galea, L.A. Chronic high corticosterone reduces neurogenesis in the dentate gyrus of adult male and female rats. Neuroscience 2010, 168, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Cameron, H.A.; Gould, E. Adult neurogenesis is regulated by adrenal steroids in the dentate gyrus. Neuroscience 1994, 61, 203–209. [Google Scholar] [CrossRef]
- Crochemore, C.; Lu, J.; Wu, Y.; Liposits, Z.; Sousa, N.; Holsboer, F.; Almeida, O.F. Direct targeting of hippocampal neurons for apoptosis by glucocorticoids is reversible by mineralocorticoid receptor activation. Mol. Psychiatry 2005, 10, 790–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueira, J.J.; Pego, J.M.; Taipa, R.; Bessa, J.M.; Almeida, O.F.; Sousa, N. Morphological correlates of corticosteroid-induced changes in prefrontal cortex-dependent behaviors. J. Neurosci. 2005, 25, 7792–7800. [Google Scholar] [CrossRef] [PubMed]
- Insel, T.; Cuthbert, B.; Garvey, M.; Heinssen, R.; Pine, D.S.; Quinn, K.; Sanislow, C.; Wang, P. Research domain criteria (RDoC): Toward a new classification framework for research on mental disorders. Am. J. Psychiatry 2010, 167, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Berglund, P.; Demler, O.; Jin, R.; Merikangas, K.R.; Walters, E.E. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the national comorbidity survey replication. Arch. Gen. Psychiatry 2005, 62, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Costello, E.J.; Mustillo, S.; Erkanli, A.; Keeler, G.; Angold, A. Prevalence and development of psychiatric disorders in childhood and adolescence. Arch. Gen. Psychiatry 2003, 60, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef] [Green Version]
- Yuen, E.Y.; Wei, J.; Yan, Z. Molecular and epigenetic mechanisms for the complex effects of stress on synaptic physiology and cognitive functions. Int. J. Neuropsychopharmacol. 2017, 20, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Nasca, C.; Davis, E.; Bigio, B.; Sandi, C.; McEwen, B.S. Effects of Stress throughout the Lifespan on the Brain and Behavior; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2017; pp. 443–463. [Google Scholar]
- Aisa, B.; Tordera, R.; Lasheras, B.; Del Rio, J.; Ramirez, M.J. Cognitive impairment associated to HPA axis hyperactivity after maternal separation in rats. Psychoneuroendocrinology 2007, 32, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Oomen, C.A.; Soeters, H.; Audureau, N.; Vermunt, L.; van Hasselt, F.N.; Manders, E.M.M.; Joels, M.; Lucassen, P.J.; Krugers, H. Severe early life stress hampers spatial learning and neurogenesis, but improves hippocampal synaptic plasticity and emotional learning under high-stress conditions in adulthood. J. Neurosci. 2010, 30, 6635–6645. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.L.; Bagot, R.C.; van Hasselt, F.; Ramakers, G.; Meaney, M.J.; de Kloet, E.R.; Joels, M.; Krugers, H. Maternal care and hippocampal plasticity: Evidence for experience-dependent structural plasticity, altered synaptic functioning, and differential responsiveness to glucocorticoids and stress. J. Neurosci. 2008, 28, 6037–6045. [Google Scholar] [CrossRef] [PubMed]
- Suri, D.; Bhattacharya, A.; Vaidya, V.A. Early stress evokes temporally distinct consequences on the hippocampal transcriptome, anxiety and cognitive behaviour. Int. J. Neuropsychopharmacol. 2014, 17, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.S.; Patki, G.; Salvi, A.; Kelly, M.; Salim, S. Behavioral effects of early life maternal trauma witness in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Klug, M.; Von Soly, S.K.; Binder, M.D.; Hannan, A.J.; van den Buuse, M. Sex-specific disruptions in spatial memory and anhedonia in a “two hit” rat model correspond with alterations in hippocampal brain-derived neurotrophic factor expression and signaling. Hippocampus 2014, 24, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, F.E. Sex differences in voluntary drinking by Long Evans rats following early stress. Alcoholism 1998, 22, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Chaby, L.E.; Sheriff, M.J.; Hirrlinger, A.M.; Lim, J.; Fetherston, T.B.; Braithwaite, V.A. Does chronic unpredictable stress during adolescence affect spatial cognition in adulthood? PLoS ONE 2015, 10, e0141908. [Google Scholar] [CrossRef] [PubMed]
- Pyter, L.M.; Kelly, S.D.; Harrell, C.S.; Neigh, G.N. Sex differences in the effects of adolescent stress on adult brain inflammatory markers in rats. Brain Behav. Immun. 2013, 30, 88–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isgor, C.; Kabbaj, M.; Akil, H.; Watson, S.J. Delayed effects of chronic variable stress during peripubertal-juvenile period on hippocampal morphology and on cognitive and stress axis functions in rats. Hippocampus 2004, 14, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Hollis, F.; Isgor, C.; Kabbaj, M. The consequnces of adolescent chronic unpredictable stress exposure on brain behavior. Neuroscience 2013, 249, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Oztan, O.; Aydin, C.; Isgor, C. Chronic variable physical stress during the peripubertal-juvenile period causes differential depressive and anxiogenic effects in the novelty-seeking phenotype: Functional implications for hippocampal and amygdalar brain-derived neurotrophic factor and the mossy fibre plasticity. Neuroscience 2011, 192, 334–344. [Google Scholar] [PubMed]
- Conrad, C.D.; McLaughlin, K.J.; Harman, J.S.; Foltz, C.; Wieczorek, L.; Lightner, E.; Wright, R.L. Chronic glucocorticoids increase hippocampal vulnerability to neurotoxicity under conditions that produce CA3 dendritic retraction but fail to impair spatial recognition memory. J. Neurosci. 2007, 27, 8278–8285. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.J.; Crowley, N.A.; Reiss, D.E.; Caulfield, J.I.; Luscher, B.; Cavigelli, S.A.; Kamens, H.M. Adolescent social stress increases anxiety-like behavior and alters synaptic transmission, without influencing nicotine responses, in a sex-dependent manner. Neuroscience 2018, 373, 182–198. [Google Scholar] [CrossRef] [PubMed]
- Sterlemann, V.; Rammes, G.; Wolf, M.; Liebl, C.; Ganea, K.; Muller, M.B.; Schmidt, M.V. Chronic social stress during adolescence induces cognitive impairment in aged mice. Hippocampus 2010, 20, 540–549. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Thomas, C.M.; Sheridan, C.S.; Nixon, F.; Flynn, J.A.; Mathews, I.Z. Social instability stress in adolescent male rats alters hippocampal neurogenesis and produces deficits in spatial location memory in adulthood. Hippocampus 2012, 22, 1300–1312. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Mathews, I.Z.; Thomas, C.; Waters, P. Investigations of HPA function and the enduring consequences of stressors in adolescence in animal models. Brain Cogn. 2010, 72, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, M.D.; Mathews, I.Z.; McCormick, C.M. Enduring deficits in contextual and auditory fear conditioning after adolescent, not adult, social instability stress in male rats. Neurobiol. Learn. Mem. 2011, 95, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.; Bureta, L.; Hill, R.A.; van den Buuse, M. Gene-environment interaction of reelin and stress in cognitive behaviours in mice: Implications for schizophrenia. Behav. Brain Res. 2015, 287, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Tannenbaum, B.; Tannenbaum, G.S.; Sudom, K.; Anisman, H. Neurochemical and behavioral alterations elicited by a chronic intermittent stressor regimen: Implications for allostatic load. Brain Res. 2002, 953, 82–92. [Google Scholar] [CrossRef]
- Crema, L.; Schlabitz, M.; Tagliari, B.; Cunha, A.; Simao, F.; Krolow, R.; Pettenuzzo, L.; Salbego, C.; Vendite, D.; Wyse, A.T.; et al. Na+, K+ ATPase activity is reduced in amygdala of rats with chronic stress-induced anxiety-like behavior. Neurochem. Res. 2010, 35, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Tuthill, D.; Mufson, R.A.; Shi, Y. Chronic restraint stress promotes lymphocyte apoptosis by modulating CD95 expression. J. Exp. Med. 2000, 191, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Kim, B.H.; Ye, S.K.; Kim, M.H. Chronic non-social stress affects depressive behaviors but not anxiety in mice. Korean J. Physiol. Pharmacol. 2014, 18, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Trofimiuk, E.; Walesiuk, A.; Braszko, J.J. St John’s wort (hypericum perforatum) diminishes cognitive impairment caused by the chronic restraint stress in rats. Pharmacol. Res. 2005, 51, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- D’Aquila, P.S.; Brain, P.; Willner, P. Effects of chronic mild stress on performance in behavioural tests relevant to anxiety and depression. Physiol. Behav. 1994, 56, 861–867. [Google Scholar] [CrossRef]
- Matuszewich, L.; Karney, J.J.; Carter, S.R.; Janasik, S.P.; O’Brien, J.L.; Friedman, R.D. The delayed effects of chronic unpredictable stress on anxiety measures. Physiol. Behav. 2007, 90, 674–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Shi, R.; Wang, J.; Wang, J.F.; Li, X.M. Unpredictable chronic mild stress not chronic restraint stress induces depressive behaviours in mice. Neuroreport 2014, 25, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.O.; Duarte, J.O.; Costa-Ferreira, W.; Morais-Silva, G.; Marin, M.T.; Crestani, C.C. Sex differences in cardiovascular, neuroendocrine and behavioral changes evoked by chronic stressors in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.E.; Zrull, M.C.; Luine, V.N. Chronic restraint stress enhances radial arm maze performance in female rats. Brain Res. 2001, 904, 279–289. [Google Scholar] [CrossRef]
- Luine, V.; Villegas, M.; Martinez, C.; McEwen, B.S. Repeated stress causes reversible impairments of spatial memory performance. Brain Res. 1994, 639, 167–170. [Google Scholar] [CrossRef]
- Patki, G.; Solanki, N.; Atrooz, F.; Allam, F.; Salim, S. Depression, anxiety-like behavior and memory impairment are associated with increased oxidative stress and inflammation in a rat model of social stress. Brain Res. 2013, 1539, 73–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, V.; Allaili, N.; Euvrard, M.; Marday, T.; Riffaud, A.; Franc, B.; Mocaer, E.; Gabriel, C.; Fossati, P.; Lehericy, S.; et al. Effect of agomelatine on memory deficits and hippocampal gene expression induced by chronic social defeat stress in mice. Sci. Rep. 2017, 7, 45907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.D.; Chen, Y.C.; Wolf, M.; Wagner, K.V.; Liebl, C.; Scharf, S.H.; Harbich, D.; Mayer, B.; Wurst, W.; Holsboer, F.; et al. Forebrain CRHR1 deficiency attenuates chronic stress-induced cognitive deficits and dendritic remodeling. Neurobiol. Dis. 2011, 42, 300–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonken, L.K.; Finy, M.S.; Walton, J.C.; Weil, Z.M.; Workman, J.L.; Ross, J.; Nelson, R.J. Influence of light at night on murine anxiety- and depressive-like responses. Behav. Brain Res. 2009, 205, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Kitsmiller, E.; Smale, L.; Nelson, R.J. Dim nighttime light impairs cognition and provokes depressive-like responses in a diurnal rodent. J. Biol. Rhythm. 2012, 27, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.P.; Frussa-Filho, R.; Fukushiro, D.F.; Chinen, C.C.; Abilio, V.C.; Silva, R.H. Effects of long-term continuous exposure to light on memory and anxiety in mice. Physiol. Behav. 2005, 86, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Craig, L.A.; McDonald, R.J. Chronic disruption of circadian rhythms impairs hippocampal memory in the rat. Brain Res. Bull. 2008, 76, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Van Donkelaar, E.L.; Vaessen, K.R.; Pawluski, J.L.; Sierksma, A.S.; Blokland, A.; Canete, R.; Steinbusch, H.W. Long-term corticosterone exposure decreases insulin sensitivity and induces depressive-like behaviour in the C57BL/6NCrl mouse. PLoS ONE 2014, 9, e106960. [Google Scholar] [CrossRef] [PubMed]
- Wong, E.Y.; Herbert, J. Raised circulating corticosterone inhibits neuronal differentiation of progenitor cells in the adult hippocampus. Neuroscience 2006, 137, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M.; Uno, H.; Rebert, C.S.; Finch, C.E. Hippocampal damage associated with prolonged glucocorticoid exposure in primates. J. Neurosci. 1990, 10, 2897–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skorzewska, A.; Lehner, M.; Wislowska-Stanek, A.; Krzascik, P.; Ziemba, A.; Plaznik, A. The effect of chronic administration of corticosterone on anxiety- and depression-like behavior and the expression of GABA-A receptor alpha-2 subunits in brain structures of low- and high-anxiety rats. Horm. Behav. 2014, 65, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Andres, S.; Cardenas, S.; Parra, C.; Bravo, J.; Greiner, M.; Rojas, P.; Morales, P.; Lara, H.; Fiedler, J. Effects of long-term adrenalectomy on apoptosis and neuroprotection in the rat hippocampus. Endocrine 2006, 29, 299–307. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, H.; Xu, C.; Yang, G.; Tao, J.; Huang, J.; Wu, J.; Duan, X.; Cao, Y.; Dong, J. Neuroprotective effects of icariin on corticosterone-induced apoptosis in primary cultured rat hippocampal neurons. Brain Res. 2011, 1375, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.; Becker, A.; Schroeder, A.; Bilkei-Gorzo, A.; Zimmer, A. Effect of chronic corticosterone application on depression-like behavior in C57BL/6N and C57BL/6J mice. Genes Brain Behav. 2015, 14, 292–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekiri, M.; Gardier, A.M.; David, D.J.; Guilloux, J.P. Chronic Corticosterone Administration Effects on Behavioral Emotionality in Female C57BL6 Mice. Exp. Clin. Psychopharmacol. 2017, 25, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Glover, V.; O’Connor, T.G.; O’Donnell, K. Prenatal stress and the programming of the HPA axis. Neurosci. Biobehav. Rev. 2010, 35, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Abbott, P.W.; Gumusoglu, S.B.; Bittle, J.; Beversdorf, D.Q.; Stevens, H.E. Prenatal stress and genetic risk: How prenatal stress interacts with genetics to alter risk for psychiatric illness. Psychoneuroendocrinology 2018, 90, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Dunn, E.; Kostaki, A.; Andrews, M.H.; Matthews, S.G. Fetal programming of hypothalamo-pituitary-adrenal function: Prenatal stress and glucocorticoids. J. Physiol. 2006, 572, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Pechtel, P.; Pizzagalli, D.A. Effects of early life stress on cognitive and affective function: An integrated review of human literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Brain on stress: How the social environment gets under the skin (vol 109, pg 17180, 2012). Proc. Natl. Acad. Sci. USA 2013, 110, 1561. [Google Scholar] [CrossRef] [PubMed]
- Hackman, D.A.; Farah, M.J.; Meaney, M.J. Science and society socioeconomic status and the brain: Mechanistic insights from human and animal research. Nat. Rev. Neurosci. 2010, 11, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Van Harmelen, A.-L.; van Tol, M.-J.; Demenescu, L.R.; van der Wee, N.J.A.; Veltman, D.J.; Aleman, A.; van Buchem, M.A.; Spinhoven, P.; Penninx, B.W.J.H.; Elzinga, B.M. Enhanced amygdala reactivity to emotional faces in adults reporting childhood emotional maltreatment. Soc. Cogn. Affect. Neurosci. 2013, 8, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Carr, C.P.; Severi Martins, C.M.; Stingel, A.M.; Lemgruber, V.B.; Juruena, M.F. The role of early life stress in adult psychiatric disorders a systematic review according to childhood trauma subtypes. J. Nerv. Ment. Dis. 2013, 201, 1007–1020. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Feldon, J. Long-term biobehavioral effects of maternal separation in the rat: Consistent or confusing? Rev. Neurosci. 2000, 11, 383–408. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.A.; Turkka, J.; Lister, R.G.; Linnoila, M. Effects of early postnatal handling on brain beta-adrenoceptors and behavior in tests related to stress. Brain Res. 1991, 542, 286–292. [Google Scholar] [CrossRef]
- Suri, D.; Veenit, V.; Sarkar, A.; Thiagarajan, D.; Kumar, A.; Nestler, E.J.; Galande, S.; Vaidya, V.A. Early stress evokes age-dependent biphasic changes in hippocampal neurogenesis, BDNF expression, and cognition. Biol. Psychiatry 2013, 73, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Wiener, S.G.; Levine, S. Perinatal malnutrition and early handling: Interactive effects on the development of the pituitary-adrenal system. Dev. Psychobiol. 1978, 11, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Wiener, S.G.; Levine, S. Influence of perinatal malnutrition and early handling on the pituitary-adrenal response to noxious stimuli in adult rats. Physiol. Behav. 1983, 31, 285–291. [Google Scholar] [CrossRef]
- Schmidt, M.V.; Oitzl, M.S.; Levine, S.; de Kloet, E.R. The HPA system during the postnatal development of CD1 mice and the effects of maternal deprivation. Dev. Brain Res. 2002, 139, 39–49. [Google Scholar] [CrossRef]
- Lehmann, J.; Russig, H.; Feldon, J.; Pryce, C.R. Effect of a single maternal separation at different pup ages on the corticosterone stress response in adult and aged rats. Pharmacol. Biochem. Behav. 2002, 73, 141–145. [Google Scholar] [CrossRef]
- Plotsky, P.M.; Meaney, M.J. Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Mol. Brain Res. 1993, 18, 195–200. [Google Scholar] [CrossRef]
- Silveira, P.P.; Portella, A.K.; Benetti, C.D.; Zugno, A.I.; Scherer, E.B.D.; Mattos, C.B.; Wyse, A.T.S.; Lucion, A.B.; Dalmaz, C. Association between Na+,K+-atpase activity and the vulnerability/resilience to mood disorders induced by early life experience. Neurochem. Res. 2011, 36, 2075–2082. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; Parent, S.; Evans, A.C.; Tremblay, R.E.; Zelazo, P.D.; Corbo, V.; Pruessner, J.C.; Seguin, J.R. Larger amygdala but no change in hippocampal volume in 10-year-old children exposed to maternal depressive symptomatology since birth. Proc. Natl. Acad. Sci. USA 2011, 108, 14324–14329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Harmelen, A.L.; de Jong, P.J.; Glashouwer, K.A.; Spinhoven, P.; Penninx, B.; Elzinga, B.M. Child abuse and negative explicit and automatic self-associations: The cognitive scars of emotional maltreatment. Behav. Res. Ther. 2010, 48, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Perez-Cerezales, S.; Ramos-Ibeas, P.; Rizos, D.; Lonergan, P.; Bermejo-Alvarez, P.; Gutierrez-Adan, A. Early sex-dependent differences in response to environmental stress. Reproduction 2018, 155, R39–R51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Xu, B.J.; Lu, B.; Hempstead, B.L. New insights in the biology of BDNF synthesis and release: Implications in CNS function. J. Neurosci. 2009, 29, 12764–12767. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Hyde, T.M.; Cassano, H.L.; Deep-Soboslay, A.; Kleinman, J.E.; Weickert, C.S. Promoter specific alterations of brain-derived neurotrophic factor mRNA in schizophrenia. Neuroscience 2010, 169, 1071–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Kim, Y.K. Reduced platelet bdnf level in patients with major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Mayer, E.A.; Acosta, J.R.; Hamadani, K.; Torgerson, C.; van Horn, J.D.; Chang, L.; Naliboff, B.; Tillisch, K.; Labus, J.S. Early adverse life events are associated with altered brain network architecture in a sex-dependent manner. Neurobiol. Stress 2017, 7, 16–26. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, J.E.; Marinelli, M. Age matters. Eur. J. Neurosci. 2009, 29, 997–1014. [Google Scholar] [CrossRef] [PubMed]
- Vanderkolk, B.A. Adolescent vulnerability to posttraumatic stress disorder Psychiatry-Interpers. Biol. Process. 1985, 48, 365–370. [Google Scholar]
- Morrison, K.E.; Epperson, C.N.; Sammel, M.D.; Ewing, G.; Podcasy, J.S.; Hantsoo, L.; Kim, D.R.; Bale, T.L. Preadolescent adversity programs a disrupted maternal stress reactivity in humans and mice. Biol. Psychiatry 2017, 81, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L. Epigenetic and transgenerational reprogramming of brain development. Nat. Rev. Neurosci. 2015, 16, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L. Lifetime stress experience: Transgenerational epigenetics and germ cell programming. Dialogues Clin. Neurosci. 2014, 16, 297–305. [Google Scholar] [PubMed]
- Roberts, A.L.; Lyall, K.; Rich-Edwards, J.W.; Ascherio, A.; Weisskopf, M.G. Association of maternal exposure to childhood abuse with elevated risk for autism in offspring. JAMA Psychiatry 2013, 70, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.L.; Liew, Z.; Lyall, K.; Ascherio, A.; Weisskopf, M.G. Association of maternal exposure to childhood abuse with elevated risk for attention deficit hyperactivity disorder in offspring. Am. J. Epidemiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.D. Pubertal maturation and programming of hypothalamic-pituitary-adrenal reactivity. Front. Neuroendocrinol. 2010, 31, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Cyranowski, J.M.; Frank, E.; Young, E.; Shear, M.K. Adolescent onset of the gender difference in lifetime rates of major depression: A theoretical model. Arch. Gen. Psychiatry 2000, 57, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Romeo, R.D.; Patel, R.; Pham, L.; So, V.M. Adolescence and the ontogeny of the hormonal stress response in male and female rats and mice. Neurosci. Biobehav. Rev. 2016, 70, 206–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakemore, S.-J.; Burnett, S.; Dahl, R.E. The role of puberty in the developing adolescent brain. Hum. Brain Mapp. 2010, 31, 926–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.; Schoemig, E.; Leweke, F.M. Acute and chronic cannabinoid treatment differentially affects recognition memory and social behavior in pubertal and adult rats. Addict. Biol. 2008, 13, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Fernandez, R.; Navarro, V.M.; Barreiro, M.L.; Vigo, E.M.; Tovar, S.; Sirotkin, A.V.; Casanueva, F.F.; Aguilar, E.; Dieguez, C.; Pinilla, L.; et al. Effects of chronic hyperghrelinemia on puberty onset and pregnancy outcome in the rat. Endocrinology 2005, 146, 3018–3025. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M. Adolescence as a vulnerable period to alter rodent behavior. Cell Tissue Res. 2013, 354, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Adamec, R.E.; Shallow, T. Lasting effects on rodent anxiety of a single exposure to a cat. Physiol. Behav. 1993, 54, 101–109. [Google Scholar] [CrossRef]
- Adamec, R. Transmitter systems involved in neural plasticity underlying increased anxiety and defense—Implications for understanding anxiety following traumatic stress. Neurosci. Biobehav. Rev. 1997, 21, 755–765. [Google Scholar] [CrossRef]
- Blanchard, D.C.; Blanchard, R.J. Ethoexperimental approaches to the biology of emotion. Annu. Rev. Psychol. 1988, 39, 43–68. [Google Scholar] [CrossRef] [PubMed]
- Cantor, C. Post-traumatic stress disorder: Evolutionary perspectives. Aust. N. Z. J. Psychiatry 2009, 43, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Davis, C.G.; Kendler, K.S. Childhood adversity and adult psychiatric disorder in the US National Comorbidity Survey. Psychol. Med. 1997, 27, 1101–1119. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Nixon, F.; Thomas, C.; Lowie, B.; Dyck, J. Hippocampal cell proliferation and spatial memory performance after social instability stress in adolescence in female rats. Behav. Brain Res. 2010, 208, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.H.; Raison, C.L. The role of inflammation in depression: From evolutionary imperative to modern treatment target. Nat. Rev. Immunol. 2016, 16, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.A. Oscillations and hippocampal-prefrontal synchrony. Curr. Opin. Neurobiol. 2011, 21, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Oitzl, M.S.; Champagne, D.L.; van der Veen, R.; de Kloet, E.R. Brain development under stress: Hypotheses of glucocorticoid actions revisited. Neurosci. Biobehav. Rev. 2010, 34, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, N.J.; Squire, L.R.; Clark, R.E. Spatial memory, recognition memory, and the hippocampus. Proc. Natl. Acad. Sci. USA 2004, 101, 14515–14520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braff, D.L.; Geyer, M.A. Sensorimotor gating and schizophrenia. Human and animal model studies. Arch. Gen. Psychiatry 1990, 47, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Klug, M.; Hill, R.A.; Choy, K.H.C.; Kyrios, M.; Hannan, A.J.; van den Buuse, M. Long-term behavioral and NMDA receptor effects of young-adult corticosterone treatment in BDNF heterozygous mice. Neurobiol. Dis. 2012, 46, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Von Soly, S.K.; Ratnayake, U.; Klug, M.; Binder, M.D.; Hannan, A.J.; van den Buuse, M. Long-term effects of combined neonatal and adolescent stress on brain-derived neurotrophic factor and dopamine receptor expression in the rat forebrain. Biochim. Biophys. Acta-Mol. Basis Dis. 2014, 1842, 2126–2135. [Google Scholar] [CrossRef] [PubMed]
- McCormick, C.M.; Mathews, I.Z. Adolescent development, hypothalamic-pituitary-adrenal function, and programming of adult learning and memory. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 756–765. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.C.; Fogaca, M.V.; Aguiar, D.C.; Guimaraes, F.S. Animal models of anxiety disorders and stress. Rev. Bras. Psiquiatr. 2013, 35, S101–S111. [Google Scholar] [CrossRef] [PubMed]
- Hedges, D.W.; Woon, F.L. Early-life stress and cognitive outcome. Psychopharmacology 2011, 214, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sommershof, A.; Aichinger, H.; Engler, H.; Adenauer, H.; Catani, C.; Boneberg, E.M.; Elbert, T.; Groettrup, M.; Kolassa, I.T. Substantial reduction of naive and regulatory T cells following traumatic stress. Brain Behav. Immun. 2009, 23, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.H.; Hong, S.I.; Ma, S.X.; Hwang, J.Y.; Kim, J.S.; Lee, J.H.; Seo, J.Y.; Lee, S.Y.; Jang, C.G. Strain differences in the chronic mild stress animal model of depression and anxiety in mice. Biomol. Ther. 2014, 22, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Troop, N.A.; Hiskey, S. Social defeat and PTSD symptoms following trauma. Br. J. Clin. Psychol. 2013, 52, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, L.K.; Nakashima, B.R.; Hong, H.; Watanabe, K. The smell of danger: A behavioral and neural analysis of predator odor-induced fear. Neurosci. Biobehav. Rev. 2005, 29, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Campeau, S.; Nyhuis, T.J.; Sasse, S.K.; Day, H.E.; Masini, C.V. Acute and chronic effects of ferret odor exposure in sprague-dawley rats. Neurosci. Biobehav. Rev. 2008, 32, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, L.K. Olfactory systems and neural circuits that modulate predator odor fear. Front. Behav. Neurosci. 2014, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, I.S. The effects of predator odors in mammalian prey species: A review of field and laboratory studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [Google Scholar] [CrossRef] [PubMed]
- Adamec, R.E.; Burton, P.; Shallow, T.; Budgell, J. Nmda receptors mediate lasting increases in anxiety-like behavior produced by the stress of predator exposure--implications for anxiety associated with posttraumatic stress disorder. Physiol. Behav. 1999, 65, 723–737. [Google Scholar] [CrossRef]
- Campos, A.C.; Ferreira, F.R.; da Silva, W.A., Jr.; Guimaraes, F.S. Predator threat stress promotes long lasting anxiety-like behaviors and modulates synaptophysin and CB1 receptors expression in brain areas associated with PTSD symptoms. Neurosci. Lett. 2013, 533, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Staples, L.G.; McGregor, I.S.; Hunt, G.E. Long-lasting FosB/DeltaFosB immunoreactivity in the rat brain after repeated cat odor exposure. Neurosci. Lett. 2009, 462, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Janitzky, K.; Peine, A.; Krober, A.; Yanagawa, Y.; Schwegler, H.; Roskoden, T. Increased CRF mRNA expression in the sexually dimorphic BNST of male but not female GAD67 mice and TMT predator odor stress effects upon spatial memory retrieval. Behav. Brain Res. 2014, 272, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Jagannath, A.; Peirson, S.N.; Foster, R.G. Sleep and circadian rhythm disruption in neuropsychiatric illness. Curr. Opin. Neurobiol. 2013, 23, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M. Glucocorticoid toxicity in the hippocampus. Temporal aspects of synergy with kainic acid. Neuroendocrinology 1986, 43, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.C.; Wellman, C.L. Chronic stress alters dendritic morphology in rat medial prefrontal cortex. J. Neurobiol. 2004, 60, 236–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauer, D.; Weis, F.; Krauseneck, T.; Vogeser, M.; Schelling, G.; Roozendaal, B. Traumatic memories, post-traumatic stress disorder and serum cortisol levels in long-term survivors of the acute respiratory distress syndrome. Brain Res. 2009, 1293, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Aerni, A.; Traber, R.; Hock, C.; Roozendaal, B.; Schelling, G.; Papassotiropoulos, A.; Nitsch, R.M.; Schnyder, U.; de Quervain, D.J. Low-dose cortisol for symptoms of posttraumatic stress disorder. Am. J. Psychiatry 2004, 161, 1488–1490. [Google Scholar] [CrossRef] [PubMed]
- Zohar, J.; Yahalom, H.; Kozlovsky, N.; Cwikel-Hamzany, S.; Matar, M.A.; Kaplan, Z.; Yehuda, R.; Cohen, H. High dose hydrocortisone immediately after trauma may alter the trajectory of PTSD: Interplay between clinical and animal studies. Eur. Neuropsychopharmacol. 2011, 21, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Smerin, S.E.; Zhang, L.; Xing, G.; Li, X.; Benedek, D.; Ursano, R.; Li, H. Corticosterone mitigates the stress response in an animal model of PTSD. J. Psychiatr. Res. 2015, 60, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Deppermann, S.; Storchak, H.; Fallgatter, A.J.; Ehlis, A.C. Stress-induced neuroplasticity: (Mal) adaptation to adverse life events in patients with PTSD—A critical overview. Neuroscience 2014, 283, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Way, B.M.; Taylor, S.E. The serotonin transporter promoter polymorphism is associated with cortisol response to psychosocial stress. Biol. Psychiatry 2010, 67, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Wankerl, M.; Stalder, T.; Kirschbaum, C.; Alexander, N. The serotonin transporter gene-linked polymorphic region (5-HTTLPR) and cortisol stress reactivity: A meta-analysis. Mol. Psychiatry 2013, 18, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; Hariri, A.R.; Holmes, A.; Uher, R.; Moffitt, T.E. Genetic sensitivity to the environment: The case of the serotonin transporter gene and its implications for studying complex diseases and traits. Am. J. Psychiatry 2010, 167, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.S.; Kumsta, R.; von Dawans, B.; Monakhov, M.; Ebstein, R.P.; Heinrichs, M. Common oxytocin receptor gene (OXTR) polymorphism and social support interact to reduce stress in humans. Proc. Natl. Acad. Sci. USA 2011, 108, 19937–19942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belay, H.; Burton, C.L.; Lovic, V.; Meaney, M.J.; Sokolowski, M.; Fleming, A.S. Early adversity and serotonin transporter genotype interact with hippocampal glucocorticoid receptor mRNA expression, corticosterone, and behavior in adult male rats. Behav. Neurosci. 2011, 125, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Petersson, M.; Uvnas-Moberg, K. Systemic oxytocin treatment modulates glucocorticoid and mineralocorticoid receptor mRNA in the rat hippocampus. Neurosci. Lett. 2003, 343, 97–100. [Google Scholar] [CrossRef]
- Oitzl, M.S.; Kloet, E.R.; Joëls, M.; Schmid, W.; Cole, T.J. Spatial learning deficits in mice with a targeted glucocorticoid receptor gene disruption. Eur. J. Neurosci. 1997, 9, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Kolber, B.J.; Wieczorek, L.; Muglia, L.J. Hypothalamic-pituitary-adrenal axis dysregulation and behavioral analysis of mouse mutants with altered glucocorticoid or mineralocorticoid receptor function: Review. Stress Int. J. Biol. Stress 2008, 11, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Blendy, J.A.; Monaghan, A.P.; Krieglstein, K.; Schmid, W.; Aguzzi, A.; Fantuzzi, G.; Hummler, E.; Unsicker, K.; Schütz, G. Targeted disruption of the glucocorticoid receptor gene blocks adrenergic chromaffin cell development and severely retards lung maturation. Genes Dev. 1995, 9, 1608–1621. [Google Scholar] [CrossRef] [PubMed]
- Berger, S.; Bleich, M.; Schmid, W.; Cole, T.J.; Peters, J.; Watanabe, H.; Kriz, W.; Warth, R.; Greger, R.; Schütz, G. Mineralocorticoid receptor knockout mice: Pathophysiology of Na+ metabolism. Proc. Natl. Acad. Sci. USA 1998, 95, 9424–9429. [Google Scholar] [CrossRef] [PubMed]
- Gass, P.; Kretz, O.; Wolfer, D.P.; Berger, S.; Tronche, F.; Reichardt, H.M.; Kellendonk, C.; Lipp, H.P.; Schmid, W.; Schütz, G. Genetic disruption of mineralocorticoid receptor leads to impaired neurogenesis and granule cell degeneration in the hippocampus of adult mice. EMBO Rep. 2000, 1, 447–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz-Baldes, A.; Berger, S.; Grahammer, F.; Warth, R.; Goldschmidt, I.; Peters, J.; Schütz, G.; Greger, R.; Bleich, M. Induction of the epithelial Na+ channel via glucocorticoids in mineralocorticoid receptor knockout mice. Pflügers Arch. 2001, 443, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Myles, K.; Purton, J.F.; Brereton, P.S.; Solomon, N.M.; Godfrey, D.I.; Funder, J.W. Grko mice express an aberrant dexamethasone-binding glucocorticoid receptor, but are profoundly glucocorticoid resistant. Mol. Cell. Endocrinol. 2001, 173, 193–202. [Google Scholar] [CrossRef]
- Ridder, S.; Chourbaji, S.; Hellweg, R.; Urani, A.; Zacher, C.; Schmid, W.; Zink, M.; Hörtnagl, H.; Flor, H.; Henn, F.A. Mice with genetically altered glucocorticoid receptor expression show altered sensitivity for stress-induced depressive reactions. J. Neurosci. 2005, 25, 6243–6250. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, H.M.; Umland, T.; Bauer, A.; Kretz, O.; Schütz, G. Mice with an increased glucocorticoid receptor gene dosage show enhanced resistance to stress and endotoxic shock. Mol. Cell. Biol. 2000, 20, 9009–9017. [Google Scholar] [CrossRef] [PubMed]
- Pepin, M.; Pothier, F.; Barden, N. Antidepressant drug action in a transgenic mouse model of the endocrine changes seen in depression. Mol. Pharmacol. 1992, 42, 991–995. [Google Scholar] [PubMed]
- Froger, N.; Palazzo, E.; Boni, C.; Hanoun, N.; Saurini, F.; Joubert, C.; Dutriez-Casteloot, I.; Enache, M.; Maccari, S.; Barden, N. Neurochemical and behavioral alterations in glucocorticoid receptor-impaired transgenic mice after chronic mild stress. J. Neurosci. 2004, 24, 2787–2796. [Google Scholar] [CrossRef] [PubMed]
- Tronche, F.; Kellendonk, C.; Kretz, O.; Gass, P.; Anlag, K.; Orban, P.C.; Bock, R.; Klein, R.; Schütz, G. Disruption of the glucocorticoid receptor gene in the nervous system results in reduced anxiety. Nat. Genet. 1999, 23, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Deroche-Gamonet, V.; Sillaber, I.; Aouizerate, B.; Izawa, R.; Jaber, M.; Ghozland, S.; Kellendonk, C.; Le Moal, M.; Spanagel, R.; Schütz, G. The glucocorticoid receptor as a potential target to reduce cocaine abuse. J. Neurosci. 2003, 23, 4785–4790. [Google Scholar] [CrossRef] [PubMed]
- Revest, J.-M.; Kaouane, N.; Mondin, M.; Le Roux, A.; Rougé-Pont, F.; Vallée, M.; Barik, J.; Tronche, F.; Desmedt, A.; Piazza, P. The enhancement of stress-related memory by glucocorticoids depends on synapsin-Ia/Ib. Mol. Psychiatry 2010, 15, 1140–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revest, J.-M.; Di Blasi, F.; Kitchener, P.; Rougé-Pont, F.; Desmedt, A.; Turiault, M.; Tronche, F.; Piazza, P.V. The MAPK pathway and Egr-1 mediate stress-related behavioral effects of glucocorticoids. Nat. Neurosci. 2005, 8, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.P.; Brewer, J.A.; Funatsu, M.; Wozniak, D.F.; Tsien, J.Z.; Izumi, Y.; Muglia, L.J. Acquired deficit of forebrain glucocorticoid receptor produces depression-like changes in adrenal axis regulation and behavior. Proc. Natl. Acad. Sci. USA 2005, 102, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Solomon, M.B.; Furay, A.R.; Jones, K.; Packard, A.E.; Packard, B.A.; Wulsin, A.C.; Herman, J.P. Deletion of forebrain glucocorticoid receptors impairs neuroendocrine stress responses and induces depression-like behavior in males but not females. Neuroscience 2012, 203, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, M.Y.; Hussain, R.J.; Zampi, M.E.; Sheeran, K.; Solomon, M.B.; Herman, J.P.; Khan, A.; Jacobson, L. Sensitivity of depression-like behavior to glucocorticoids and antidepressants is independent of forebrain glucocorticoid receptors. Brain Res. 2013, 1525, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, S.; Wolfer, D.P.; Selbach, O.; Alter, H.; Erdmann, G.; Reichardt, H.M.; Chepkova, A.N.; Welzl, H.; Haas, H.L.; Lipp, H.-P. Loss of the limbic mineralocorticoid receptor impairs behavioral plasticity. Proc. Natl. Acad. Sci. USA 2006, 103, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, J.; Van Der Mark, M.; Arp, M.; Berger, S.; De Kloet, E.; Oitzl, M. Stress or no stress: Mineralocorticoid receptors in the forebrain regulate behavioral adaptation. Neurobiol. Learn. Mem. 2012, 98, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, J.P.; Kentrop, J.; Arp, M.; Hubens, C.J.; de Kloet, E.R.; Oitzl, M.S. Spatial learning of female mice: A role of the mineralocorticoid receptor during stress and the estrous cycle. Front. Behav. Neurosci. 2013, 7, 56. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, H.M.; Kaestner, K.H.; Tuckermann, J.; Kretz, O.; Wessely, O.; Bock, R.; Gass, P.; Schmid, W.; Herrlich, P.; Angel, P. DNA binding of the glucocorticoid receptor is not essential for survival. Cell 1998, 93, 531–541. [Google Scholar] [CrossRef]
- Heck, S.; Kullmann, M.; Gast, A.; Ponta, H.; Rahmsdorf, H.; Herrlich, P.; Cato, A. A distinct modulating domain in glucocorticoid receptor monomers in the repression of activity of the transcription factor AP-1. EMBO J. 1994, 13, 4087–4095. [Google Scholar] [PubMed]
- Oitzl, M.S.; Reichardt, H.M.; Joëls, M.; de Kloet, E.R. Point mutation in the mouse glucocorticoid receptor preventing DNA binding impairs spatial memory. Proc. Natl. Acad. Sci. 2001, 98, 12790–12795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, E.; Moser, M.-B.; Andersen, P. Spatial learning impairment parallels the magnitude of dorsal hippocampal lesions, but is hardly present following ventral lesions. J. Neurosci. 1993, 13, 3916–3925. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M.S.; Dong, H.-W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.A.; Beattie, J.; Reid, I.; Balfour, D. Regulation of corticosteroid receptors in the rat brain: The role of serotonin and stress. Eur. J. Neurosci. 2005, 21, 1511–1520. [Google Scholar] [CrossRef] [PubMed]
- Buret, L.; van den Buuse, M. Corticosterone treatment during adolescence induces down-regulation of reelin and NMDA receptor subunit GLUN2C expression only in male mice: Implications for schizophrenia. Int. J. Neuropsychopharmacol. 2014, 17, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Gass, P.; Reichardt, H.M.; Strekalova, T.; Henn, F.; Tronche, F. Mice with targeted mutations of glucocorticoid and mineralocorticoid receptors: Models for depression and anxiety? Physiol. Behav. 2001, 73, 811–825. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Stress Timing | Type of Stress | Behavioral Phenotype in Adulthood | Molecular Phenotype in Adulthood |
|---|---|---|---|
| Early life stress | psychological | depression-like behavior, cognitive impairment [41] | HPA-axis abnormalities [41] |
| memory deficits, enhanced emotional learning [42] | improved hippocampal neurogenesis [42] | ||
| improved hippocampal-dependent memory [43] | reduced levels of hippocampal glucocorticoid and mineralocorticoid receptors [43] | ||
| improved spatial learning and enhanced anxiety at 2 months, but spatial memory deficits and normal anxiety levels at 15 months [44] | impaired LTP under basal conditions, but increased LTP in response to high CORT [43] | ||
| depression-like behavior in males and females with greater effect in males [45] | decreased mBDNF in the dorsal hippocampus in males only [46] | ||
| increased preference for alcohol in males but not females [47] | increased mBDNF in the ventral hippocampus in females only [46] | ||
| Adolescent stress | physical and social | impaired learning behavior in radial water maze while working and spatial memory remained intact [48] | liposaccharide (LPS) induced exaggerated elevation of the pro-inflammatory cytokines IL-1β and TNF-α in males but not in females [49] |
| physical | spatial memory deficits and hippocampal volume changes [50] | reduced hippocampal GR, increased hippocampal volume [51] | |
| spatial memory deficits [51] | |||
| reduced social interaction, depression-like behavior [52] | |||
| spatial memory deficit [53] | |||
| social | no spatial memory deficits or hippocampal volume changes [50] | decreased amplitude of spontaneous excitatory postsynaptic currents in the PFC only in male mice; decreased amplitude of spontaneous excitatory postsynaptic currents in the nucleus accumbens only in female mice [54] | |
| impaired spatial memory in Y maze and working memory in Morris water maze, while social recognition memory and episodic memory are intact [55] | |||
| spatial memory deficits in males and females [56,57] | |||
| reduced contextual fear conditioning [58] | |||
| anxiety-like behavior [54] | |||
| pharmacological | no changes in spatial memory, novel object recognition, anhedonia or anxiety in males or females (CORT 50 mg/L) [46] | deficits in sensory gating as measured by PPI only in males (CORT 50 mg/L) [59] | |
| no spatial memory deficit (CORT 40 mg/L) | |||
| Adult stress | physical and social | depression-like/anxiety-like and submissive phenotype [60] | |
| physical | anxiety-like behavior [61] | reduced immune response [62] | |
| depression-like phenotype and body weight loss [63] | |||
| deficit in recall memory (Morris Water Maze) [64] | |||
| anhedonia-like phenotype [65] | |||
| anhedonia-like and anxiety-like phenotype [66] | |||
| anxiety-like phenotype in defensive burying test; no anxiety-like phenotype in EPM or light/dark box [67] | |||
| chronic restraint stress induced anxiety-like phenotype; CUS induces anxiety-like and depression-like phenotype [68] | |||
| somatic effects in males and anxiety-like phenotype in females [69] | |||
| 21 days of restraint stress enhanced spatial memory, while 28 days of restraint stress either had no effect or impaired spatial memory in females [70] | |||
| 14 days of stress enhanced spatial memory, while 21 days of stress impaired spatial memory in males [71] | |||
| social | depression- and anxiety-like phenotype; impaired memory [72] anxiety-like phenotype and episodic memory deficit [73] spatial-memory impairment [74] | increased oxidative stress and inflammation [72] reduced complexity of apical dendrites of CA3 neurons [74] | |
| Disruptions to normal circadian rhythm (physical and psychological) | depression-like and anxiety-like phenotype [75] | reduced dendritic length in the DG and CA1 [76] | |
| no changes in memory or anxiety [77] | |||
| hippocampal memory deficit [78] | |||
| anhedonia-like and depression-like phenotype; learning and memory impairment [76] | |||
| pharmacological | depression-like phenotype [79] | reduced hippocampal neurogenesis [80,81] | |
| increased anxiety-like behavior [82] | increased apoptosis [33,83,84] | ||
| depression-like phenotype in C57BL6/J but not in C57BL6/N [85] | reduced hippocampal neurogenesis in males and females [31] | ||
| enhanced emotionality score or no effect in females [86] | no changes in cell proliferation, survival or neuronal maturation in DG of the hippocampus in females [86] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeder, A.; Notaras, M.; Du, X.; Hill, R.A. On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior. Brain Sci. 2018, 8, 121. https://doi.org/10.3390/brainsci8070121
Schroeder A, Notaras M, Du X, Hill RA. On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior. Brain Sciences. 2018; 8(7):121. https://doi.org/10.3390/brainsci8070121
Chicago/Turabian StyleSchroeder, Anna, Michael Notaras, Xin Du, and Rachel A. Hill. 2018. "On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior" Brain Sciences 8, no. 7: 121. https://doi.org/10.3390/brainsci8070121
APA StyleSchroeder, A., Notaras, M., Du, X., & Hill, R. A. (2018). On the Developmental Timing of Stress: Delineating Sex-Specific Effects of Stress across Development on Adult Behavior. Brain Sciences, 8(7), 121. https://doi.org/10.3390/brainsci8070121