
Physicochemical, Antioxidant, and Anti-Inflammatory Properties of Rapeseed Lecithin Liposomes Loading a Chia (Salvia hispanica L.) Seed Extract

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
Obtaining of Chia Seed Extract
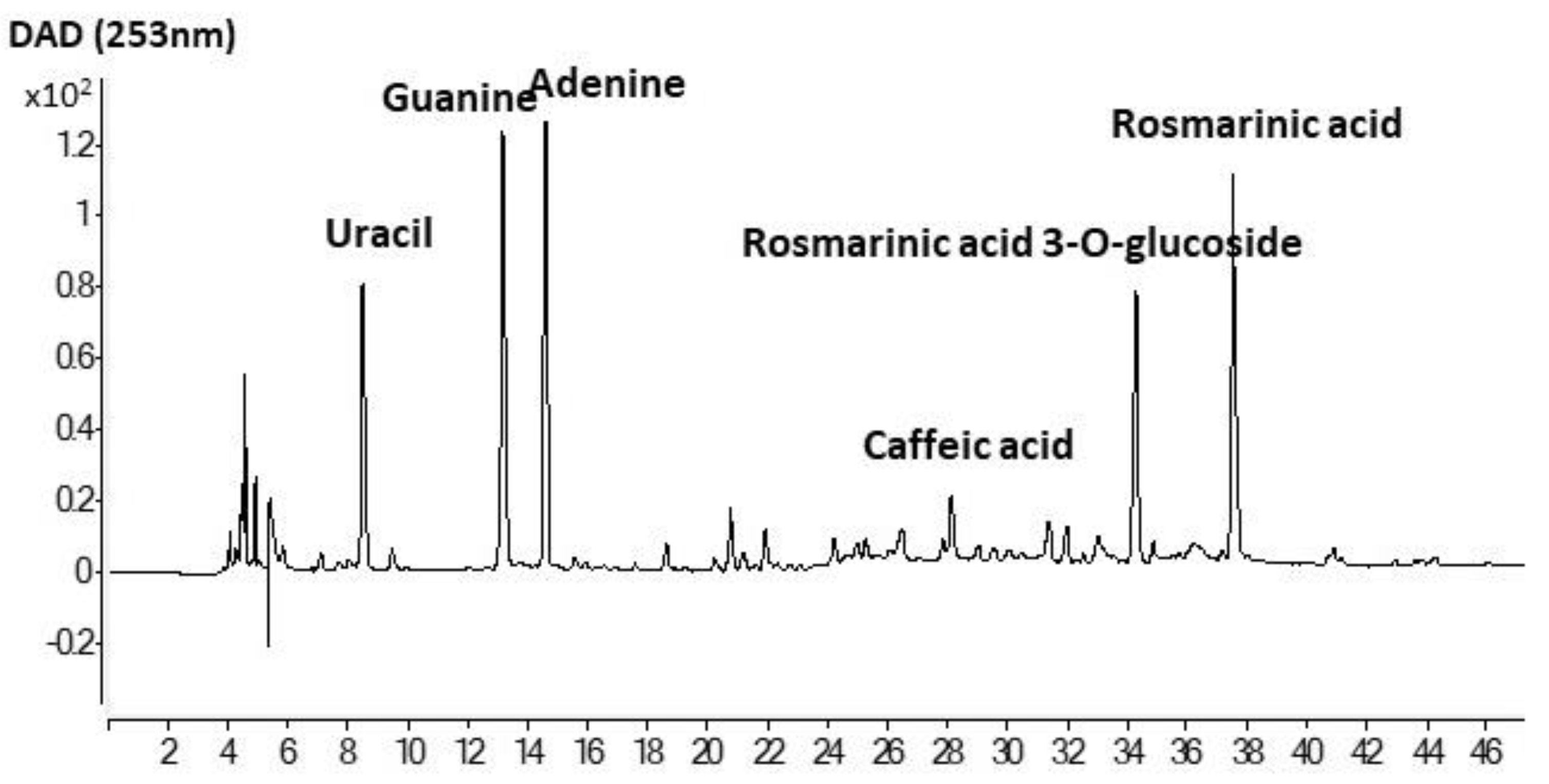
2.2. Identification of Phenolic Compounds by HPLC-ESI-QTOF Analysis
2.3. Preparation of Liposomes
2.4. Chemical Composition of LEC and PPL
2.5. Infrared Spectroscopy (ATR-FTIR)
2.6. Differential Scanning Calorimetry (DSC)
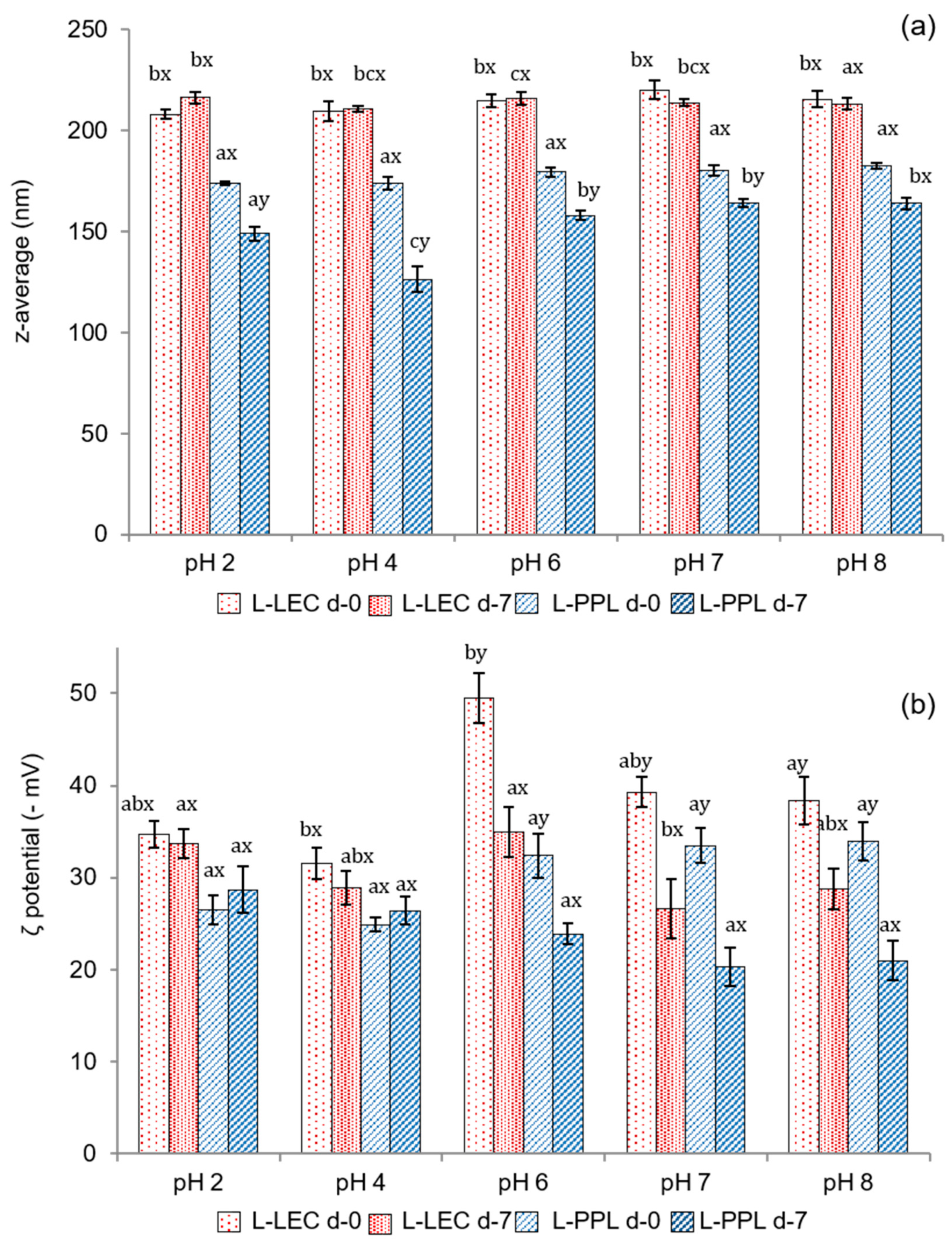
2.7. Liposome Hydrodynamic Properties
2.8. Entrapment Efficiency
2.9. Antioxidant Properties
2.10. Cytotoxicity and Anti-Inflammatory Properties
2.10.1. Cell Culture and Cell Viability
2.10.2. Immunostimulation
2.11. Statistical Analysis
3. Results and Discussion
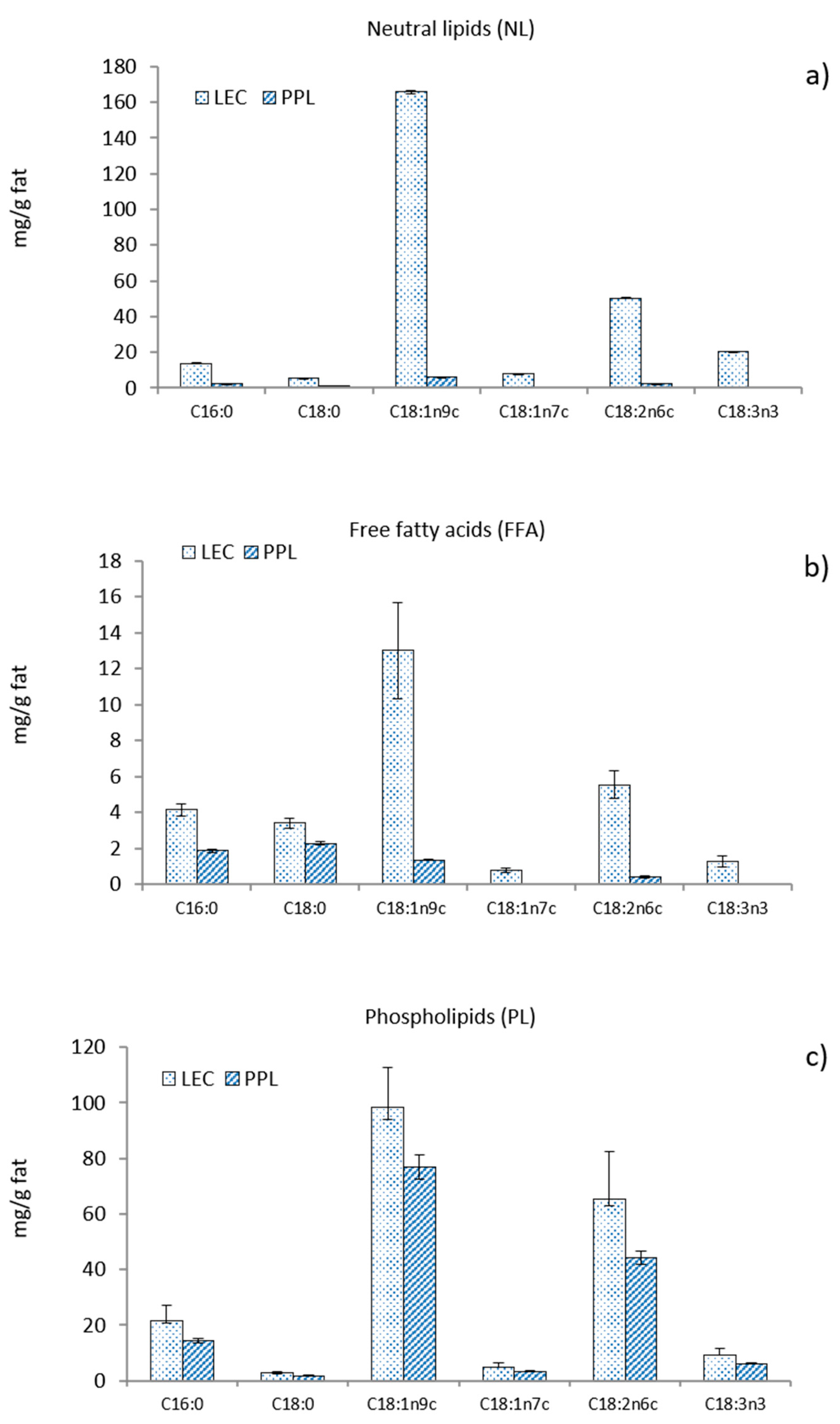
3.1. Fatty Acid Composition
3.2. Sterols, Tocopherols and Amino Acid Composition
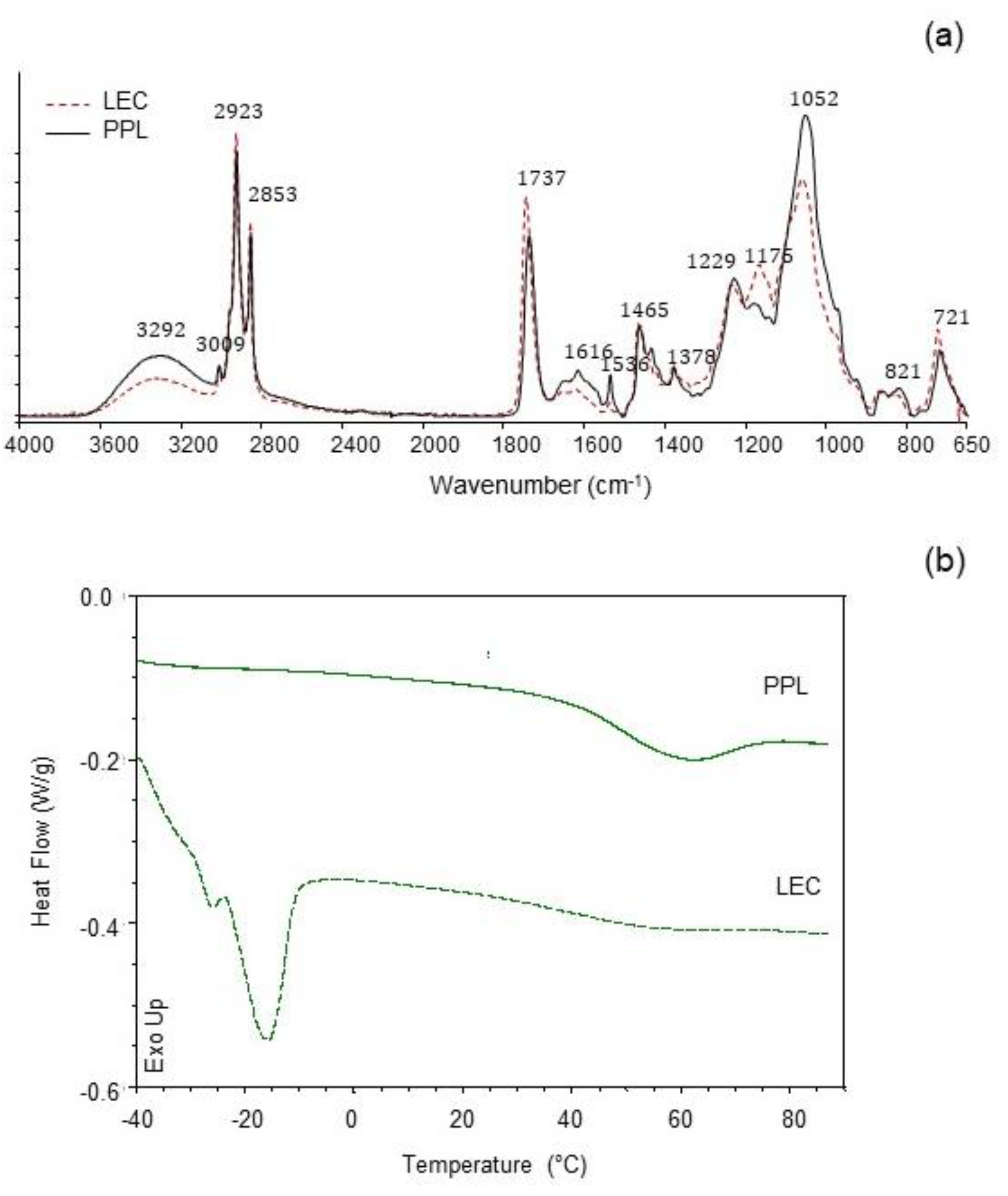
3.3. Infrared Spectroscopy (ATR-FTIR)
3.4. Thermal Properties
3.5. Liposomal Encapsulation
3.6. Antioxidant Activity and Phenolic Composition
3.7. Cytotoxicity
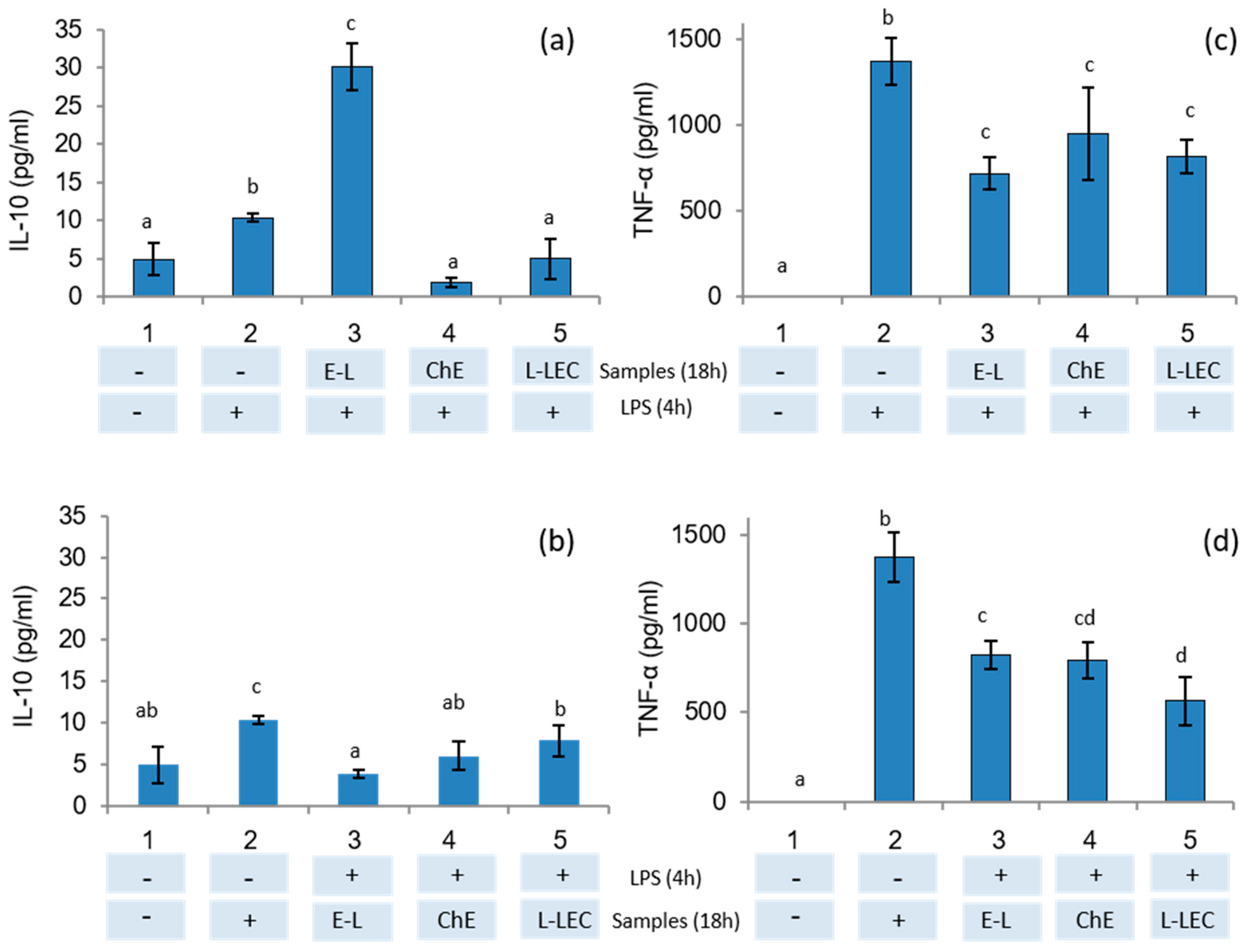
3.8. Anti-Inflammatory Activity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Marín, D.; Alemán, A.; Sánchez-Faure, A.; Montero, P.; Gómez-Guillén, M.C. Freeze-dried phosphatidylcholine liposomes encapsulating various antioxidant extracts from natural waste as functional ingredients in surimi gels. Food Chem. 2018, 245, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.M.; Xue, Y.; Sun, S.H.; Wen, M.; Li, Z.J.; Xu, J.; Wang, J.F.; Yanagita, T.; Wang, Y.M.; Xue, C.H. Effects of different fatty acids composition of phosphatidylcholine on brain function of dementia mice induced by scopolamine. Lipids Health Dis. 2016, 15, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara, D.; Shene, C. Encapsulation of lactoferrin into rapeseed phospholipids based liposomes: Optimization and physicochemical characterization. J. Food Eng. 2019, 262, 29–38. [Google Scholar] [CrossRef]
- Paini, M.; Daly, S.R.; Aliakbarian, B.; Fathi, A.; Tehrany, E.A.; Perego, P.; Valtchev, P. An efficient liposome based method for antioxidants encapsulation. Colloids Surf. B Biointerfaces 2015, 136, 1067–1072. [Google Scholar] [CrossRef]
- Tehrany, E.A.; Kahn, C.J.F.; Baravian, C.; Maherani, B.; Belhaj, N.; Wang, X.; Linder, M. Elaboration and characterization of nanoliposome made of soya; rapeseed and salmon lecithins: Application to cell culture. Colloids Surf. B Biointerfaces 2012, 95, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, T. Oxidative stability of egg and soy lecithin as affected by transition metal ions and pH in emulsion. J. Agric. Food Chem. 2008, 56, 11424–11431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biltonen, R.L.; Lichtenberg, D. The use of differential scanning calorimetry as a tool to characterize liposome preparations. Chem. Phys. Lipids 1993, 64, 129–142. [Google Scholar] [CrossRef]
- Borst, J.W.; Visser, N.V.; Kouptsova, O.; Visser, A.J.W.G. Oxidation of unsaturated phospholipids in membrane bilayer mixtures is accompanied by membrane fluidity changes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1487, 61–73. [Google Scholar] [CrossRef]
- Butina, E.A.; Gerasimenko, E.O.; Bugaets, I.A.; Kopteva, A.A. Comparative assessment of plant lecithins as the technologically functional components for creating encapsulated biologically active substances. Int. J. Pharm. Res. 2018, 10, 297–304. [Google Scholar]
- Taladrid, D.; Marín, D.; Alemán, A.; Álvarez-Acero, I.; Montero, P.; Gómez-Guillén, M.C. Effect of chemical composition and sonication procedure on properties of food-grade soy lecithin liposomes with added glycerol. Food Res. Int. 2017, 100, 541–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenzel, M.; Steffen-Heins, A. Impact of quercetin and fish oil encapsulation on bilayer membrane and oxidation stability of liposomes. Food Chem. 2015, 185, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Fernández-López, J.; Lucas-González, R.; Viuda-Martos, M.; Sayas-Barberá, E.; Pérez-Álvarez, J.A. Chia oil extraction coproduct as a potential new ingredient for the food industry: Chemical, physicochemical, techno-functional and antioxidant properties. Plant. Foods Hum. Nutr. 2018, 73, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Alcântara, M.A.; de Lima Brito Polari, I.; de Albuquerque Meireles, B.R.L.; de Lima, A.E.A.; da Silva Junior, J.C.; de Andrade Vieira, É.; dos Santos, N.A.; de Magalhães Cordeiro, A.M.T. Effect of the solvent composition on the profile of phenolic compounds extracted from chia seeds. Food Chem. 2019, 275, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.D.; Amaral, Q.D.F.D.; Duarte, J.A.; Limberger, J.T.; Chaves, P.E.E.; Zuravski, L.; Oliveira, L.F.S.D.; Machado, M.M. Antigenotoxic, antimutagenic and cytoprotective potential of Salvia hispanica L. seed extract on human leukocytes exposed to oxidative damage. J. Funct. Foods 2017, 38, 505–509. [Google Scholar] [CrossRef]
- Pigni, N.B.; Aranibar, C.; Lucini Mas, A.; Aguirre, A.; Borneo, R.; Wunderlin, D.; Baroni, M.V. Chemical profile and bioaccessibility of polyphenols from wheat pasta supplemented with partially-deoiled chia flour. LWT-Food Sci. Technol. 2020, 124, 109134. [Google Scholar] [CrossRef]
- Teoh, S.L.; Lai, N.M.; Vanichkulpitak, P.; Vuksan, V.; Ho, H.; Chaiyakunapruk, N. Clinical evidence on dietary supplementation with chia seed (Salvia hispanica L.): A systematic review and meta-analysis. Nutr. Rev. 2018, 76, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Alemán, A.; Marín, D.; Taladrid, D.; Montero, P.; Gómez-Guillén, M.C. Encapsulation of antioxidant sea fennel (Crithmum maritimum) aqueous and ethanolic extracts in freeze-dried soy phosphatidylcholine liposomes. Food Res. Int. 2019, 119, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Ahn, D.U. Simultaneous Analysis of Tocopherols, Cholesterol, and Phytosterols Using Gas Chromatography. J. Food Sci. 2002, 6, 1696–1700. [Google Scholar] [CrossRef]
- Lund, M.A.; Bronwyn, A.J.T.; O’Brien, S.D. The choice of phorbol 12-myristate 13-acetate differentiation protocol influences the response of THP-1 macrophages to a pro-inflammatory stimulus. J. Immunol. Methods 2016, 430, 64–70. [Google Scholar] [CrossRef]
- Van Nieuwenhuyzen, W.; Tomás, M.C. Update on vegetable lecithin and phospholipid technologies. Eur. J. Lipid Sci. Technol. 2008, 110, 472–486. [Google Scholar] [CrossRef]
- Xu, B.; Zhang, L.; Ma, F.; Zhang, W.; Wang, X.; Zhang, Q.; Luo, D.; Ma, H.; Li, P. Determination of free steroidal compounds in vegetable oils by comprehensive two-dimensional gas chromatography coupled to time-of-flight mass spectrometry. Food Chem. 2018, 245, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Sakouhi, F.; Absalon, C.; Harrabi, S.; Vitry, C.; Boukhcina, S.; Fouquet, E.; Kallel, H. Dynamic accumulation of 4-desmethylsterols and phytostanols during ripening of Tunisian olives (Olea europea L). Food Chem. 2009, 112, 897–902. [Google Scholar] [CrossRef]
- Jones, P.J.; Macdougall, D.E.; Ntanios, F.; Vanstone, C.A. Dietary phytosterols ascholesterol-lowering agents in humans. Can. J. Physiol. Pharmacol. 1997, 75, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Bouic, P.J.D. The role of phytosterols and phytosterolins in immune modulation: A review of the past 10 years. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 471–475. [Google Scholar] [CrossRef]
- Unsalan, C.A.; Sahin, I.; Kazanci, N. A concentration dependent spectroscopic study of binary mixtures of plant sterol stigmasterol and zwitterionic dimyristoyl phosphatidylcholine multilamellar vesicles: An FTIR study. J. Mol. Struct. 2018, 1174, 127–132. [Google Scholar] [CrossRef]
- Hielscher, R.; Wenz, T.; Hunte, C.; Hellwig, P. Monitoring the redox and protonation dependent contributions of cardiolipin in electrochemically induced FTIR difference spectra of the cytochrome bc1 complex from yeast. Biochim. Biophys. Acta Bioenerg 2009, 1787, 617–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Ye, Q.; Pan, Q.; Ding, Y.; Wei, M.; Liu, Y.; Van De Voort, F.R. Total phospholipids in edible oils by in-vial solvent extraction coupled with FTIR analysis. J. Agric. Food Chem. 2014, 62, 3101–3107. [Google Scholar] [CrossRef] [PubMed]
- Hübner, W.; Mantsch, H.H.; Kates, M. Intramolecular hydrogen bonding in cardiolipin. Biochim. Biophys. Acta Biomembr. 1991, 1066, 166–174. [Google Scholar] [CrossRef]
- Lee, D.C.; Miller, I.R.; Chapman, D. An infrared spectroscopic study of metastable and stable forms of hydrated cerebroside bilayers. Biochim. Biophys. Acta Biomembr. 1986, 859, 266–270. [Google Scholar] [CrossRef]
- Xie, M.; Dunford, N.T. Lipid composition and emulsifying properties of canola lecithin from enzymatic degumming. Food Chem. 2017, 218, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Briggs, J.L. Rheological and thermal properties of soybean oils with modified FA compositions. J. Am. Oil Chem. Soc. 2002, 79, 831–836. [Google Scholar] [CrossRef]
- Tan, C.P.; Che Man, Y.B. Differential scanning calorimetric analysis of edible oils: Comparison of thermal properties and chemical composition. J. Am. Oil Chem. Soc. 2000, 77, 143–155. [Google Scholar] [CrossRef]
- Goñi, F.M.; Alonso, A. Biophysics of sphingolipids I. Membrane properties of sphingosine, ceramides and other simple sphingolipids. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1902–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelwahed, W.; Degobert, G.; Stainmesse, S.; Fessi, H. Freeze-drying of nanoparticles: Formulation, process and storage considerations. Adv. Drug Deliv. Rev. 2006, 58, 1688–1713. [Google Scholar] [CrossRef] [PubMed]
- Tai, K.; Liu, F.; He, X.; Ma, P.; Mao, L.; Gao, Y.; Yuan, F. The effect of sterol derivatives on properties of soybean and egg yolk lecithin liposomes: Stability, structure and membrane characteristics. Food Res. Int. 2018, 109, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.J.; Costa de Camargo, A.; Shahidi, F. Phenolic and polyphenolic profiles of chia seeds and their in vitro biological activities. J. Funct. Foods 2017, 35, 622–634. [Google Scholar] [CrossRef]
- Bruno, M.C.; Posik, D.M.; Zappa, M.E.; Baroni, V.; Wunderlin, D.; Giovambattista, G.; Peral García, P. Genetic identification of flax, chia and sesame seeds in processed foods. Food Control. 2020, 118, 107374. [Google Scholar] [CrossRef]
- De Marco, S.; Sichetti, M.; Muradyan, D.; Piccioni, M.; Traina, G.; Pagiotti, R.; Pietrella, D. Probiotic cell-free supernatants exhibited anti-inflammatory and antioxidant activity on human gut epithelial cells and macrophages stimulated with LPS. Hindawi. Evid. Based Complement. Alternat. Med. 2018, 2018. [Google Scholar] [CrossRef]
- Ueki, K.; Tabeta, K.; Yoshie, H.; Yamazaki, K. Self-heat shock protein 60 induces tumour necrosis factor-α in monocyte-derived macrophage: Possible role in chronic inflammatory periodontal disease. Clin. Exp. Immunol. 2002, 127, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Savelkoul, H.F.J.; Wichers, H.J. Transcription profiles of LPS-stimulated THP-1 monocytes and macrophages: A tool to study inflammation modulating effects of food-derived compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.; Vidueiros, S.M.; Ayerza, R.; Coates, W.; Pallaro, A. Impact of chia (Salvia hispanica L.) on the immune system: Preliminary study. Proc. Nutr. Soc. 2008, 67, E12. [Google Scholar] [CrossRef] [Green Version]
- Ullah, R.; Nadeem, M.; Khalique, A.; Imran, M.; Mehmood, S.; Javid, A.; Hussain, J. Nutritional and therapeutic perspectives of Chia (Salvia hispanica L.): A review. J. Food Sci. Technol. 2016, 53, 1750–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LEC | PPL | |
|---|---|---|
| Fatty Acids (g/100 g) | ||
| C14:0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| C16:0 | 7.3 ± 0.0 | 9.6 ± 0.2 |
| C16:1n7 | 0.3 ± 0.0 | 0.4 ± 0.0 |
| C17:0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| C18:0 | 1.2 ± 0.0 | 0.9 ± 0.1 |
| C18:1n9c | 56.0 ± 0.4 | 51.8 ± 0.5 |
| C18:1n7c | 2.6 ± 0.0 | 2.3 ± 0.0 |
| C18:2n6c | 25.0 ± 0.1 | 29.9 ± 0.3 |
| C18:3n3 | 6.0 ± 0.0 | 4.3 ± 0.0 |
| C20:0 | 0.3 ± 0.0 | 0.1 ± 0.0 |
| C20:1n9 | 0.7 ± 0.0 | 0.2 ± 0.0 |
| C22:0 | 0.2 ± 0.0 | 0.1 ± 0.0 |
| C24:0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| ∑ SFA | 9.4 | 11.1 |
| ∑ MUFA | 59.6 | 54.7 |
| ∑ PUFA | 31.0 | 34.2 |
| Sterols (mg/100 g) | ||
| Campesterol | 223 ± 23.8 | 12.81 ± 1.37 |
| Stigmasterol | 5.83 ± 0.62 | 4.07 ± 0.44 |
| β-Sitosterol | 215 ± 23.0 | 15.23 ± 1.63 |
| Cholesterol | 2.08 ± 0.22 | n.d. |
| Cycloartenol | n.q. | n.q |
| Stigmasterol der. | n.q | n.d. |
| Tocopherols (mg/100 g) | ||
| γ-tocopherol | 18.1 ± 0.12 | 0.81 ± 0.18 |
| δ-tocopherol | 0.84 ± 0.01 | 0.04 ± 0.00 |
| α-tocopherol | 1.53 ± 0.09 | 0.03 ± 0.00 |
| Amino Acids (mg/100 g) | ||
| Asp | 2.70 ± 0.94 | 3.84 ± 0.07 |
| Thr | 0.75 ± 0.30 | 1.15 ± 0.32 |
| Ser | 1.86 ± 0.49 | 3.15 ± 0.28 |
| Glu | 3.68 ± 1.44 | 3.71 ± 0.21 |
| Gly | 1.42 ± 0.52 | 2.02 ± 0.06 |
| Ala | 1.04 ± 0.41 | 1.40 ± 0.10 |
| Cys | 0.46 ± 0.08 | 0.68 ± 0.05 |
| Val | 1.51 ± 0.51 | 2.23 ± 0.38 |
| Met | 0.38 ± 0.34 | 0.32 ± 0.05 |
| Ile | 0.88 ± 0.44 | 1.13 ± 0.20 |
| Leu | 1.52 ± 0.67 | 1.91 ± 0.29 |
| Tyr | 0.86 ± 0.31 | 0.87 ± 0.21 |
| Phe | 0.82 ± 0.29 | 1.48 ± 0.26 |
| His | 0.64 ± 0.29 | 1.05 ± 0.09 |
| Lys | 1.21 ± 0.36 | 2.06 ± 0.18 |
| Arg | 0.63 ± 0.31 | 0.90 ± 0.10 |
| Pro | 1.16 ± 0.36 | 2.46 ± 1.28 |
| Sample | FRS (mg GAE/g) | ABTS (mg Vit C eq./g) | FRAP (mM Fe2+ eq./g) |
|---|---|---|---|
| L-LEC | 10.7 ± 1.0 a | 11.1 ± 0.2 a | 179.1 ± 8.2 a |
| L-PPL | 9.5 ± 1.4 a | 9.7 ± 0.1 b | 143.3 ± 5.0 b |
| ChE | 42.2 ± 3.7 b | 26.1 ± 1.3 c | 922 ± 69 c |
| Tentative Identification | Retention Time (min) | [M-H] (m/z) | Molecular Formula | Score (%) | MS/MS Fragments (m/z) |
|---|---|---|---|---|---|
| Malic acid | 6.22 | 133.01 | C4H6O5 | 96 | 71, 79, 116 |
| Citric acid | 8.52 | 191.02 | C6H8O7 | 95 | 111, 87 |
| Quinic acid | 18.62 | 191.05 | C7H12O6 | 99 | 101, 114, 85 |
| Protocatechuic acid-O-hexoside | 19.18 | 315.07 | C13H16O9 | 98 | 153, 109 |
| Protocatechuic acid | 21.12 | 153.02 | C7H6O4 | 99 | 109 |
| Salicylic acid glucoside | 21.65 | 299.08 | C13H16O8 | 95 | 137, 237 |
| p-Hydroxybenzoic acid | 24.72 | 137.02 | C7H6O3 | 91 | 108, 92 |
| Caffeic acid hexoside | 25.13 | 341.09 | C15H18O9 | 90 | 135, 179 |
| Caftaric acid | 26.23 | 311.04 | C13H12O9 | 92 | 135, 179 |
| Caffeic acid | 28.20 | 179.04 | C9H8O4 | 87 | 135 |
| Salvianolic acid I/H | 30.03 | 537.11 | C27H22O12 | 94 | 269,183, 109 |
| Coutaric acid | 30.90 | 295.05 | C13H12O8 | 84 | 179, 133, 71 |
| Fertaric acid | 31.49 | 325.06 | C14H14O9 | 96 | 193, 134 |
| Rosmarinic acid 3-O-glucoside | 34.49 | 521.13 | C24H26O13 | 98 | 161, 323, 179 |
| Ferulic acid | 35.79 | 193.05 | C10H10O4 | 97 | 90 |
| Ellagic acid | 36.09 | 301.00 | C14H6O8 | 97 | 145, 228 |
| Salvianolic acid E/B/L | 36.58 | 717.15 | C36H30O16 | 89 | 519, 338, 118 |
| Rosmarinic acid | 37.87 | 359.08 | C18H16O8 | 99 | 161, 135, 73 |
| Salvianolic acid C | 41.12 | 491.10 | C26H20O10 | 88 | 311, 267, 123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alemán, A.; Pérez-García, S.; Fernández de Palencia, P.; Montero, M.P.; Gómez-Guillén, M.d.C. Physicochemical, Antioxidant, and Anti-Inflammatory Properties of Rapeseed Lecithin Liposomes Loading a Chia (Salvia hispanica L.) Seed Extract. Antioxidants 2021, 10, 693. https://doi.org/10.3390/antiox10050693
Alemán A, Pérez-García S, Fernández de Palencia P, Montero MP, Gómez-Guillén MdC. Physicochemical, Antioxidant, and Anti-Inflammatory Properties of Rapeseed Lecithin Liposomes Loading a Chia (Salvia hispanica L.) Seed Extract. Antioxidants. 2021; 10(5):693. https://doi.org/10.3390/antiox10050693
Chicago/Turabian StyleAlemán, Ailén, Selene Pérez-García, Pilar Fernández de Palencia, María Pilar Montero, and María del Carmen Gómez-Guillén. 2021. "Physicochemical, Antioxidant, and Anti-Inflammatory Properties of Rapeseed Lecithin Liposomes Loading a Chia (Salvia hispanica L.) Seed Extract" Antioxidants 10, no. 5: 693. https://doi.org/10.3390/antiox10050693
APA StyleAlemán, A., Pérez-García, S., Fernández de Palencia, P., Montero, M. P., & Gómez-Guillén, M. d. C. (2021). Physicochemical, Antioxidant, and Anti-Inflammatory Properties of Rapeseed Lecithin Liposomes Loading a Chia (Salvia hispanica L.) Seed Extract. Antioxidants, 10(5), 693. https://doi.org/10.3390/antiox10050693