
Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Preparation and In Vitro Condition
2.2. Determination of Heavy Metal Accumulation and Nitric Oxide Concentrations in Shoots, Stems, and Roots
2.3. Measurement of Concentrations of Protein Thiol, Nonprotein, and a Total of Thiol
2.4. Quantification of Glycine Betaine (GB), Proline Contents (Pro), and GSH
2.5. Determination of Lipid Peroxidation (MDA), Superoxide Radical (O2•−), Hydrogen Peroxide (H2O2), Soluble Proteins (SP), and Electrolyte Leakage (EL)
2.6. Determination of Antioxidant Activities
2.7. Determination of Photosynthetic Pigments (Chlorophyll and Carotenoids)
2.8. Determination of Shoot Dry Weight, Root Dry Weight, and Shoot Length
2.9. Statistical Analysis
3. Results
3.1. NO Donors (SNPs) Reduce Pb and Cd Accumulation in Shoot, Stems, and Roots in Plants under Heavy Metals
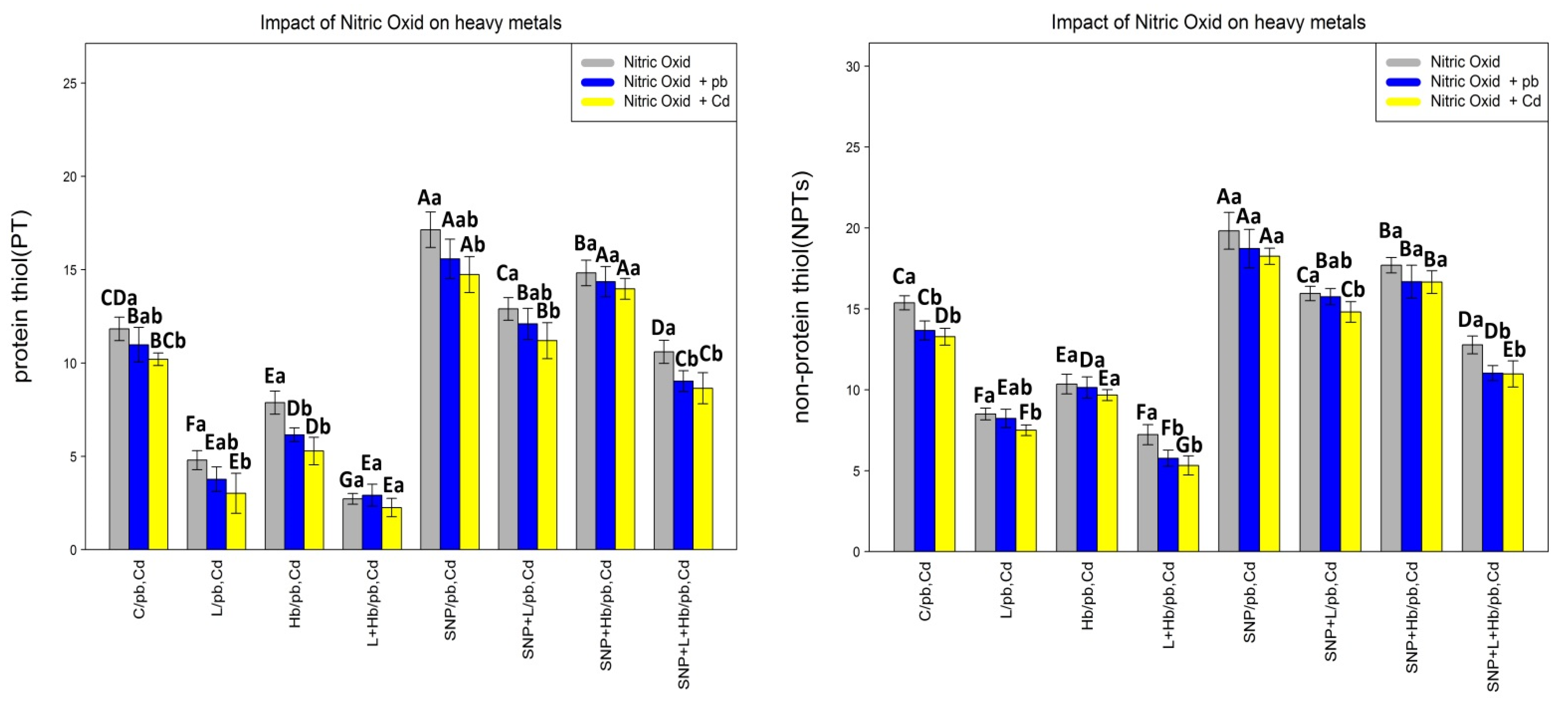
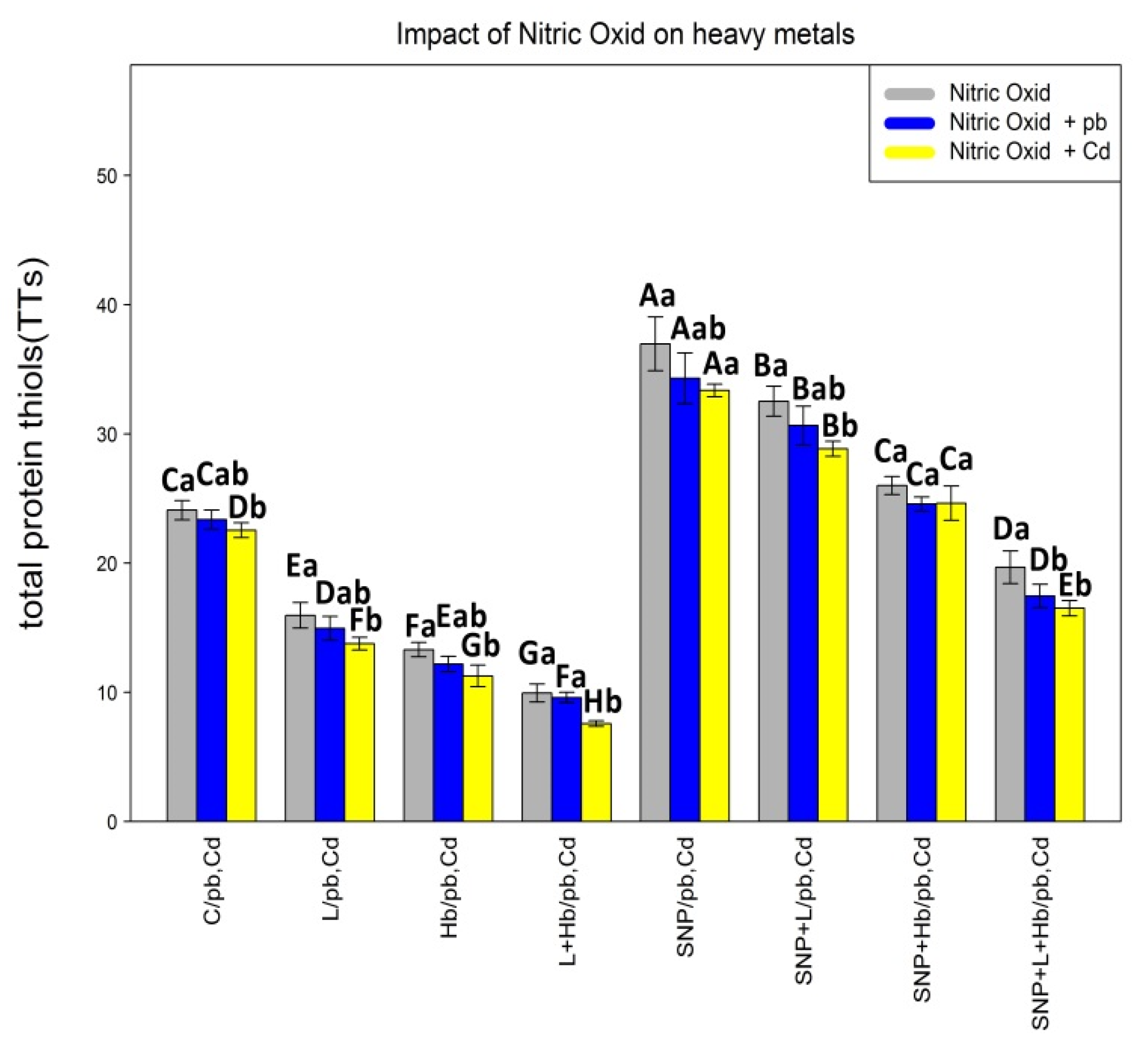
3.2. The NO Donor (SNP) Increased the Protein, Non-Protein, and Total Thiol Contents in Plants under Heavy Metal Toxicity (Pb and Cd)
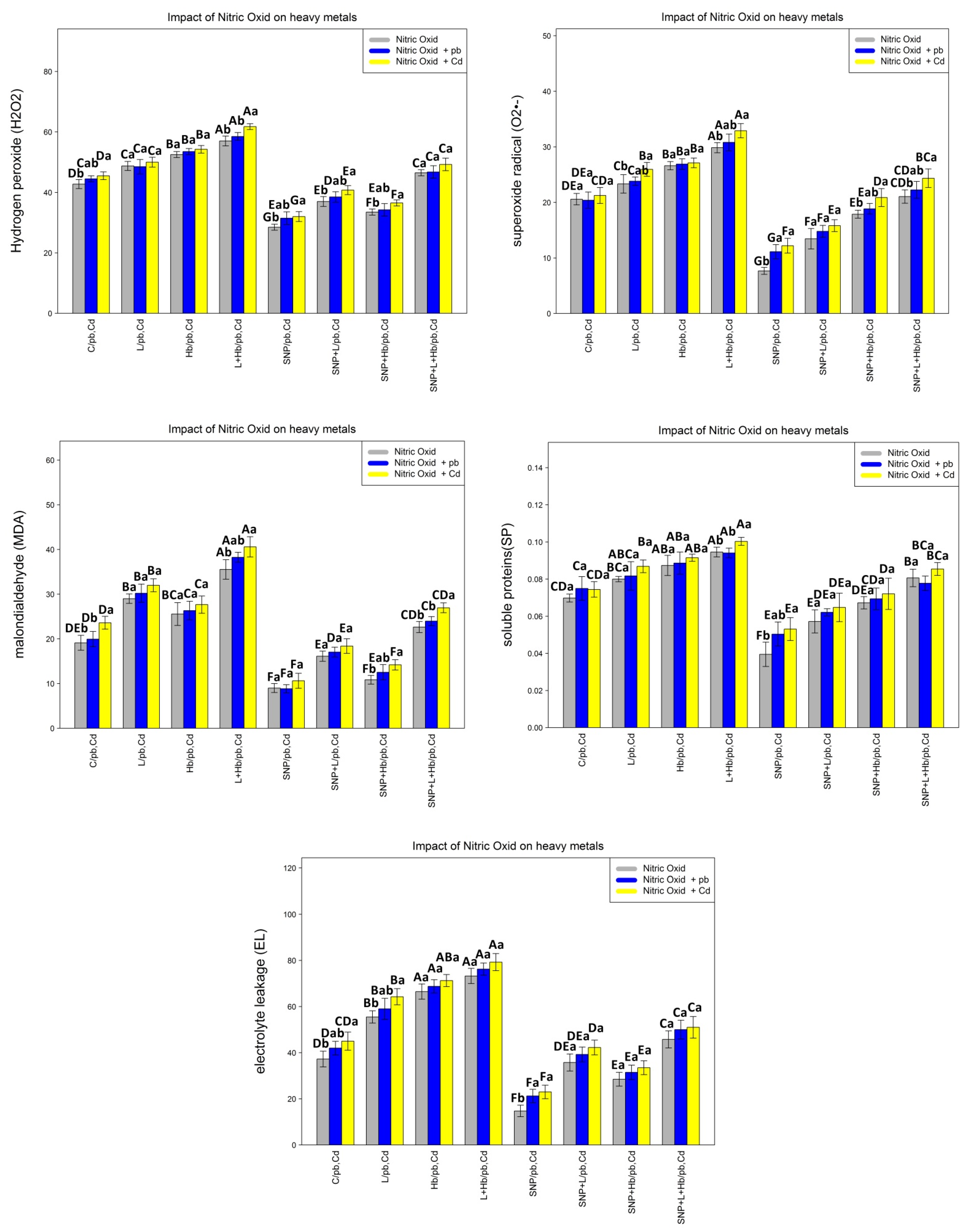
3.3. NO Donors (SNPs) Improve Membrane Injury and Increase the Cell Protection of Bamboo Plants under Heavy Metal Toxicity (Pb and Cd)
3.4. The NO Donor (SNP) Increased Glycine Betaine and Proline Contents and GSH Content in Plants Experiencing Heavy Metal Toxicity (Pb and Cd)
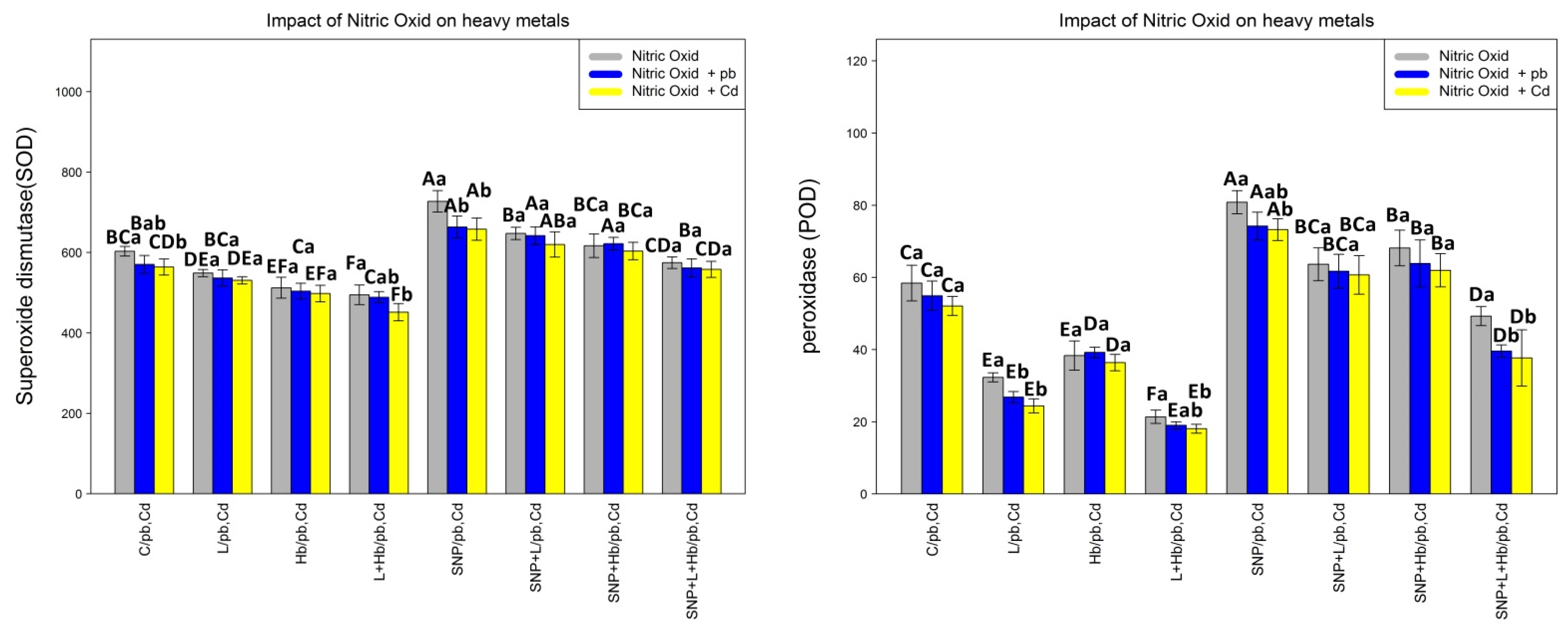
3.5. NO Donors (SNPs) Improve Antioxidant Enzyme Activities in Bamboo Plants under Heavy Metals (Pb and Cd)
3.6. NO Donors (SNPs) Improve Photosynthetic Pigments (Chlorophyll and Carotenoids) in Plants under Heavy Metal Toxicity (Pb and Cd)
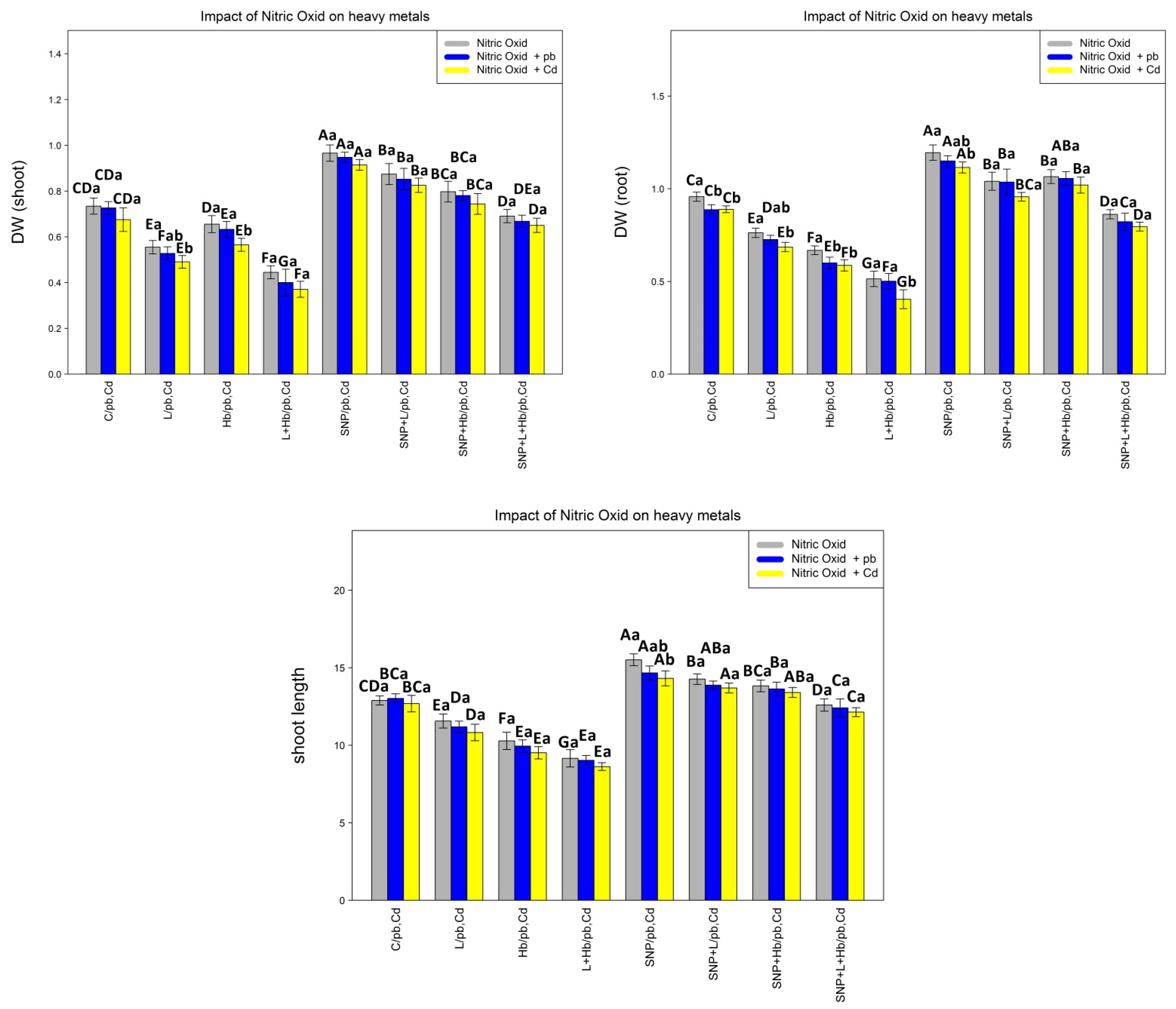
3.7. NO Donors (SNPs) Improve Plant Growth and Biomass under Pb and Cd Toxicity
3.8. NO Donors (SNPs) Reduce the Translocation Factor (TF) and Bioaccumulation Factor in Shoots, as Well as Increase the Tolerance Index (TI) in the Shoots and Roots of Bamboo Species under 200 μM Pb and 200 μM Cd
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wuana, R.A.; Okieimen, F.E. Heavy metals in contaminated soils: A review of sources, chemistry, risks and best available strategies for remediation. Int. Sch. Res. Not. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhong, T.; Liu, L.; Ouyang, X. Impact of soil heavy metal pollution on food safety in China. PLoS ONE 2015, 10, e0135182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; De Sherbinin, A.; Ye, C.; Shi, G. China’s soil pollution: Farms on the frontline. Science 2014, 344, 691. [Google Scholar] [CrossRef]
- Rian, Y.; He, L.; Cai, R.; Li, B.; Li, Z.; Yang, K. Heavy metal pollution and health risk in China. Glob. Health J. 2017, 1, 47–55. [Google Scholar] [CrossRef]
- Qin, Q.; Li, X.; Wu, H.; Zhang, Y.; Feng, Q.; Tai, P. Characterization of cadmium (108Cd) distribution and accumulation in Tageteserecta L. seedlings: Effect of split-root and of remove-xylem/phloem. Chemosphere 2013, 93, 2284–2288. [Google Scholar] [CrossRef]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J.; Gleń-Karolczyk, K. The use of macroelements from municipal sewage sludge by the Multiflora rose and the Virginia fanpetals. J. Ecol. Eng. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Serrano, R.M.; Puertas, R.M.C.; Zabalza, A.; Corpas, F.J.; Gomez, M.; Del Rio, L.A.; Sandalio, L.M. Cadmium effect on oxidativemetabolism of pea (Pisum sativum L.) roots. Imaging of reactive oxygen species and nitric oxide accumulationin vivo. Plant Cell Environ. 2006, 29, 1532–1544. [Google Scholar] [CrossRef]
- Nowak, H.B.; Dresler, S.; Matraszek, R. Exogenous malic and acetic acids reduce cadmium phytotoxicity and enhance cadmiumaccumulation in roots of sunflower plants. Plant Physiol. Biochem. 2015, 94, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Stegeman, J.J.; Brouwer, M.; Di Giulio, R.T.; Forlin, L.; Fowler, B.A. Enzyme and protein synthesis as indicator of contaminantexposure and effect. In Biomarkers: Biochemical, Physiological, and Histological Markers of Anthropogenic Stress; Huggett, R.J., Ed.; SETAC Special Publication Series; Lewis Publishers: Chelsea, MI, USA, 1992; pp. 235–335. [Google Scholar]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Antioxidant response of bamboo (Indocalamus latifolius) as affected by heavy metal stress. J. Elem. 2018, 23, 341–352. [Google Scholar] [CrossRef]
- Malar, S.; Manikandan, R.; Favas, P.J.; Vikram, S.S.; Venkataahalam, P. Effect of lead on phytotoxicity, growth, biochemical alterations and its role on genomic template stability in Sesbania grandiflora: A potential plant for phytoremediation. Ecotoxicol. Environ. Saf. 2014, 108, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Domingos, P.; Prado, A.M.; Wong, A.; Gehring, C.; Feijo, J.A. Nitric oxide: A multitasked signaling gas in plants. Mol. Plant 2015, 8, 506–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, M.; Lee, G.I.; Wang, Y. AtNOS/AtNOA1 is a functional Arabidopsis thaliana cGTPase and not a nitric oxide synthase. J. Biol. Chem. 2008, 283, 32957–32967. [Google Scholar] [CrossRef] [Green Version]
- Buet, A.; Galatro, A.; Ramos-Artuso, F.; Simontacchi, M. Nitric oxide and plant mineral nutrition: Current knowledge. J. Exp. Bot. 2019, 70, 4461–4476. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.G.; Tian, Q.Y.; Zhang, W.H. Nitric oxide synthase-dependent nitric oxide production is associated with salt tolerance in Arabidopsis. Plant Physiol. 2007, 144, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Tamás, L.; Demecsová, L.; Zelinová, V. L-NAME decreases the amount of nitric oxide and enhances the toxicity of cadmium via superoxide generation in barley root tip. J. Plant Physiol. 2018, 224, 68–74. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N.; Farooq, M.A.; Sandalio, L.M. Nitric oxide improves tolerance to arsenic stress in Isatis cappadocica desv. Shoots by enhancing antioxidant defenses. Chemosphere 2020, 239, 124523. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Alyemeni, M.; Alyemeni, M.; Wijaya, L.; Wijaya, L. Nitric oxide donor, sodium nitroprusside, mitigates mercury toxicity in different cultivars of soybean. J. Hazard. Mater. 2020, 408, 124852. [Google Scholar] [CrossRef]
- Rezayian, M.; Ebrahimzadeh, H.; Niknam, V. Nitric oxide stimulates antioxidant system and osmotic adjustment in soybean under drought stress. J. Soil Sci. Plant Nutr. 2020, 20, 1122–1132. [Google Scholar] [CrossRef]
- Akhter Bhat, J.; Ahmad, P.; Corpas, F.J. Main nitric oxide (NO) hallmarks to relieve arsenic stress in higher plants. J. Hazard. Mater. 2021, 406, 124289. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Liu, W.; Jing, Q.; Cao, W. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environ. Exp. Bot. 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense, and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Farnese, F.S.; Oliveira, J.A.; Paiva, E.A.; Menezes-Silva, P.E.; da Silva, A.A.; Campos, F.V.; Ribeiro, C. The involvement of nitric oxide in integration of plant physiological and ultrastructural adjustments in response to arsenic. Front. Plant Sci. 2017, 8, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratner, B.A.; Massod, A.; Sehar, Z.; Majid, A.; Anjum, N.A.; Khan, N.A. Mechanisms and role of nitric oxide in phytotoxicity-mitigation of copper. Front. Plant Sci. 2020, 11, 675. [Google Scholar] [CrossRef]
- Hogarth, N.; Belcher, B. The contribution of bamboo to household income and rural livelihoods in a poor and mountainouscounty in Guangxi, China. Int. For. Rev. 2013, 15, 71–81. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Zhang, Y.; Booth, T.; He, X. Changes of carbon stocks in bamboo stands in China during 100 years. For. Ecol. Manag. 2009, 258, 1489–1496. [Google Scholar] [CrossRef]
- Yao, W.; Li, C.; Lin, S.; Ren, L.; Wan, Y.; Zhang, L.; Ding, Y. Morphological characteristics and transcriptome comparisons of the shoot buds from flowering and non-flowering Pleioblastus pygmaeus. Forests 2020, 11, 1229. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Ranaei, F.; Ahmad, Z. Application of bamboo plants in nine aspects. Sci. World J. 2020, 2020, 7284203. [Google Scholar] [CrossRef]
- State Forestry Administration of China. Statistical Yearbook of Forestry; State Forestry Administration of China: Beijing, China, 2012.
- Ramakrishnan, M.; Yrjl, K.; Vinod, K.K.; Sharma, A.; Zhou, M. Genetics and genomics of moso bamboo (Phyllostachys edulis): Current status, future challenges, and biotechnological opportunities toward a sustainable bamboo industry. Food Energy Secur. 2020, 9, e229. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Ramakrishnan, M.; Ahmad, Z.; Xie, Y. Different Physiological and Biochemical Responses of Bamboo to the Addition of TiO2 NPs under Heavy Metal Toxicity. Forests 2021, 12, 759. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Karimi, N.; Ghaderian, S.M.; Raab, A.; Feldmann, J.; Meharg, A.A. An arsenic accumulating, hyper-tolerant brassica, Isatis cappadocica Desv. New Phytol. 2009, 184, 41–47. [Google Scholar] [CrossRef]
- Shukla, P.; Singh, A.K. Nitric oxide mitigates arsenic-induced oxidative stress and genotoxicity in Vicia faba L. Environ. Sci. Pollut. Res. Int. 2015, 22, 13881–13891. [Google Scholar] [CrossRef]
- Murphy, M.E.; Noack, E. Nitric oxide assay using hemoglobin method. Methods Enzymol. 1994, 233, 240–250. [Google Scholar]
- Sedlak, J.; Lindsay, R. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeonpea (Cajanus cajan (L.) Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Li, C.; Bai, T.; Ma, F.; Han, M. Hypoxia tolerance and adaptation of anaerobic respiration to hypoxia stress in two Malus species. Sci. Hortic. 2010, 124, 274–279. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid sensitive method for the quan-tification of microgram quantities of protein utilising the principle ofprotein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Valentovic, P.; Luxova, M.; Kolarovic, L.; Gasparikova, O. Effect of osmotic stress on compatible solutes content, membrane stability and water relations in two maize cultivars. Plant Soil Environ. 2006, 52, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Dhindsa, R.S.; Matowe, W. Drought tolerance in two mosses: Correlated with enzymatic defence against lipid peroxidation. J. Exp. Bot. 1981, 32, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. In Research Methodology of Crop Physiology; Agriculture Press: Beijing, China, 1992. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Foster, J.G.; Hess, J.L. Responses of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol. 1980, 66, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Rohman, M.M.; Hossain, M.D.; Suzuki, T.; Takada, G.; Fujita, M. Quercetin-4′-glucoside: A physiological inhibitor of the activities of dominant glutathione S-transferases in onion (Allium cepa L.) bulb. Acta Physiol. Plant. 2009, 31, 301–309. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV–VIS spectroscopy. In Current Protocols in Food Analytical Chemistry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2001; p. 4. [Google Scholar]
- Souri, Z.; Karimi, N. Enhanced phytoextraction by as hyperaccumulator Isatis cappadocica spiked with sodium nitroprusside. Soil Sediment Contam. Int. J. 2017, 26, 457–468. [Google Scholar] [CrossRef]
- Víteček, J.; Lojek, A.; Valacchi, G.; Kubala, L. Arginine-based inhibitors of nitric oxide synthase: Therapeutic potential and challenges. Mediat. Inflamm. 2012, 2012, 318087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Kabir, A.H.; Song, Y.; Lee, S.-H.; Hasanuzzaman, M.; Lee, K.-W. Nitric oxide prevents Fe deficiency-induced photosynthetic disturbance, and oxidative stress in alfalfa by regulating Fe acquisition and antioxidant defense. Antioxidants 2021, 10, 1556. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Inafuku, M.; Nahar, K.; Fujita, M.; Oku, H. Nitric oxide regulates plant growth, physiology, antioxidant defense, and Ion homeostasis to confer salt tolerance in the mangrove species, Kandelia obovata. Antioxidants 2021, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- Terrón-Camero, L.C.; Peláez-Vico, M.A.; Del Val, C.; Sandalio, L.M.; Romero-Puertas, M.C. Role of nitric oxide in plant responses to heavy metal stress: Exogenous application versusendogenous production. J. Exp. Bot. 2019, 70, 4477–4488. [Google Scholar] [CrossRef] [PubMed]
- Hill, B.G.; Dranka, B.P.; Bailey, S.M.; Lancaster, J.R.; Darley-Usmar, V.M. What part of NO don’t you understand? Some answers to the cardinal questions in nitric oxide biology. J. Biol. Chem. 2010, 285, 19699–19704. [Google Scholar] [CrossRef] [Green Version]
- Huaifu, F.; Shirong, G.; Yansheng, J.; Runhua, Z.; Juan, L. Effects of exogenous nitric oxide on growth, active oxygen species metabolism, and photosynthetic characteristics in cucumber seedlings under NaCl stress. Front. Agric. China 2007, 1, 308–314. [Google Scholar]
- Gan, L.; Wu, X.; Zhong, Y. Exogenously applied nitric oxide enhances the drought tolerance in hulless barley. Plant Prod. Sci. 2015, 18, 52–56. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.J.; Fernie, A.R.; Kaiser, W.M.; van Dongen, J.T. On the origins of nitric oxide. Trends Plant Sci. 2011, 16, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Sharma, V.P.; Sharma, N.; Kohli, R.K. Nitric oxide alleviates arsenic toxicity by reducing oxidative damage in the roots of Oryza sativa (rice). Nitric Oxide 2009, 20, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Fujita, M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology 2013, 22, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva, C.J.; Canatto, R.A.; Cardoso, A.A.; Ribeiro, C.; Oliveira, J.A. Oxidative stress triggered by arsenic in a tropical macrophyte is alleviated by endogenous and exogenous nitric oxide. Braz. J. Bot. 2017, 41, 21–28. [Google Scholar] [CrossRef]
- Baena, G.; Feria, A.B.; Echevarría, C.; Echevarría, C.; Monreal, J.A.; García-Mauriño, S. Salinity promotes opposite patterns of carbonylation and nitrosylation of C4 phosphoenolpyruvate carboxylase in sorghum leaves. Planta 2017, 246, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Chandrakar, V.; Yadu, B.; Meena, R.K.; Dubey, A.; Keshavkant, S. Arsenic-induced genotoxic responses and their amelioration by diphenylene iodonium, 24-epibrassinolide and proline in Glycine max L. Plant Physiol. Biochem. 2017, 112, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Mehta, S.K.; Liu, Z.P.; Yang, Z.M. Copper-induced proline synthesis is associated with nitric oxide generation in Chlamydomonas reinhardtii. Plant Cell Physiol. 2008, 49, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.K.; Liu, C.; Wang, F.; Ahammed, G.J.; Zhou, J.; Xu, M.X.; Yu, J.Q.; Xia, X.J. Glutathione-mediated regulation of nitric oxide, S-nitrosothiol and redox homeostasis confers cadmium tolerance by inducing transcription factors and stress response genes in tomato. Chemosphere 2016, 161, 36–45. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.I.; Neukermans, J.; Marquez−Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.M.; Diao, X.P.; ur Rehman, A.; Imtiaz, M.; Shakoor, A.; Altaf, M.A.; Younis, H.; Fu, P.; Ghani, M.U. Effect of vanadium on growth, photosynthesis, reactive oxygen species, antioxidant enzymes, and cell death of rice. J. Soil Sci. Plant Nutr. 2020, 20, 2643–2656. [Google Scholar] [CrossRef]
- Manai, J.; Kalai, T.; Gouia, H.; Corpas, F.J. Exogenous nitric oxide (NO) ameliorates salinity-induced oxidative stress in tomato (Solanum lycopersicum) plants. J. Soil Sci. Plant Nutr. 2014, 14, 433–446. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.S.P. Nitric oxide mediates hydrogenperoxide-and salicylic acid-induced salt tolerance rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Bai, X.Y.; Dong, Y.J.; Wang, Q.H.; Xu, L.L.; Kong, J.; Liu, S. Effects of lead and nitric oxide on photosynthesis, antioxidative ability, and mineral element content of perennial ryegrass. Biol. Plant 2015, 59, 163–170. [Google Scholar] [CrossRef]
- Egbichi, I.; Keyster, M.; Ludidi, N. Effect of exogenous application of nitric oxide on salt stress responses of soybean. S. Afr. J. Bot. 2014, 90, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Deckert, R.; Rucinska-Sobkowiak, R.; Gzyl, J.; Pawlak-Sprada, S.; Abramowski, D.; Jelonek, T.; Gwozdz, E.A. Nitric oxide implication in cadmium-induced programmed cell death in roots and signaling response of yellow lupine plants. Plant Physiol. Biochem. 2012, 58, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Usman, K.; Al-Ghouti, M.A.; Abu-Dieyeh, M.H. The assessment of cadmium, chromium, copper, and nickel tolerance and bioaccumulation by shrub plant Tetraena qataranse. Sci. Rep. 2019, 9, 5658. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.J.M.; Brooks, R.R.; Pease, A.J.; Malaisse, F. Studies on copper and cobalt tolerance in three closely related taxa within the genus Silene, L. (Caryophyllaceae) from Zaïre. Plant Soil 1983, 73, 377–385. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J. Phytoextraction of metals and metalloids from contaminated soils. Curr. Opin. Biotechnol. 2003, 14, 277–282. [Google Scholar] [CrossRef]
- Shukla, P.; Singh, S.; Dubey, P.; Singh, A.; Singh, A.K. Nitric oxide mediated amelioration of arsenic toxicity which alters the alternative oxidase (Aox1) gene expression in Hordeum vulgare L. Ecotoxol. Environ. Saf. 2015, 120, 59–65. [Google Scholar] [CrossRef]
- Yoon, J.; Cao, X.; Zhou, Q.; Ma, L.Q. Accumulation of Pb, Cu, and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef]
- Bian, F.; Zhong, Z.; Zhang, X.; Yang, C.; Gai, X. Bamboo—An untapped plant resource for the phytoremediation of heavy metal contaminated soils. Chemosphere 2020, 246, 125750. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.H.; Liu, Y.J.; Zhou, S.B.; Guo, F.G.; Bi, D.; Guo, X.H.; Baker, A.J.M.; Smith, J.A.C.; Luo, Y.M. Sedum plumbizincicola XH Guo et SB Zhou ex LH Wu (Crassulaceae): A new species from Zhejiang Province, China. Plant Syst. Evol. 2013, 299, 487–498. [Google Scholar] [CrossRef]
- Collin, B.; Doelsch, E.; Keller, C.; Panfili, F.; Meunier, J.D. Distribution and variability of silicon, copper and zinc in different bamboo species. Plant Soil 2010, 351, 377–387. [Google Scholar] [CrossRef]
- Bian, F.Y.; Zhong, Z.K.; Zhang, X.P.; Yang, C.B. Phytoremediation potential of moso bamboo (Phyllostachys pubescens) intercropped with Sedum plumbizincicola in metal-contaminated soil. Environ. Sci. Pollut. Res. 2017, 24, 27244–27253. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.Y.; Zhong, Z.K.; Wu, S.C.; Zhang, X.P.; Yang, C.B.; Xiong, X.Y. Comparison of heavy metal phytoremediation in monoculture and intercropping systems of Phyllostachys praecox and Sedum plumbizincicola in polluted soil. Int. J. Phytoremediat. 2018, 20, 490–498. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Concentrations |
|---|---|
| Control | 0 |
| Pb | 200 µM Pb |
| Cd | 200 µM Cd |
| L-NAME | 50 µM L-NAME |
| L-NAME + Pb | 50 µM L-NAME + 200 µM Pb |
| L-NAME + Cd | 50 µM L-NAME + 200 µM Cd |
| Hb | 0.1% Hb |
| Hb + Pb | 0.1% Hb + 200 µM Pb |
| Hb + Cd | 0.1% Hb + 200 µM Cd |
| L-NAME + Hb | 50 µM L-NAME + 0.1% Hb |
| L-NAME + Hb + Pb | 50 µM L-NAME + 0.1% Hb + 200 µM Pb |
| L-NAME + Hb + Cd | 50 µM L-NAME + 0.1% Hb + 200 µM Cd |
| SNP | 200 µM SNP |
| SNP + Pb | 200 µM SNP + 200 µM Pb |
| SNP + Cd | 200 µM SNP + 200 µM Cd |
| SNP + L-NAME | 200 µM SNP + 50 µM L-NAME |
| SNP + L-NAME + Pb | 200 µM SNP + 50 µM L-NAME + 200 µM Pb |
| SNP + L-NAME + Cd | 200 µM SNP + 50 µM L-NAME + 200 µM Cd |
| SNP + Hb | 200 µM SNP + 0.1% Hb |
| SNP + Hb + Pb | 200 µM SNP + 0.1% Hb + 200 µM Pb |
| SNP + Hb +Cd | 200 µM SNP + 0.1% Hb + 200 µM Cd |
| SNP + L-NAME + Hb | 200 µM SNP + 50 µM L-NAME + 0.1% Hb |
| SNP + L-NAME + Hb + Pb | 200 µM SNP + 50 µM L-NAME + 0.1% Hb + 200 µM Pb |
| SNP + L-NAME + Hb + Cd | 200 µM SNP + 50 µM L-NAME + 0.1% Hb + 200 µM Cd |
| Nitric Oxid Concentration | Heavy Metals Content | Heavy Metal Accumulation (Shoot) | Nitric Oxide Accumulation (Shoot) | Heavy Metal Accumulation (Stem) | Nitric Oxide Accumulation (Stem) | Heavy Metal Accumulation (Root) | Nitric Oxide Accumulation (Root) |
|---|---|---|---|---|---|---|---|
| µmol/L | µmol/L | µg/L | µg/L | µg/L | µg/L | µg/L | µg/L |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 0 | 200 µM Pb | 21.35 ± 0.61 Eb | 0 | 23.00 ± 0.81 Eb | 0 | 32.87 ± 0.47 Db | 0 |
| 0 | 200 µM Cd | 24.35 ± 0.38 Da | 0 | 25.25 ± 0.50 Ea | 0 | 35.90 ± 0.40 Da | 0 |
| 50 µM L-NAME | 0 | 0 | 15.55 ± 0.45 Da | 0 | 19.65 ± 0.90 Ea | 0 | 25.60 ± 0.35 Ea |
| 50 µM L-NAME | 200 µM Pb | 26.17 ± 1.11 Cb | 14.15 ± 0.50 Eb | 28.26 ± 0.93 Cb | 16.87 ± 0.53 Eb | 39.61 ± 1.00 Cb | 22.40 ± 0.82 Eb |
| 50 µM L-NAME | 200 µM Cd | 27.75 ± 0.52 Ca | 12.75 ± 0.50 Ec | 30.12 ± 0.83 Ca | 16.37 ± 0.81 Eb | 42.55 ± 0.43 Ca | 19.92 ± 1.09 Ec |
| 0.1% Hb | 0 | 0 | 12.75 ± 0.50 Ea | 0 | 14.87 ± 0.78 Fa | 0 | 17.75 ± 0.70 Fa |
| 0.1% Hb | 200 µM Pb | 29.25 ± 0.81 Bb | 11.52 ± 0.58 Fb | 32.47 ± 0.75 Bb | 11.92 ± 1.01 Fb | 45.37 ± 0.55 Bb | 15.12 ± 0.62 Fb |
| 0.1% Hb | 200 µM Cd | 31.25 ± 0.50 Ba | 10.75 ± 0.50 Fb | 35.25 ± 0.87 Ba | 10.25 ± 0.50 Fc | 47.81 ± 0.59 Ba | 14.07 ± 0.65 Fb |
| 50 µM L-NAME + 0.1% Hb | 0 | 0 | 8.67 ± 0.35 Fa | 0 | 10.00 ± 0.60 Ga | 0 | 11.97 ± 0.58 Ga |
| 50 µM L-NAME + 0.1% Hb | 200 µM Pb | 34.47 ± 0.51 Ab | 5.57 ± 0.41 Gb | 38.37 ± 0.93 Ab | 7.50 ± 0.29 Gb | 49.87 ± 0.59 Ab | 11.52 ± 0.48 Ga |
| 50 µM L-NAME + 0.1% Hb | 200 µM Cd | 36.02 ± 0.71 Aa | 4.57 ± 0.48 Gc | 39.87 ± 0.58 Aa | 6.45 ± 0.50 Gc | 53.12 ± 0.66 Aa | 8.27 ± 0.40 Gb |
| 200 µM SNP | 0 | 0 | 28.77 ± 0.55 Aa | 0 | 36.77 ± 0.41 Aa | 0 | 51.15 ± 0.90 Aa |
| 200 µM SNP | 200 µM Pb | 7.62 ± 0.38 Hb | 25.45 ± 0.58 Ab | 8.50 ± 0.60 Hb | 33.75 ± 0.50 Ab | 16.55 ± 0.50 Gb | 47.55 ± 0.63 Ab |
| 200 µM SNP | 200 µM Cd | 10.32 ± 0.46 Ga | 24.47 ± 0.23 Ac | 12.08 ± 0.61 Ha | 31.70 ± 0.53 Ac | 20.50 ± 1.14 Ga | 46.25 ± 1.03 Ab |
| SNP + L-NAME | 0 | 0 | 23.50 ± 0.53 Ba | 0 | 29.62 ± 0.45 Ba | 0 | 43.92 ± 0.80 Ba |
| SNP + L-NAME | 200 µM Pb | 12.25 ± 0.50 Gb | 21.42 ± 0.72 Bb | 13.92 ± 0.72 Gb | 27.22 ± 0.89 Bb | 23.57 ± 0.69 Fb | 41.12 ± 0.62 Bb |
| SNP + L-NAME | 200 µM Cd | 14.25 ± 0.52 Fa | 19.87 ± 0.45 Bc | 16.72 ± 0.71 Ga | 26.75 ± 0.50 Bb | 25.67 ± 0.75 Fa | 40.10 ± 0.54 Bb |
| SNP + Hb | 0 | 0 | 19.57 ± 0.55 Ca | 0 | 26.02 ± 0.73 Ca | 0 | 38.45 ± 0.38 Ca |
| SNP + Hb | 200 µM Pb | 17.27 ± 0.48 Fb | 18.05 ± 0.50 Cb | 18.60 ± 0.34 Fb | 23.60 ± 0.42 Cb | 28.55 ± 0.70 Eb | 35.45 ± 0.98 Cb |
| SNP + Hb | 200 µM Cd | 18.90 ± 0.98 Ea | 17.27 ± 0.85 Cb | 20.65 ± 1.48 Fa | 23.25 ± 0.50 Cb | 31.50 ± 0.66 Ea | 33.55 ± 0.59 Cc |
| SNP + L-NAME + Hb | 0 | 0 | 16.52 ± 0.58 Da | 0 | 22.02 ± 0.58 Da | 0 | 30.72 ± 0.65 Da |
| SNP + L-NAME + Hb | 200 µM Pb | 24.25 ± 0.50 Db | 15.75 ± 0.50 Da | 25.67 ± 0.83 Db | 20.15 ± 0.56 Db | 33.65 ± 0.77 Db | 27.10 ± 0.95 Db |
| SNP + L-NAME + Hb | 200 µM Cd | 26.37 ± 0.55 Ca | 15.47 ± 0.55 Ca | 27.83 ± 1.16 Da | 19.05 ± 0.50 Dc | 36.17 ± 0.79 Da | 26.67 ± 0.48 Db |
| Treament | Proline Content (Pro) | Glycine Betaine (GB) | Glutathione (GSH) |
|---|---|---|---|
| Control | 463.75 ± 19.73 CDa | 1015.06 ± 23.75 Ca | 148.75 ± 3.40 Ca |
| 200 µM Pb | 440.0 ± 29.43 BCa | 902.18 ± 35.98 Db | 141.25 ± 3.40 Cab |
| 200 µM Cd | 432.5 ± 28.72 BCa | 858.75 ± 39.23 Cb | 135.00 ± 4.89 Cb |
| 50 µM L-NAME | 367.50 ± 15.00 Ea | 712.50 ± 25.00 Ea | 115.75 ± 3.86 Ea |
| 50 µM L-NAME + 200 µM Pb | 340.0 ± 32.65 Dab | 650.00 ± 40.82 Fab | 107.50 ± 5.00 Eab |
| 50 µM L-NAME + 200 µM Cd | 312.5 ± 22.17 DEb | 620.00 ± 37.41 Db | 105.25 ± 6.23 Db |
| 0.1% Hb | 300.00 ± 24.49 Fa | 565.0 ± 40.41 Fa | 92.50 ± 5.06 Fa |
| 0.1% Hb + 200 µM Pb | 260.0 ± 21.60 Eab | 532.50 ± 41.93 Gab | 92.50 ± 5.06 Fa |
| 0.1% Hb + 200 µM Cd | 242.5 ± 22.17 EFb | 490.00 ± 27.08 Eb | 83.00 ± 10.23 Ea |
| 50 µM L-NAME + 0.1% Hb | 237.50 ± 25.00 Ga | 447.50 ± 29.86 Ga | 80.25 ± 1.89 Ga |
| 50 µM L-NAME + 0.1% Hb + 200 µM Pb | 202.5 ± 22.17 Ea | 427.50 ± 43.49 Ha | 75.25 ± 3.20 Ga |
| 50 µM L-NAME + 0.1% Hb + 200 µM Cd | 200.0 ± 40.82 Fa | 387.50 ± 29.86 Aa | 66.25 ± 2.50 Fb |
| 200 µM SNP | 620.00 ± 40.82 Aa | 1407.50 ± 61.84 Aa | 211.25 ± 2.62 Aa |
| 200 µM SNP + 200 µM Pb | 572.5 ± 28.72 Aa | 1320.00 ± 50.99 Aab | 188.50 ± 3.00 Ab |
| 200 µM SNP + 200 µM Cd | 562.5 ± 25.00 Aa | 1260.31 ± 66.22 Ab | 185.25 ± 5.43 Ab |
| SNP + L-NAME | 537.50 ± 17.07 Ba | 1180.0 ± 28.28 Ba | 177.50 ± 5.00 Ba |
| SNP + L-NAME + 200 µM Pb | 500.0 ± 14.14 Ba | 1148.75 ± 20.96 Ba | 170.75 ± 7.76 Ba |
| SNP + L-NAME+ 200 µM Cd | 500.0 ± 32.65 ABa | 1172.50 ± 43.49 Ba | 166.50 ± 5.68 Ba |
| SNP + Hb | 485.00 ± 34.39 BCa | 1037.50 ± 25.00 Ca | 157.50 ± 5.00 Ca |
| SNP + Hb + 200 µM Pb | 480.0 ± 41.63 Ba | 993.43 ± 29.11 Cab | 149.00 ± 6.05 Cab |
| SNP + Hb + 200 µM Cd | 457.5 ± 28.72 Ba | 952.50 ± 41.12 Bb | 143.25 ± 6.89 Cb |
| SNP + L-NAME + Hb | 410.00 ± 20.00 DEa | 826.25 ± 37.27 Da | 136.50 ± 5.06 Da |
| SNP + L-NAME + Hb + 200 µM Pb | 400.0 ± 32.65 CDa | 785.62 ± 29.14 Ea | 128.00 ± 6.97 Dab |
| SNP + L-NAME + Hb + 200 µM Cd | 380.0 ± 35.59 CDa | 712.81 ± 29.40 Cb | 119.25 ± 5.50 Db |
| Treatment | Chl-a | Chl-b | Chl a + b | Caratenoids |
|---|---|---|---|---|
| Control | 10.85 ± 0.74 BCa | 8.74 ± 0.58 Ca | 20.03 ± 0.76 Ca | 1.40 ± 0.31 BCa |
| 200 µM Pb | 10.61 ± 0.62 BCa | 8.59 ± 0.91 Ca | 19.28 ± 0.88 Ca | 1.28 ± 0.18 Ba |
| 200 µM Cd | 10.32 ± 0.51 BCa | 8.22 ± 0.59 CDa | 18.78 ± 1.27 Ca | 1.36 ± 0.22 ABa |
| 50 µM L-NAME | 9.20 ± 0.73 DEa | 6.92 ± 0.57 DEa | 16.51 ± 0.57 DEa | 1.04 ± 0.34 Ca |
| 50 µM L-NAME + 200 µM Pb | 9.00 ± 0.66 DEa | 7.00 ± 1.09 CDa | 15.98 ± 0.77 DEa | 1.14 ± 0.70 Ba |
| 50 µM L-NAME + 200 µM Cd | 8.58 ± 0.45 DEa | 6.77 ± 0.67 DEa | 15.55 ± 1.21 Da | 1.18 ± 0.39 Ba |
| 0.1% Hb | 7.91 ± 0.74 EFa | 7.57 ± 0.33 CDa | 15.41 ± 1.06 Ea | 1.70 ± 0.32 ABCa |
| 0.1% Hb + 200 µM Pb | 7.79 ± 0.49 EFa | 6.84 ± 0.55 DEab | 14.54 ± 0.18 Eab | 1.47 ± 0.09 Ba |
| 0.1% Hb + 200 µM Cd | 7.58 ± 0.80 EFa | 5.83 ± 1.00 EFb | 13.45 ± 0.29 Eb | 1.17 ± 0.60 Ba |
| 50 µM L-NAME + 0.1% Hb | 7.36 ± 0.56 Fa | 5.69 ± 0.51 Ea | 12.63 ± 1.23 Fa | 1.22 ± 0.37 Ca |
| 50 µM L-NAME + 0.1% Hb + 200 µM Pb | 7.09 ± 0.69 Fa | 5.30 ± 0.77 Ea | 12.07 ± 0.69 Fab | 1.15 ± 0.17 Ba |
| 50 µM L-NAME + 0.1% Hb + 200 µM Cd | 6.72 ± 0.65 Fa | 4.47 ± 0.75 Ea | 10.90 ± 0.44 Fb | 1.05 ± 0.40 Ba |
| 200 µM SNP | 12.89 ± 0.39 Aa | 12.81 ± 0.73 Aa | 25.73 ± 0.80 Aa | 2.24 ± 0.27 Aa |
| 200 µM SNP + 200 µM Pb | 12.27 ± 0.41 Aa | 12.55 ± 0.57 Aa | 24.78 ± 1.00 Aa | 2.29 ± 0.41 Aa |
| 200 µM SNP + 200 µM Cd | 12.18 ± 0.74 Aa | 12.16 ± 1.08 Aa | 24.28 ± 0.43 Aa | 2.13 ± 0.32 Aa |
| SNP + L-NAME | 11.93 ± 0.57 ABa | 11.96 ± 0.51 ABa | 23.51 ± 0.79 Ba | 2.20 ± 0.29 Aa |
| SNP + L-NAME + 200 µM Pb | 11.52 ± 0.38 ABa | 10.90 ± 0.52 Bab | 22.64 ± 0.97 Ba | 1.88 ± 0.30 ABa |
| SNP + L-NAME + 200 µM Cd | 11.46 ± 0.61 ABa | 11.22 ± 0.50 ABb | 22.32 ± 0.90 Ba | 2.06 ± 0.25 Aa |
| SNP + Hb | 11.10 ± 0.54 BCa | 10.52 ± 0.57 Ba | 21.42 ± 1.34 BCa | 1.94 ± 0.10 ABa |
| SNP + Hb + 200 µM pb | 11.01 ± 0.63 ABCa | 10.38 ± 0.33 Ba | 21.16 ± 1.09 BCa | 1.84 ± 0.07 ABa |
| SNP + Hb + 200 µM Cd | 10.69 ± 0.74 ABCa | 10.00 ± 1.03 BCa | 20.31 ± 0.70 Ca | 1.83 ± 0.30 ABa |
| SNP + L-NAME + Hb | 10.21 ± 0.89 CDa | 7.86 ± 1.13 CDa | 17.77 ± 0.63 Da | 1.22 ± 0.33 BC |
| SNP + L-NAME + Hb + 200 µM Pb | 9.84 ± 0.54 CDa | 7.46 ± 0.28 CDa | 17.08 ± 0.97 Da | 1.20 ± 0.14 B |
| SNP + L-NAME + Hb + 200 µM Cd | 9.65 ± 0.66 CDa | 7.05 ± 0.81 DEa | 16.43 ± 0.45 Da | 1.06 ± 0.22 B |
| Treatment | Translocation Factor (TF) | Tolerance Index (TI) (shoot) | Tolerance Index (TI) (root) | Bioaccumulation Factor (shoot) (BF) |
|---|---|---|---|---|
| Control | 0.00 ± 0.00 Cc | 1.00 ± 0.00 BCDa | 1.00 ± 0.00 CDa | 0.00 ± 0.00 Ac |
| 200 µM Pb | 0.64 ± 0.01 Ab | 0.95 ± 0.093 BCa | 0.95 ± 0.09 BCa | 0.10 ± 0.00 Db |
| 200 µM Cd | 0.67 ± 0.01 Aa | 0.90 ± 0.06 BCDa | 0.92 ± 0.01 BCa | 0.11 ± 0.00 Da |
| 50 µM L-NAME | 0.60 ± 0.01 ABb | 0.73 ± 0.07 EFa | 0.79 ± 0.01 EFa | 0.00 ± 0.00 Ac |
| 50 µM L-NAME + 200 µM Pb | 0.64 ± 0.01 Aab | 0.69 ± 0.08 DEa | 0.75 ± 0.01 DEb | 0.12 ± 0.00 Cb |
| 50 µM L-NAME + 200 µM Cd | 0.65 ± 0.03 Aa | 0.64 ± 0.08 EFa | 0.71 ± 0.01 DEc | 0.14 ± 0.00 Ca |
| 0.1% Hb | 0.71 ± 0.01 Aa | 0.87 ± 0.056 DEa | 0.69 ± 0.01 FGa | 0.00 ± 0.00 Ac |
| 0.1% Hb + 200 µM Pb | 0.66 ± 0.02 Ab | 0.83 ± 0.09 CDa | 0.65 ± 0.07 EFa | 0.14 ± 0.00 Bb |
| 0.1% Hb + 200 µM Cd | 0.67 ± 0.02 Aab | 0.77 ± 0.05 DEa | 0.60 ± 0.02 Ea | 0.15 ± 0.00 Ba |
| 50 µM L-NAME + 0.1% Hb | 0.69 ± 0.11 Aa | 0.58 ± 0.07 Fa | 0.56 ± 0.09 Ga | 0.00 ± 0.00 Ac |
| 50 µM L-NAME + 0.1% Hb + 200 µM Pb | 0.63 ± 0.05 Aa | 0.53 ± 0.11 Ea | 0.50 ± 0.04 Fa | 0.16 ± 0.00 Ab |
| 50 µM L-NAME + 0.1% Hb + 200 µM Cd | 0.65 ± 0.02 Aa | 0.48 ± 0.07 Fa | 0.44 ± 0.07 Fa | 0.17 ± 0.00 Aa |
| 200 µM SNP | 0.56 ± 0.004 ABa | 1.30 ± 0.093 Aa | 1.26 ± 0.04 Aa | 0.00 ± 0.00 Ac |
| 200 µM SNP + 200 µM Pb | 0.51 ± 0.01 Bb | 1.25 ± 0.11 Aa | 1.19 ± 0.009 Ab | 0.03 ± 0.00 Gb |
| 200 µM SNP + 200 µM Cd | 0.51 ± 0.02 Bb | 1.21 ± 0.11 Aa | 1.16 ± 0.02 Ab | 0.04 ± 0.00 Ga |
| SNP + L-NAME | 0.52 ± 0.03 Ba | 1.16 ± 0.082 ABa | 1.08 ± 0.05 BCa | 0.00 ± 0.00 Ac |
| SNP + L-NAME + 200 µM Pb | 0.51 ± 0.02 Ba | 1.14 ± 0.08 ABa | 1.06 ± 0.06 ABa | 0.06 ± 0.00 Fb |
| SNP + L-NAME + 200 µM Cd | 0.51 ± 0.003 Ba | 1.09 ± 0.10 ABa | 1.03 ± 0.09 ABa | 0.07 ± 0.00 Fa |
| SNP + Hb | 0.50 ± 0.02 Bb | 1.06 ± 0.06 BCa | 1.15 ± 0.12 ABa | 0.00 ± 0.00 Ac |
| SNP + Hb + 200 µM pb | 0.54 ± 0.03 Bab | 1.03 ± 0.10 ABCa | 1.12 ± 0.08 Aa | 0.08 ± 0.00 Eb |
| SNP + Hb + 200 µM Cd | 0.56 ± 0.02 Ba | 0.99 ± 0.054 BCa | 1.10 ± 0.11 Aa | 0.09 ± 0.00 Ea |
| SNP + L-NAME + Hb | 0.61 ± 0.14 ABa | 0.90 ± 0.10 CDEa | 0.90 ± 0.01 DEa | 0.00 ± 0.00 Ac |
| SNP + L-NAME + Hb + 200 µM Pb | 0.65 ± 0.01 Aa | 0.879 ± 0.09 CDa | 0.86 ± 0.05 CDab | 0.12 ± 0.00 Cb |
| SNP + L-NAME + H b+ 200 µM Cd | 0.65 ± 0.02 Aa | 0.85 ± 0.12 CDa | 0.83 ± 0.01 CDb | 0.12 ± 0.00 CDa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emamverdian, A.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Ramakrishnan, M.; Liu, G.; Li, Y. Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation. Antioxidants 2021, 10, 1981. https://doi.org/10.3390/antiox10121981
Emamverdian A, Ding Y, Barker J, Mokhberdoran F, Ramakrishnan M, Liu G, Li Y. Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation. Antioxidants. 2021; 10(12):1981. https://doi.org/10.3390/antiox10121981
Chicago/Turabian StyleEmamverdian, Abolghassem, Yulong Ding, James Barker, Farzad Mokhberdoran, Muthusamy Ramakrishnan, Guohua Liu, and Yang Li. 2021. "Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation" Antioxidants 10, no. 12: 1981. https://doi.org/10.3390/antiox10121981
APA StyleEmamverdian, A., Ding, Y., Barker, J., Mokhberdoran, F., Ramakrishnan, M., Liu, G., & Li, Y. (2021). Nitric Oxide Ameliorates Plant Metal Toxicity by Increasing Antioxidant Capacity and Reducing Pb and Cd Translocation. Antioxidants, 10(12), 1981. https://doi.org/10.3390/antiox10121981