
Parenteral Antioxidant Supplementation at Birth Improves the Response to Intranasal Vaccination in Newborn Dairy Calves



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Treatment Allocation, and Sample Collection
2.2. Assessment of Transfer of Passive Immunity
2.3. Systemic Redox Balance Determination
2.4. Quantification of Mucosal Antigen-Specific Immunoglobulin A
2.5. Statistical Analyses
3. Results and Discussion
3.1. Homogeneity of Groups
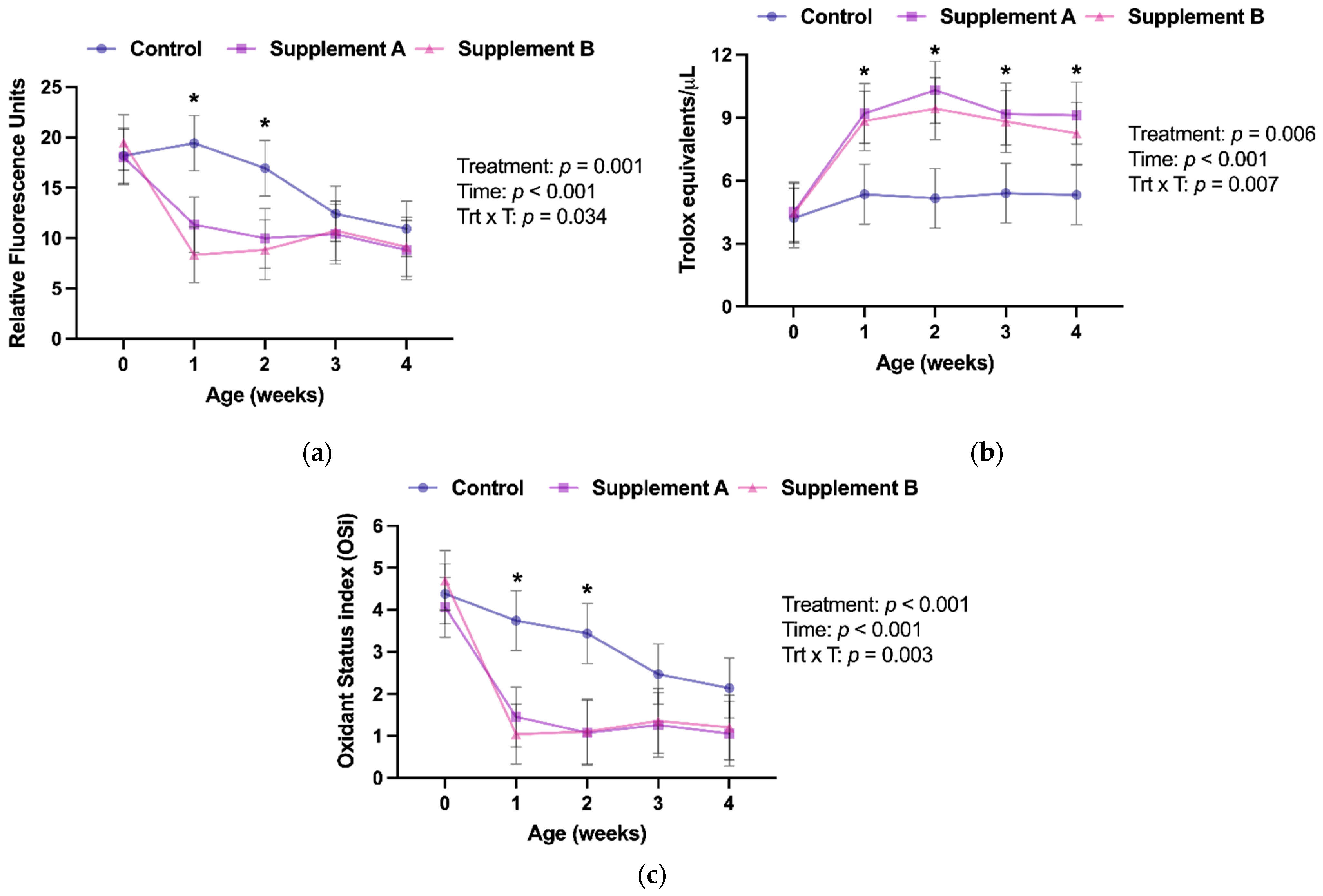
3.2. Redox Balance
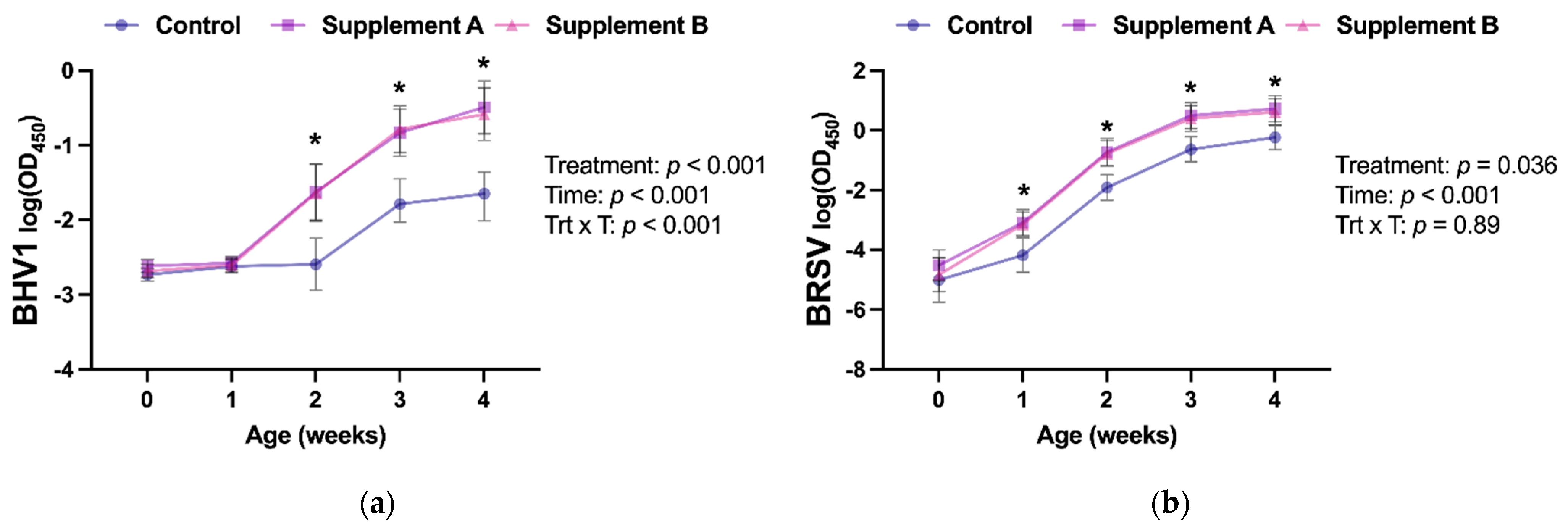
3.3. Intranasal Vaccine Response
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NAHMS: National Animal Health Monitoring Service. Dairy 2014. Part 1: Reference of Dairy Health and Management in the United States; USDA, APHIS Veterinary Services: Fort Collins, CO, USA, 2014.
- Windeyer, M.C.; Leslie, K.E.; Godden, S.M.; Hodgins, D.C.; Lissemore, K.D.; LeBlanc, S.J. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med. 2014, 113, 231–240. [Google Scholar] [CrossRef]
- Abuelo, A.; Havrlant, P.; Wood, N.; Hernandez-Jover, M. An investigation of dairy calf management practices, colostrum quality, failure of transfer of passive immunity, and occurrence of enteropathogens among Australian dairy farms. J. Dairy Sci. 2019, 102, 8352–8366. [Google Scholar] [CrossRef]
- NAHMS: National Animal Health Monitoring Service. Dairy Heifer Morbidity, Mortality and Health Management Practices Focusing on Preweaned Heifers; USDA, APHIS Veterinary Services: Ft Collins, CO, USA, 2007.
- Chase, C.C.; Hurley, D.J.; Reber, A.J. Neonatal immune development in the calf and its impact on vaccine response. Vet. Clin. N. Am. Food Anim. Pr. 2008, 24, 87–104. [Google Scholar] [CrossRef]
- Windeyer, M.C.; Gamsjager, L. Vaccinating Calves in the Face of Maternal Antibodies: Challenges and Opportunities. Vet. Clin. N. Am. Food Anim. Pr. 2019, 35, 557–573. [Google Scholar] [CrossRef] [PubMed]
- Chase, C.; Kaushik, R.S. Mucosal Immune System of Cattle: All Immune Responses Begin Here. Vet. Clin. N. Am. Food Anim. Pr. 2019, 35, 431–451. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, E.N. Exploiting mucosal surfaces for the development of mucosal vaccines. Vaccine 2011, 29, 8506–8511. [Google Scholar] [CrossRef]
- Abuelo, A.; Perez-Santos, M.; Hernandez, J.; Castillo, C. Effect of colostrum redox balance on the oxidative status of calves during the first 3 months of life and the relationship with passive immune acquisition. Vet. J. 2014, 199, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranade, R.; Talukder, S.; Muscatello, G.; Celi, P. Assessment of oxidative stress biomarkers in exhaled breath condensate and blood of dairy heifer calves from birth to weaning. Vet. J. 2014, 202, 583–587. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. Redox Biology in Transition Periods of Dairy Cattle: Role in the Health of Periparturient and Neonatal Animals. Antioxidants 2019, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Cuervo, W.; Sordillo, L.M.; Abuelo, A. Oxidative Stress Compromises Lymphocyte Function in Neonatal Dairy Calves. Antioxidants 2021, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Woolums, A.R.; Berghaus, R.D.; Berghaus, L.J.; Ellis, R.W.; Pence, M.E.; Saliki, J.T.; Hurley, K.A.; Galland, K.L.; Burdett, W.W.; Nordstrom, S.T.; et al. Effect of calf age and administration route of initial multivalent modified-live virus vaccine on humoral and cell-mediated immune responses following subsequent administration of a booster vaccination at weaning in beef calves. Am. J. Vet. Res. 2013, 74, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Froehlich, K.A.; Abdelsalam, K.W.; Chase, C.; Koppien-Fox, J.; Casper, D.P. Evaluation of essential oils and prebiotics for newborn dairy calves. J. Anim. Sci. 2017, 95, 3772–3782. [Google Scholar] [CrossRef] [PubMed]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Animal 2013, 7 (Suppl. S1), 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buczinski, S.; Gicquel, E.; Fecteau, G.; Takwoingi, Y.; Chigerwe, M.; Vandeweerd, J.M. Systematic Review and Meta-Analysis of Diagnostic Accuracy of Serum Refractometry and Brix Refractometry for the Diagnosis of Inadequate Transfer of Passive Immunity in Calves. J. Vet. Intern. Med. 2018, 32, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Godden, S.M.; Lombard, J.E.; Woolums, A.R. Colostrum Management for Dairy Calves. Vet. Clin. N. Am. Food Anim. Pr. 2019, 35, 535–556. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Gandy, J.C.; Neuder, L.; Brester, J.; Sordillo, L.M. Short communication: Markers of oxidant status and inflammation relative to the development of claw lesions associated with lameness in early lactation cows. J. Dairy Sci. 2016, 99, 5640–5648. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. Oxidative stress index (OSi) as a new tool to assess redox status in dairy cattle during the transition period. Animal 2013, 7, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Lombard, J.; Urie, N.; Garry, F.; Godden, S.; Quigley, J.; Earleywine, T.; McGuirk, S.; Moore, D.; Branan, M.; Chamorro, M.; et al. Consensus recommendations on calf- and herd-level passive immunity in dairy calves in the United States. J. Dairy Sci. 2020, 103, 7611–7624. [Google Scholar] [CrossRef]
- Ling, T.; Hernandez-Jover, M.; Sordillo, L.M.; Abuelo, A. Maternal late-gestation metabolic stress is associated with changes in immune and metabolic responses of dairy calves. J. Dairy Sci. 2018, 101, 6568–6580. [Google Scholar] [CrossRef] [PubMed]
- Perez-Santos, M.; Castillo, C.; Hernandez, J.; Abuelo, A. Biochemical variables from Holstein-Friesian calves older than one week are comparable to those obtained from adult animals of stable metabolic status on the same farm. Vet. Clin. Pathol. 2015, 44, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albera, E.; Kankofer, M. The comparison of antioxidative/oxidative profile in blood, colostrum and milk of early post-partum cows and their newborns. Reprod. Domest. Anim. 2011, 46, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Celi, P. Biomarkers of oxidative stress in ruminant medicine. Immunopharmacol. Immunotoxicol. 2011, 33, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef]
- Weiss, W.P.; Colenbrander, V.F.; Cunningham, M.D.; Callahan, C.J. Selenium/Vitamin E: Role in Disease Prevention and Weight Gain of Neonatal Calves. J. Dairy Sci. 1983, 66, 1101–1107. [Google Scholar] [CrossRef]
- Teixeira, A.G.; Lima, F.S.; Bicalho, M.L.; Kussler, A.; Lima, S.F.; Felippe, M.J.; Bicalho, R.C. Effect of an injectable trace mineral supplement containing selenium, copper, zinc, and manganese on immunity, health, and growth of dairy calves. J. Dairy Sci. 2014, 97, 4216–4226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, B.; Sblendorio, V. Oxidative stress tests: Overview on reliability and use. Part I. Eur. Rev. Med. Pharmacol. Sci. 2007, 11, 309–342. [Google Scholar] [PubMed]
- Palmieri, B.; Sblendorio, V. Oxidative stress tests: Overview on reliability and use. Part II. Eur. Rev. Med. Pharmacol. Sci. 2007, 11, 383–399. [Google Scholar]
- Hill, K.; Arsic, N.; Nordstrom, S.; Griebel, P.J. Immune memory induced by intranasal vaccination with a modified-live viral vaccine delivered to colostrum fed neonatal calves. Vaccine 2019, 37, 7455–7462. [Google Scholar] [CrossRef]
- Ellis, J.A.; Gow, S.P.; Mahan, S.; Leyh, R. Duration of immunity to experimental infection with bovine respiratory syncytial virus following intranasal vaccination of young passively immune calves. J. Am. Vet. Med. Assoc. 2013, 243, 1602–1608. [Google Scholar] [CrossRef] [Green Version]
- Bornheim, H.N.; Chamorro, M.F.; Cernicchiaro, N.; Reppert, E.J.; Larson, R.L.; Huser, S.; Thoresen, M.; Jones, K.; Weaber, R.L.; Woolums, A.R. Evaluation of specific immunoglobulin A in nasal secretions and neutralizing antibodies in serum collected at multiple time points from young beef calves following intranasal or subcutaneous administration of a modified-live bovine respiratory syncytial virus vaccine. Am. J. Vet. Res. 2021, 82, 746–751. [Google Scholar] [CrossRef] [PubMed]
- Palomares, R.A.; Hurley, D.J.; Bittar, J.H.; Saliki, J.T.; Woolums, A.R.; Moliere, F.; Havenga, L.J.; Norton, N.A.; Clifton, S.J.; Sigmund, A.B.; et al. Effects of injectable trace minerals on humoral and cell-mediated immune responses to Bovine viral diarrhea virus, Bovine herpes virus 1 and Bovine respiratory syncytial virus following administration of a modified-live virus vaccine in dairy calves. Vet. Immunol. Immunopathol. 2016, 178, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Heaton, P.R.; Reed, C.F.; Mann, S.J.; Ransley, R.; Stevenson, J.; Charlton, C.J.; Smith, B.H.E.; Harper, E.J.; Rawlings, J.M. Role of Dietary Antioxidants to Protect against DNA Damage in Adult Dogs. J. Nutr. 2002, 132, 1720S–1724S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuelo, A.; Cullens, F.; Hanes, A.; Brester, J.L. Impact of 2 Versus 1 Colostrum Meals on Failure of Transfer of Passive Immunity, Pre-Weaning Morbidity and Mortality, and Performance of Dairy Calves in a Large Dairy Herd. Animals 2021, 11, 782. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | Control | Supplement A | Supplement B | p-Value |
|---|---|---|---|---|
| Sex (n female/n male) | 6 a/1 | 6 a/1 | 6/1 | 1.0 |
| Birth weight (kg) | 37.2 (29.0–51.7) | 36.7 (29.9–43.5) | 37.2 (32.2–47.2) | 0.98 |
| Serum total protein (g/dL) | 6.5 (6.3–6.8) | 6.6 (6.2–6.9) | 6.5 (6.2–6.8) | 0.93 |
| Variable (Units) | Treatment Groups | p-Values | ||||
|---|---|---|---|---|---|---|
| Control | Suppl. A | Suppl. B | Control vs. Suppl. A | Control vs. Suppl. B | Suppl. A vs. Suppl. B | |
| Reactive Oxygen and Nitrogen Species (RFU) | 15.6 (14.5–16.7) | 11.7 (10.6–12.9) | 11.3 (10.2–12.5) | 0.0003 | 0.0001 | 0.85 |
| Antioxidant Potential (TE/μL) | 5.09 (3.85–6.33) | 8.07 (6.82–9.32) | 7.46 (6.21–8.71) | 0.0062 | 0.0288 | 0.76 |
| Oxidant Status index (arbitrary units) | 3.23 (2.82–3.65) | 1.78 (1.35–2.22) | 1.88 (1.45–2.32) | 0.0002 | 0.0005 | 0.94 |
| Anti-BHV1 IgA (log (OD450)) | −2.27 (−2.39–−2.15) | −1.63 (−1.76–−1.50) | −1.65 (−1.79–−1.53) | <0.0001 | <0.0001 | 0.93 |
| Anti-BRSV IgA (log (OD450)) | −2.43 (−3.10–−1.76) | −1.12 (−1.77–−0.67) | −1.44 (−2.09–−0.79) | 0.036 | 0.024 | 0.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nayak, A.; Abuelo, A. Parenteral Antioxidant Supplementation at Birth Improves the Response to Intranasal Vaccination in Newborn Dairy Calves. Antioxidants 2021, 10, 1979. https://doi.org/10.3390/antiox10121979
Nayak A, Abuelo A. Parenteral Antioxidant Supplementation at Birth Improves the Response to Intranasal Vaccination in Newborn Dairy Calves. Antioxidants. 2021; 10(12):1979. https://doi.org/10.3390/antiox10121979
Chicago/Turabian StyleNayak, Arpita, and Angel Abuelo. 2021. "Parenteral Antioxidant Supplementation at Birth Improves the Response to Intranasal Vaccination in Newborn Dairy Calves" Antioxidants 10, no. 12: 1979. https://doi.org/10.3390/antiox10121979
APA StyleNayak, A., & Abuelo, A. (2021). Parenteral Antioxidant Supplementation at Birth Improves the Response to Intranasal Vaccination in Newborn Dairy Calves. Antioxidants, 10(12), 1979. https://doi.org/10.3390/antiox10121979