
Lycopene Improves In Vitro Development of Porcine Embryos by Reducing Oxidative Stress and Apoptosis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Oocyte Collection and In Vitro Maturation (IVM)
2.3. In Vitro Fertilization (IVF) and In Vitro Culture (IVC)
2.4. Differential Staining of Blastocysts
2.5. Measurement of Intracellular ROS Levels
2.6. Measurement of Mitochondrial Membrane Potential (ΔΨm)
2.7. Immunocytochemical Staining
2.8. TUNEL Assay
2.9. Real-Time Quantitative PCR (qPCR)
2.10. Experimental Design
2.11. Statistical Analysis
3. Results
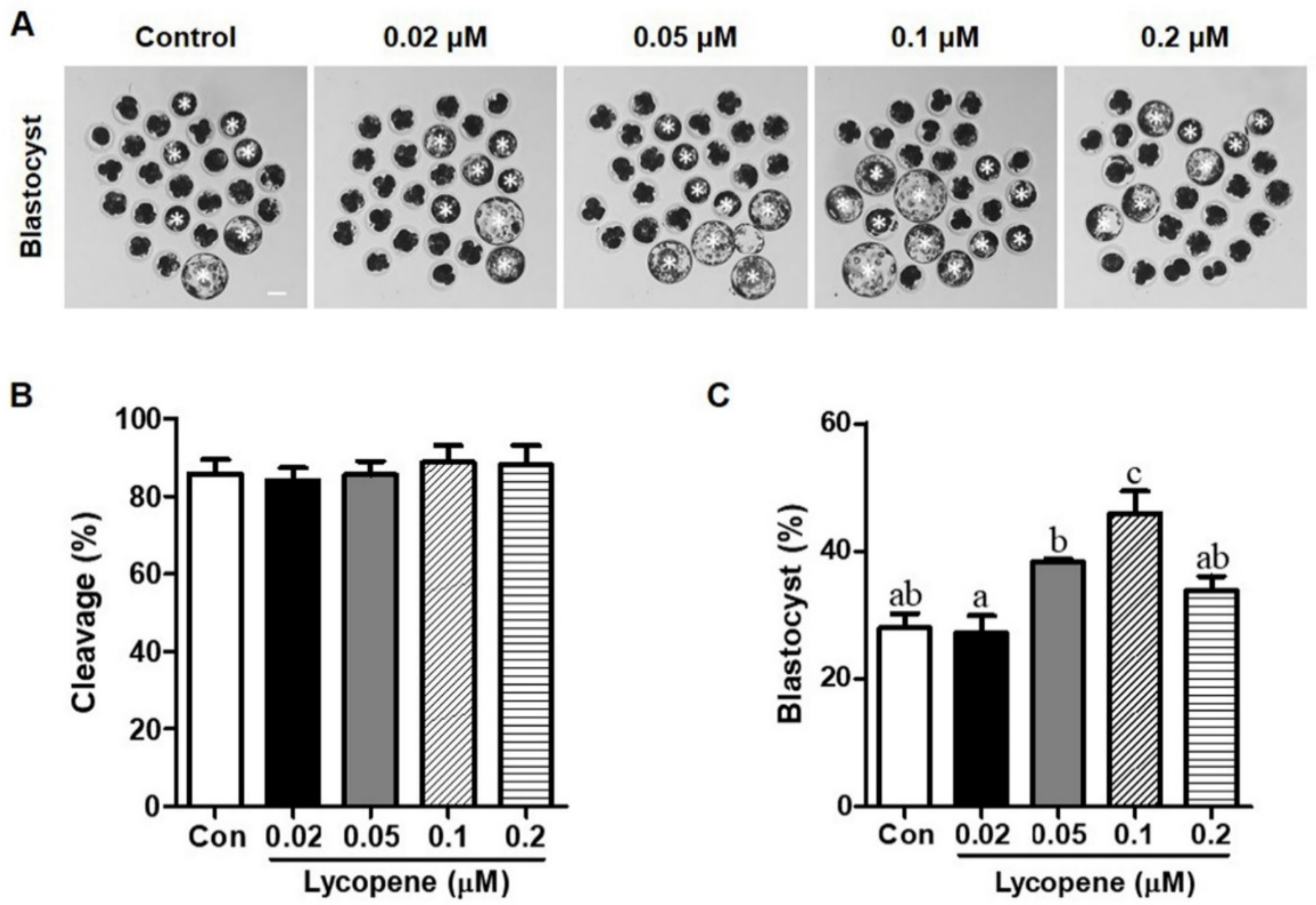
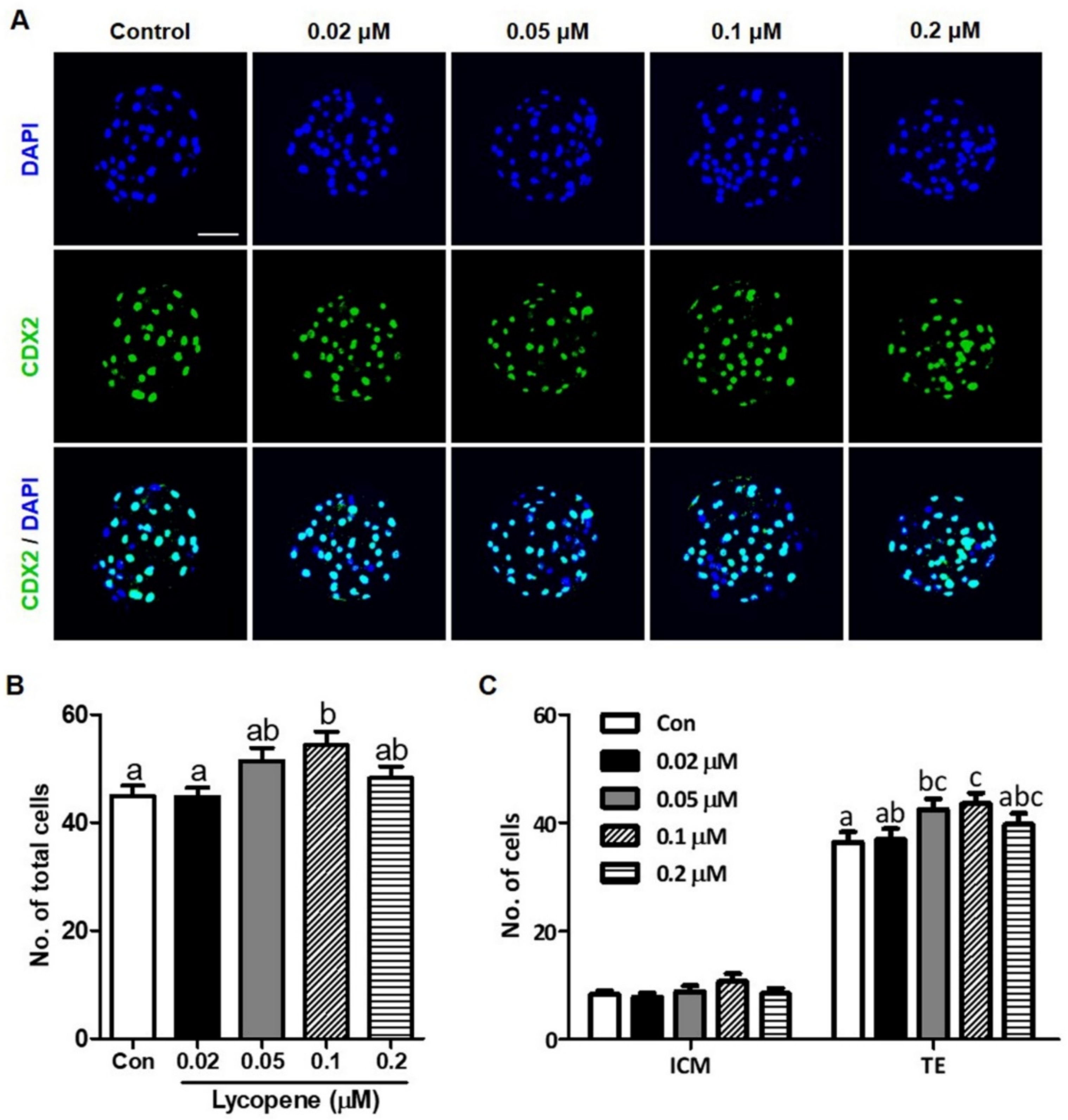
3.1. Effects of Various Concentrations of Lycopene on In Vitro Development of IVF Embryos
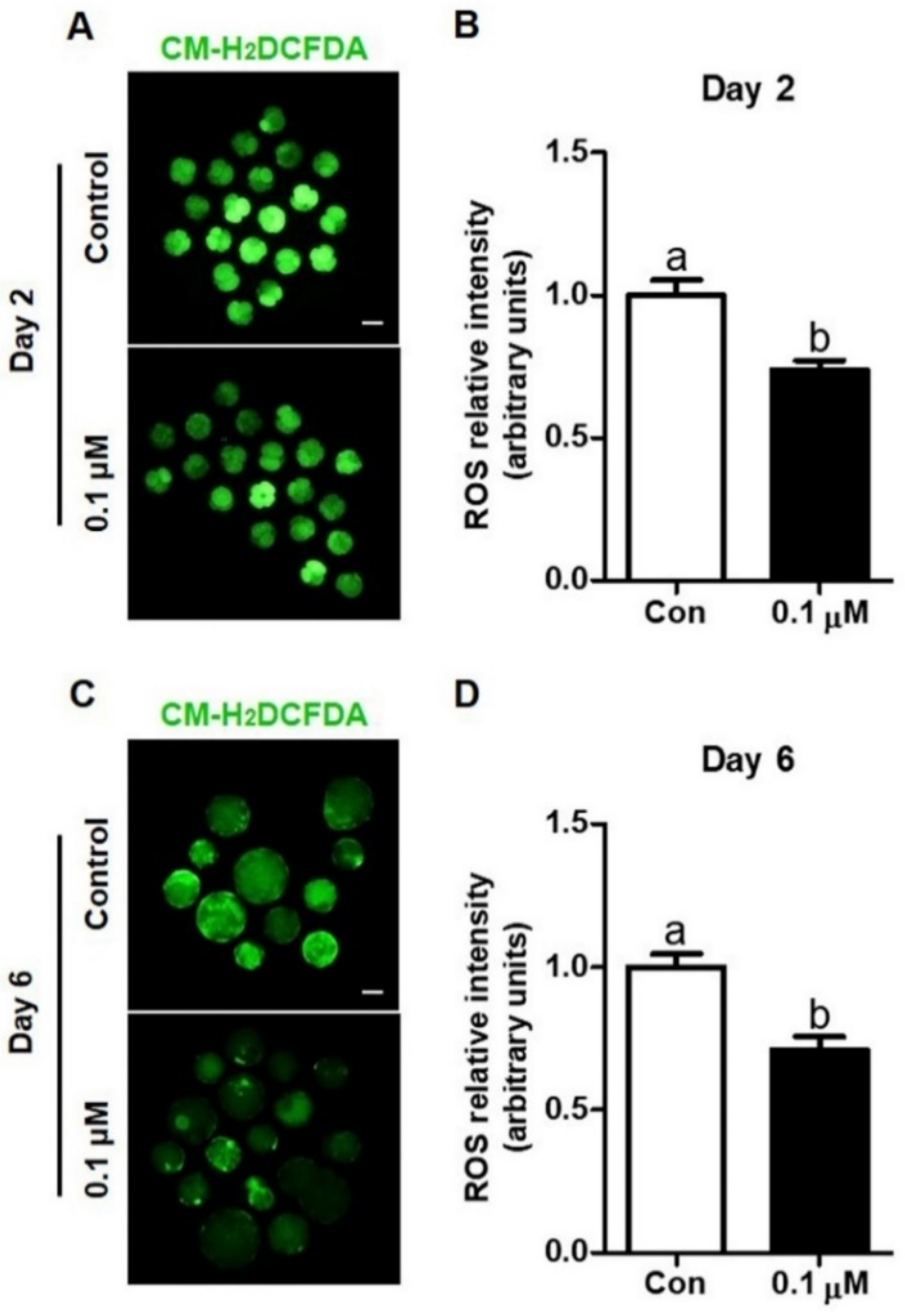
3.2. Effect of Lycopene on Intracellular ROS Levels in IVF Embryos
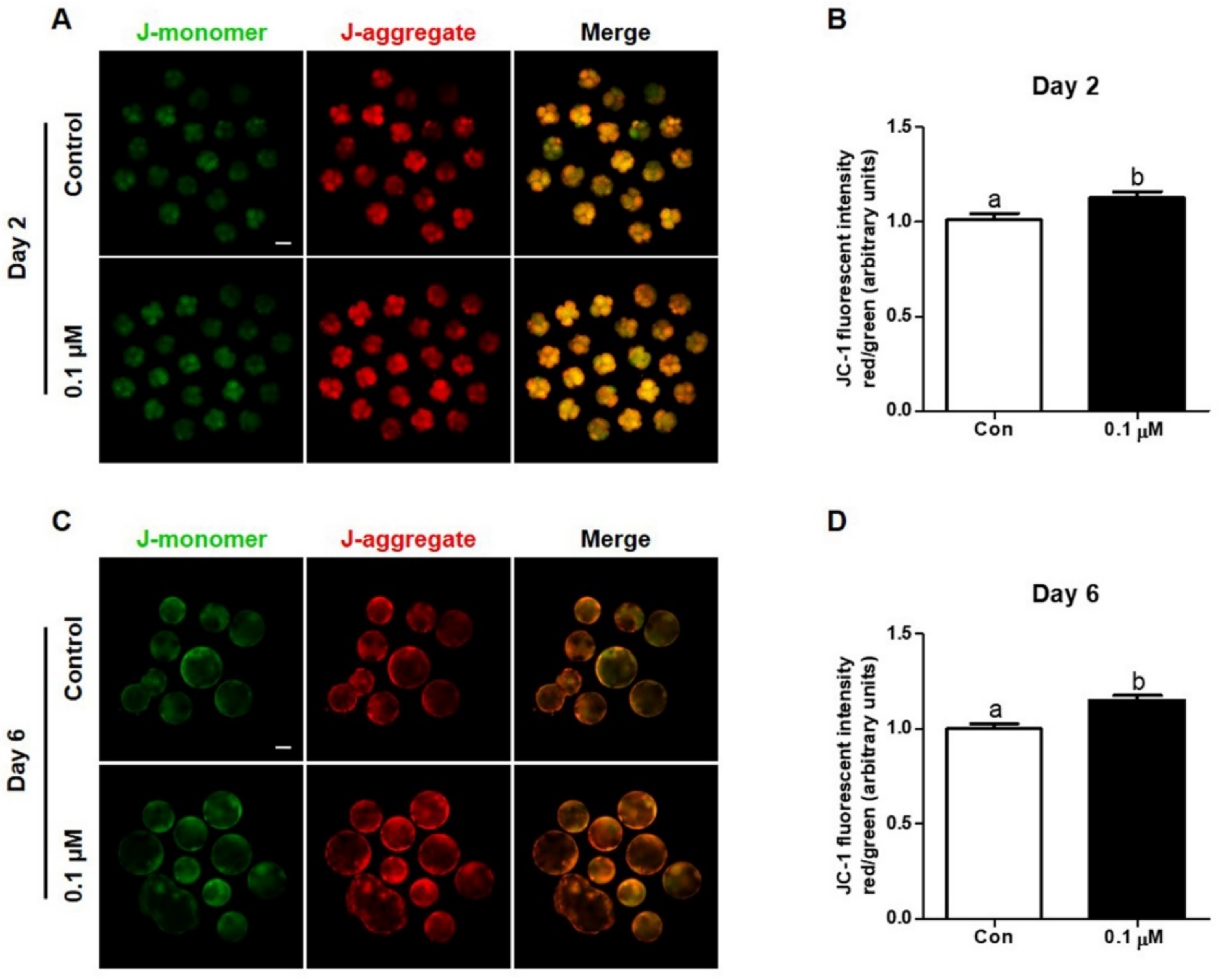
3.3. Effect of Lycopene on Mitochondrial Membrane Potential in IVF Embryos
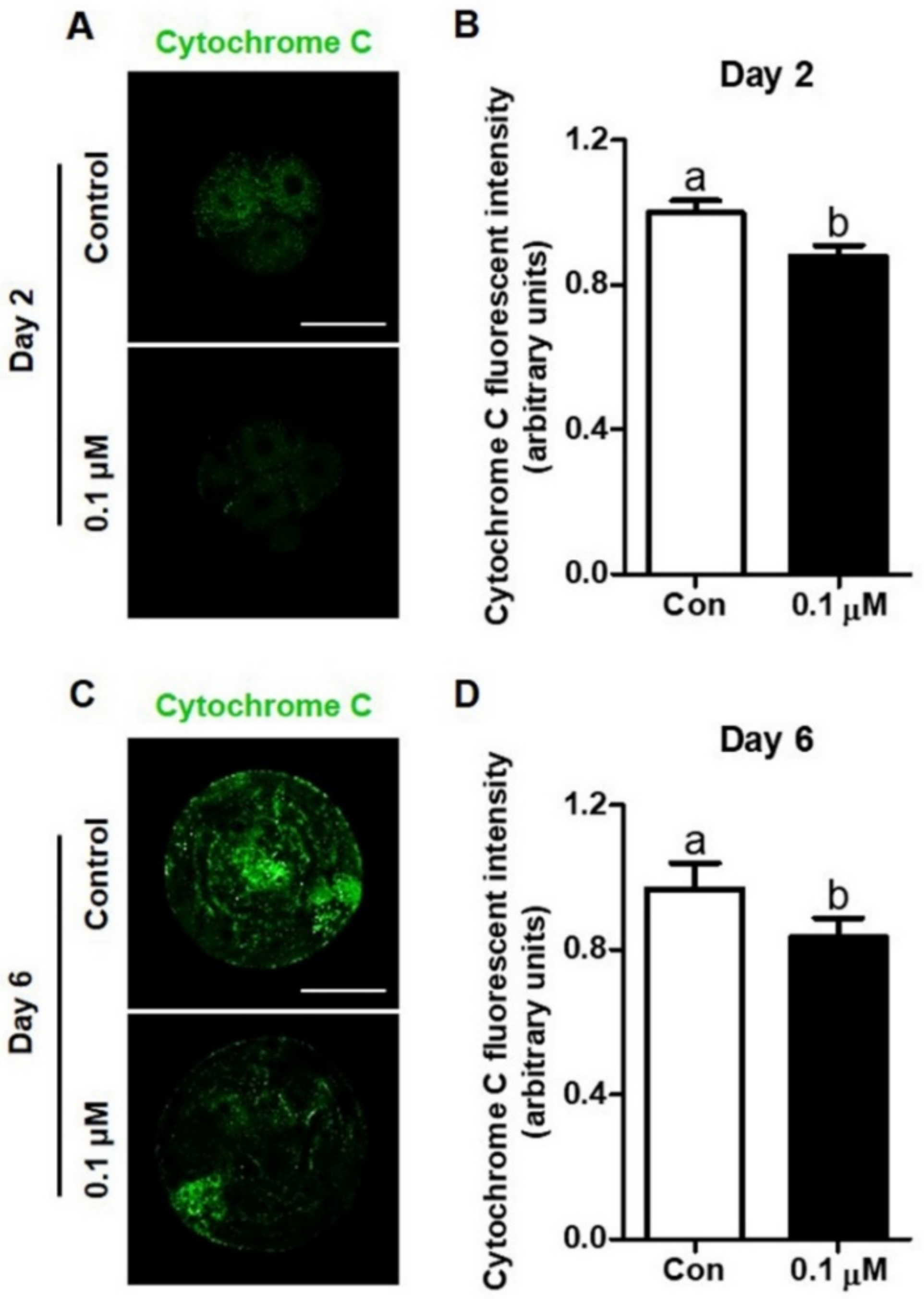
3.4. Effect of Lycopene on Cytochrome C Expression in IVF Embryos
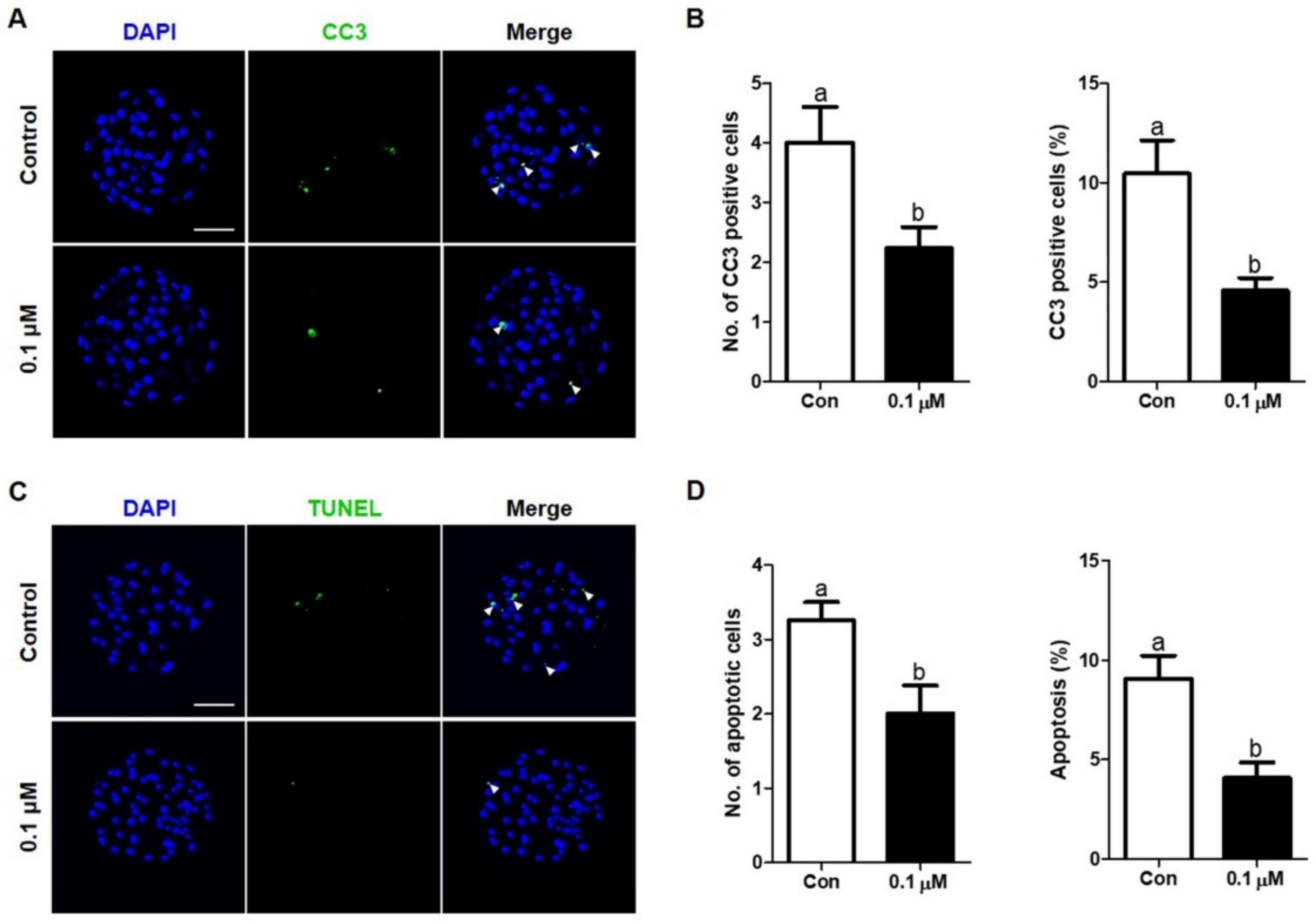
3.5. Effect of Lycopene on the Expression of Cleaved Caspase-3 and Apoptosis Levels in IVF Embryos
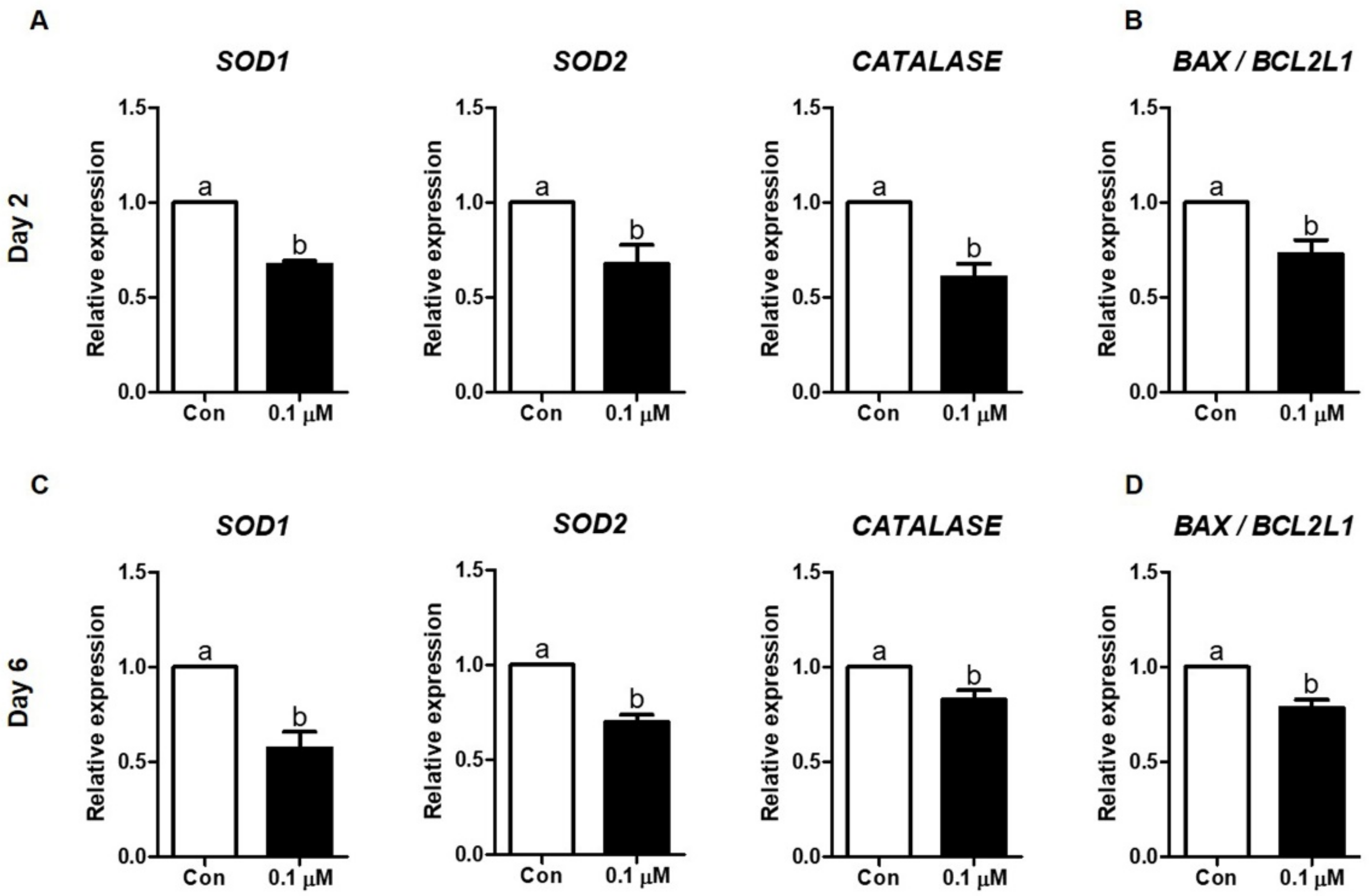
3.6. Effect of Lycopene on the Expression of Antioxidant Enzyme- and Apoptosis-Related mRNAs in IVF Embryos
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Walters, E.M.; Prather, R.S. Advancing swine models for human health and diseases. Mo. Med. 2013, 110, 212–215. [Google Scholar] [PubMed]
- Agarwal, A.; Durairajanayagam, D.; du Plessis, S.S. Utility of antioxidants during assisted reproductive techniques: An evidence based review. Reprod. Biol. Endocrinol. 2014, 12, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Long, H.; Lyu, Q.F.; Zhang, Q.H.; Yan, Z.G.; Liang, H.X.; Chai, W.R.; Yan, Z.; Kuang, Y.P.; Qi, C. Protective effect of quercetin on the development of preimplantation mouse embryos against hydrogen peroxide-induced oxidative injury. PLoS ONE 2014, 9, e89520. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.A.; Stephens, K.K.; Winuthayanon, W. Extracellular Vesicles and the Oviduct Function. Int. J. Mol. Sci. 2020, 21, 8280. [Google Scholar] [CrossRef]
- Poljsak, B.; Suput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef]
- Khazaei, M.; Aghaz, F. Reactive Oxygen Species Generation and Use of Antioxidants during In Vitro Maturation of Oocytes. Int. J. Fertil. Steril. 2017, 11, 63–70. [Google Scholar]
- Farnezi, H.C.M.; Goulart, A.C.X.; Santos, A.D.; Ramos, M.G.; Penna, M.L.F. Three-parent babies: Mitochondrial replacement therapies. JBRA Assist. Reprod. 2020, 24, 189–196. [Google Scholar] [CrossRef]
- Vakifahmetoglu-Norberg, H.; Ouchida, A.T.; Norberg, E. The role of mitochondria in metabolism and cell death. Biochem. Biophys. Res. Commun. 2017, 482, 426–431. [Google Scholar] [CrossRef]
- Ong, S.B.; Gustafsson, A.B. New roles for mitochondria in cell death in the reperfused myocardium. Cardiovasc. Res. 2012, 94, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.G.; Park, H.J.; Kim, J.W.; Jung, J.M.; Kim, M.J.; Jegal, H.G.; Kim, I.S.; Kang, M.J.; Wee, G.; Yang, H.Y.; et al. Mito-TEMPO improves development competence by reducing superoxide in preimplantation porcine embryos. Sci. Rep. 2018, 8, 10130. [Google Scholar] [CrossRef] [PubMed]
- Colombini, M. Ceramide channels and mitochondrial outer membrane permeability. J. Bioenerg. Biomembr. 2017, 49, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Seervi, M.; Joseph, J.; Sobhan, P.K.; Bhavya, B.C.; Santhoshkumar, T.R. Essential requirement of cytochrome c release for caspase activation by procaspase-activating compound defined by cellular models. Cell Death Dis. 2011, 2, e207. [Google Scholar] [CrossRef] [PubMed]
- Do, L.T.; Luu, V.V.; Morita, Y.; Taniguchi, M.; Nii, M.; Peter, A.T.; Otoi, T. Astaxanthin present in the maturation medium reduces negative effects of heat shock on the developmental competence of porcine oocytes. Reprod. Biol. 2015, 15, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cheng, D.; Gao, X.; Bao, J.; Ma, X.; Wang, H. Vitamin C enhances the in vitro development of porcine pre-implantation embryos by reducing oxidative stress. Reprod. Domest. Anim. 2012, 47, 873–879. [Google Scholar] [CrossRef]
- Lee, S.; Park, E.J.; Moon, J.H.; Kim, S.J.; Song, K.; Lee, B.C. Sequential treatment with resveratrol-trolox improves development of porcine embryos derived from parthenogenetic activation and somatic cell nuclear transfer. Theriogenology 2015, 84, 145–154. [Google Scholar] [CrossRef]
- You, J.; Kim, J.; Lim, J.; Lee, E. Anthocyanin stimulates in vitro development of cloned pig embryos by increasing the intracellular glutathione level and inhibiting reactive oxygen species. Theriogenology 2010, 74, 777–785. [Google Scholar] [CrossRef]
- Liang, S.; Jin, Y.X.; Yuan, B.; Zhang, J.B.; Kim, N.H. Melatonin enhances the developmental competence of porcine somatic cell nuclear transfer embryos by preventing DNA damage induced by oxidative stress. Sci. Rep. 2017, 7, 11114. [Google Scholar] [CrossRef] [Green Version]
- Pirayesh Islamian, J.; Mehrali, H. Lycopene as a carotenoid provides radioprotectant and antioxidant effects by quenching radiation-induced free radical singlet oxygen: An overview. Cell J. 2015, 16, 386–391. [Google Scholar]
- Tamilselvan, P.; Langeswaran, K.; Vijayaprakash, S.; Kumar, S.G.; Revathy, R.; Balasubramanian, M.P. Efficiency of lycopene against reproductive and developmental toxicity of Bisphenol A in male Sprague Dawley rats. Biomed. Prev. Nutr. 2014, 4, 491–498. [Google Scholar] [CrossRef]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M.; et al. Lycopene as a Natural Antioxidant Used to Prevent Human Health Disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, A.K.; Chopra, K. Lycopene abrogates Abeta(1-42)-mediated neuroinflammatory cascade in an experimental model of Alzheimer’s disease. J. Nutr. Biochem. 2015, 26, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, H.; Cantley, T.C.; Stumpf, T.T.; Terlouw, S.L.; Day, B.N. In vitro development of in vitro-matured porcine oocytes following chemical activation or in vitro fertilization. Biol. Reprod. 1994, 50, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, H.; Cantley, T.C.; Day, B.N. Synchronization of meiosis in porcine oocytes by exposure to dibutyryl cyclic adenosine monophosphate improves developmental competence following in vitro fertilization. Biol. Reprod. 1997, 57, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Machaty, Z.; Day, B.N.; Prather, R.S. Development of early porcine embryos in vitro and in vivo. Biol. Reprod. 1998, 59, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Nasr-Esfahani, M.H.; Aitken, J.R.; Johnson, M.H. Hydrogen peroxide levels in mouse oocytes and early cleavage stage embryos developed in vitro or in vivo. Development 1990, 109, 501–507. [Google Scholar]
- Kim, J.S.; Cho, Y.S.; Song, B.S.; Wee, G.; Park, J.S.; Choo, Y.K.; Yu, K.; Lee, K.K.; Han, Y.M.; Koo, D.B. Exogenous dibutyryl cAMP affects meiotic maturation via protein kinase A activation; it stimulates further embryonic development including blastocyst quality in pigs. Theriogenology 2008, 69, 290–301. [Google Scholar] [CrossRef]
- Kim, N.H.; Simerly, C.; Funahashi, H.; Schatten, G.; Day, B.N. Microtubule organization in porcine oocytes during fertilization and parthenogenesis. Biol. Reprod. 1996, 54, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Long, C.R.; Dobrinsky, J.R.; Garrett, W.M.; Johnson, L.A. Dual labeling of the cytoskeleton and DNA strand breaks in porcine embryos produced in vivo and in vitro. Mol. Reprod. Dev. 1998, 51, 59–65. [Google Scholar] [CrossRef]
- Kuijk, E.W.; du Puy, L.; van Tol, H.T.; Haagsman, H.P.; Colenbrander, B.; Roelen, B.A. Validation of reference genes for quantitative RT-PCR studies in porcine oocytes and preimplantation embryos. BMC Dev. Biol. 2007, 7, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cagnone, G.; Sirard, M.A. The embryonic stress response to in vitro culture: Insight from genomic analysis. Reproduction 2016, 152, R247–R261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweigert, F.J.; Steinhagen, B.; Raila, J.; Siemann, A.; Peet, D.; Buscher, U. Concentrations of carotenoids, retinol and alpha-tocopherol in plasma and follicular fluid of women undergoing IVF. Hum. Reprod. 2003, 18, 1259–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissanen, T.H.; Voutilainen, S.; Nyyssonen, K.; Salonen, R.; Kaplan, G.A.; Salonen, J.T. Serum lycopene concentrations and carotid atherosclerosis: The Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin. Nutr. 2003, 77, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kausar, S.; Wang, F.; Cui, H. The Role of Mitochondria in Reactive Oxygen Species Generation and Its Implications for Neurodegenerative Diseases. Cells 2018, 7, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siristatidis, C.; Vogiatzi, P.; Varounis, C.; Askoxylaki, M.; Chrelias, C.; Papantoniou, N. The Effect of Reactive Oxygen Species on Embryo Quality in IVF. In Vivo 2016, 30, 149–153. [Google Scholar] [PubMed]
- Bae, Y.S.; Oh, H.; Rhee, S.G.; Yoo, Y.D. Regulation of reactive oxygen species generation in cell signaling. Mol. Cells 2011, 32, 491–509. [Google Scholar] [CrossRef] [Green Version]
- Bain, N.T.; Madan, P.; Betts, D.H. The early embryo response to intracellular reactive oxygen species is developmentally regulated. Reprod. Fertil. Dev. 2011, 23, 561–575. [Google Scholar] [CrossRef]
- Li, X.X.; Lee, K.B.; Lee, J.H.; Kim, K.J.; Kim, E.Y.; Han, K.W.; Park, K.S.; Yu, J.; Kim, M.K. Glutathione and cysteine enhance porcine preimplantation embryo development in vitro after intracytoplasmic sperm injection. Theriogenology 2014, 81, 309–314. [Google Scholar] [CrossRef]
- Mun, S.E.; Sim, B.W.; Yoon, S.B.; Jeong, P.S.; Yang, H.J.; Choi, S.A.; Park, Y.H.; Kim, Y.H.; Kang, P.; Jeong, K.J.; et al. Dual effect of fetal bovine serum on early development depends on stage-specific reactive oxygen species demands in pigs. PLoS ONE 2017, 12, e0175427. [Google Scholar] [CrossRef] [Green Version]
- Santulli, G.; Marks, A.R. Essential Roles of Intracellular Calcium Release Channels in Muscle, Brain, Metabolism, and Aging. Curr. Mol. Pharmacol. 2015, 8, 206–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez-Jurado, S.; Diaz-Colunga, J.; das Neves, R.P.; Martinez-Lorente, A.; Almazan, F.; Guantes, R.; Iborra, F.J. Mitochondrial levels determine variability in cell death by modulating apoptotic gene expression. Nat. Commun. 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Babayev, E.; Seli, E. Oocyte mitochondrial function and reproduction. Curr. Opin. Obstet. Gynecol. 2015, 27, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romek, M.; Gajda, B.; Rolka, M.; Smorag, Z. Mitochondrial activity and morphology in developing porcine oocytes and pre-implantation non-cultured and cultured embryos. Reprod. Domest. Anim. 2011, 46, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, S.L.; Lane, M.; Mitchell, M. Impaired mitochondrial function in the preimplantation embryo perturbs fetal and placental development in the mouse. Biol. Reprod. 2011, 84, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhang, H.; Zhang, Y.; Yang, Q.; Zhao, S. Caspase-dependent mitochondrial apoptotic pathway is involved in astilbin-mediated cytotoxicity in breast carcinoma cells. Oncol. Rep. 2018, 40, 2278–2286. [Google Scholar] [CrossRef] [PubMed]
- Kimura, N.; Tsunoda, S.; Iuchi, Y.; Abe, H.; Totsukawa, K.; Fujii, J. Intrinsic oxidative stress causes either 2-cell arrest or cell death depending on developmental stage of the embryos from SOD1-deficient mice. Mol. Hum. Reprod. 2010, 16, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wen, C.; Yang, M.; Gan, D.; Fan, C.; Li, A.; Li, Q.; Zhao, J.; Zhu, L.; Lu, D. Lycopene protects against t-BHP-induced neuronal oxidative damage and apoptosis via activation of the PI3K/Akt pathway. Mol. Biol. Rep. 2019, 46, 3387–3397. [Google Scholar] [CrossRef] [PubMed]
- Buettner, G.R. Superoxide dismutase in redox biology: The roles of superoxide and hydrogen peroxide. Anticancer Agents Med. Chem. 2011, 11, 341–346. [Google Scholar] [CrossRef]
- Khan, S.N.; Shaeib, F.; Najafi, T.; Kavdia, M.; Gonik, B.; Saed, G.M.; Goud, P.T.; Abu-Soud, H.M. Diffused Intra-Oocyte Hydrogen Peroxide Activates Myeloperoxidase and Deteriorates Oocyte Quality. PLoS ONE 2015, 10, e0132388. [Google Scholar] [CrossRef] [Green Version]
- Waiz, S.A.; Raies-Ul-Haq, M.; Dhanda, S.; Kumar, A.; Goud, T.S.; Chauhan, M.S.; Upadhyay, R.C. Heat stress and antioxidant enzyme activity in bubaline (Bubalus bubalis) oocytes during in vitro maturation. Int. J. Biometeorol. 2016, 60, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Arias-Alvarez, M.; Garcia-Garcia, R.M.; Lopez-Tello, J.; Rebollar, P.G.; Gutierrez-Adan, A.; Lorenzo, P.L. In vivo and in vitro maturation of rabbit oocytes differently affects the gene expression profile, mitochondrial distribution, apoptosis and early embryo development. Reprod. Fertil. Dev. 2017, 29, 1667–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′-3′) | Accession Number |
|---|---|---|
| GAPDH | F-CCCTGAGACACGATGGTGAA | NM_001206359.1 |
| R-GGAGGTCAATGAAGGGGTCA | ||
| SOD1 | F-GGTGGGCCAAAGGATCAAGA | NM_001190422.1 |
| R-TACACAGTGGCCACACCATC | ||
| SOD2 | F-GGTGGAGGCCACATCAATCA | NM_214127.2 |
| R-AACAAGGGCAATCTGCAAG | ||
| CATALASE | F-TGTACCCGCTATTCTGGGGA | NM_214301.2 |
| R-TCACACAGGCGTTTCCTCTC | ||
| BAX | F-CGATCTCGAAGGAAGTCCAG | XM_003127290.5 |
| R-AAGCGCATTGGAGATGAACT | ||
| BCL2L1 | F-AGGGCATTCAGTGACCTGAC | NM_214285.1 |
| R-CGATCCGACTCACCAATACC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.-G.; Lee, S.; Jeong, P.-S.; Kim, M.J.; Park, S.-H.; Joo, Y.E.; Park, S.H.; Song, B.-S.; Kim, S.-U.; Kim, M.K.; et al. Lycopene Improves In Vitro Development of Porcine Embryos by Reducing Oxidative Stress and Apoptosis. Antioxidants 2021, 10, 230. https://doi.org/10.3390/antiox10020230
Kang H-G, Lee S, Jeong P-S, Kim MJ, Park S-H, Joo YE, Park SH, Song B-S, Kim S-U, Kim MK, et al. Lycopene Improves In Vitro Development of Porcine Embryos by Reducing Oxidative Stress and Apoptosis. Antioxidants. 2021; 10(2):230. https://doi.org/10.3390/antiox10020230
Chicago/Turabian StyleKang, Hyo-Gu, Sanghoon Lee, Pil-Soo Jeong, Min Ju Kim, Soo-Hyun Park, Ye Eun Joo, Sung Hyun Park, Bong-Seok Song, Sun-Uk Kim, Min Kyu Kim, and et al. 2021. "Lycopene Improves In Vitro Development of Porcine Embryos by Reducing Oxidative Stress and Apoptosis" Antioxidants 10, no. 2: 230. https://doi.org/10.3390/antiox10020230
APA StyleKang, H. -G., Lee, S., Jeong, P. -S., Kim, M. J., Park, S. -H., Joo, Y. E., Park, S. H., Song, B. -S., Kim, S. -U., Kim, M. K., & Sim, B. -W. (2021). Lycopene Improves In Vitro Development of Porcine Embryos by Reducing Oxidative Stress and Apoptosis. Antioxidants, 10(2), 230. https://doi.org/10.3390/antiox10020230