
A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective


Abstract
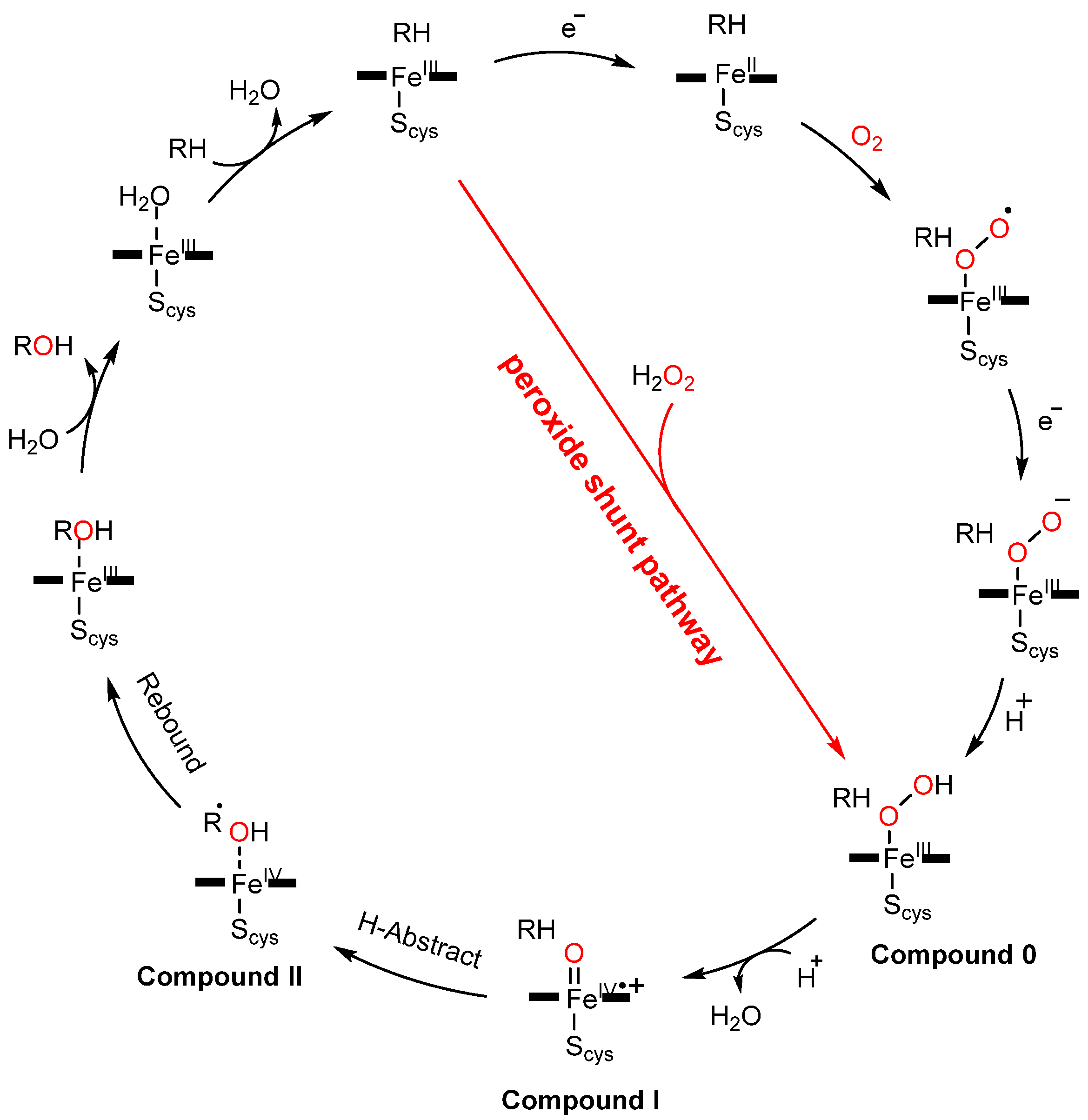
:1. Introduction
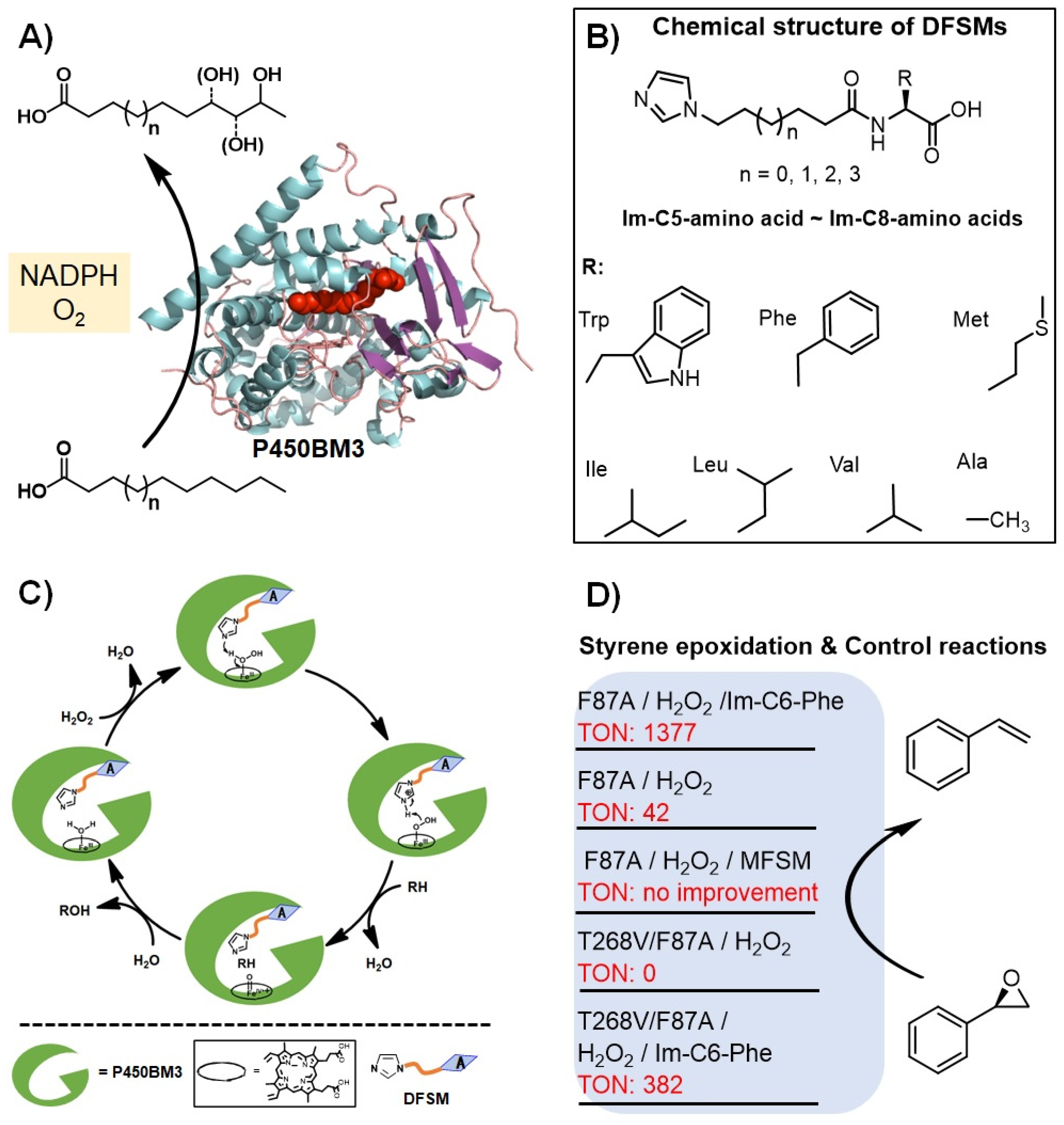
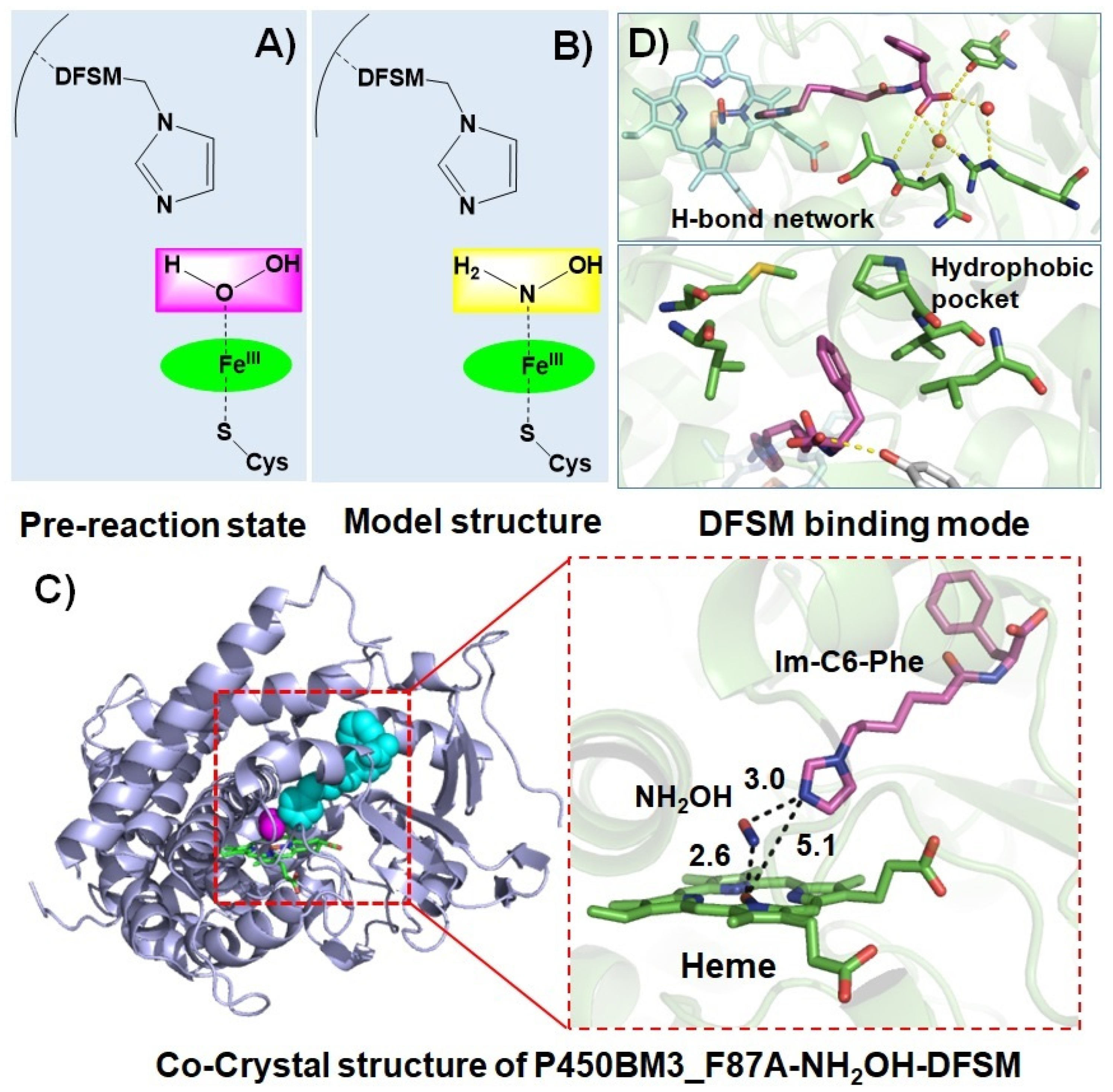
2. Proof-of-Concept of the DFSM-Facilitated P450 Peroxygenase
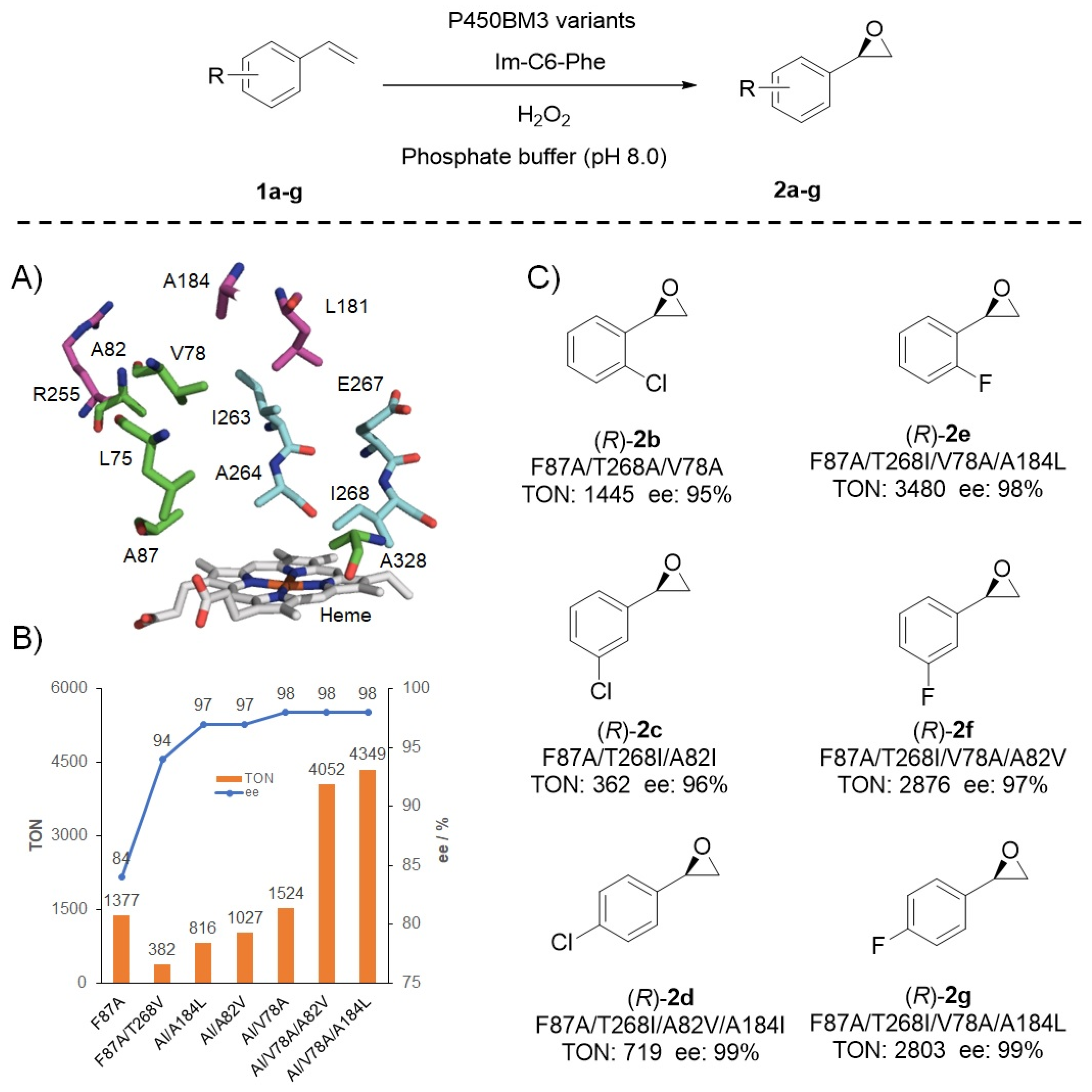
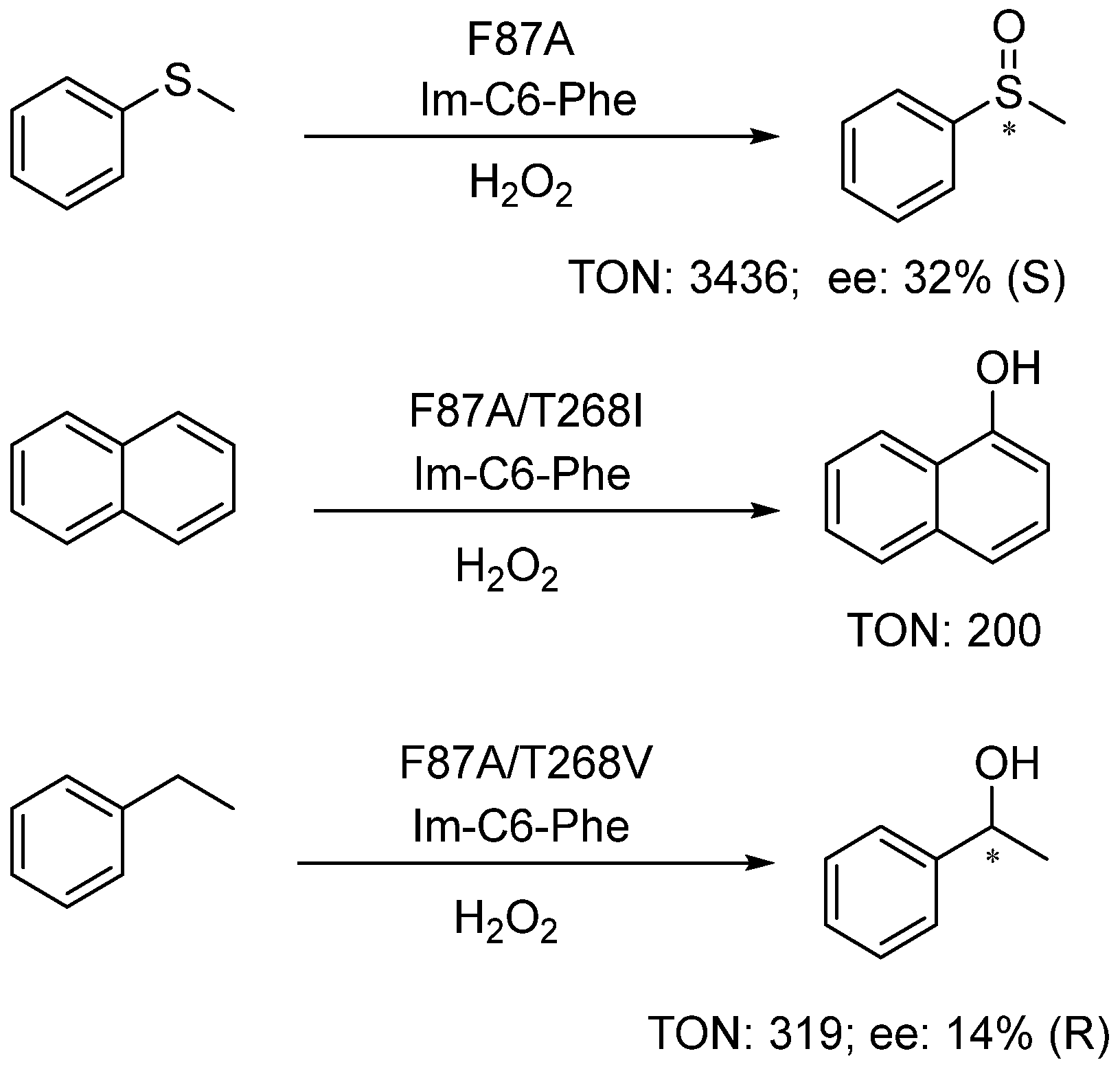
3. Catalytic Applications of the DFSM-Facilitated P450 Peroxygenase
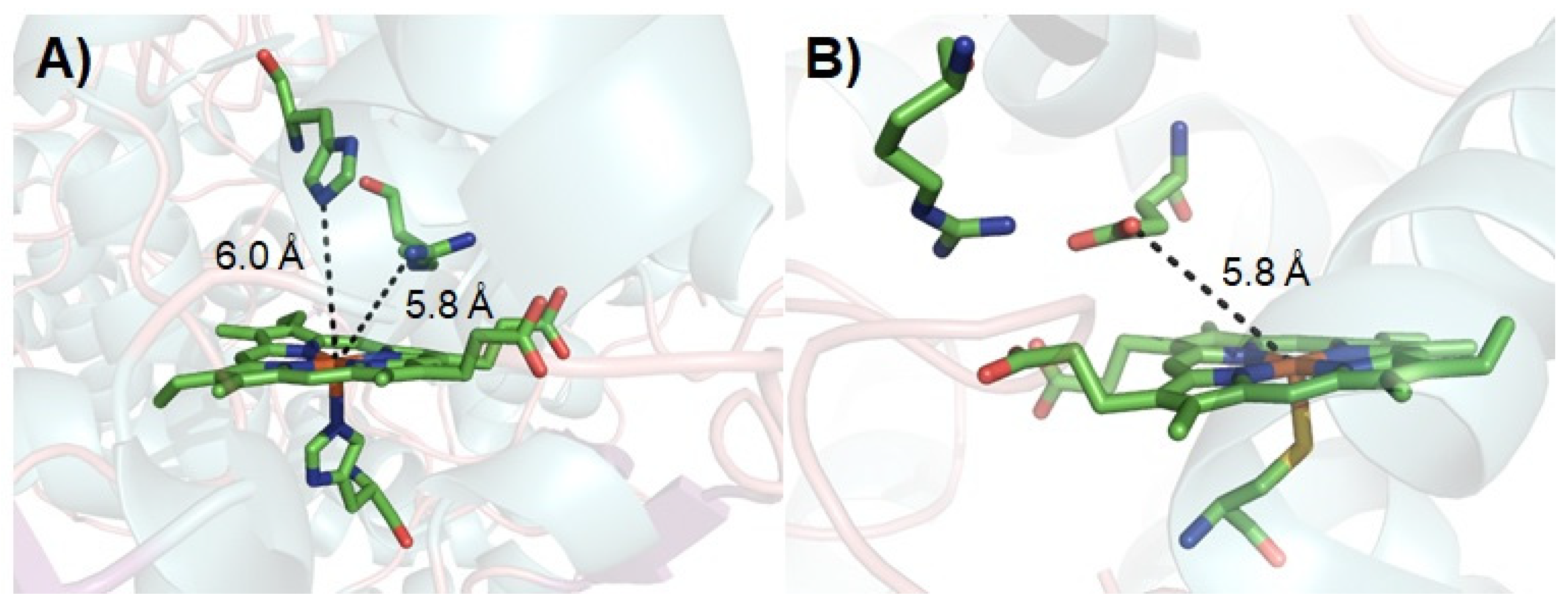
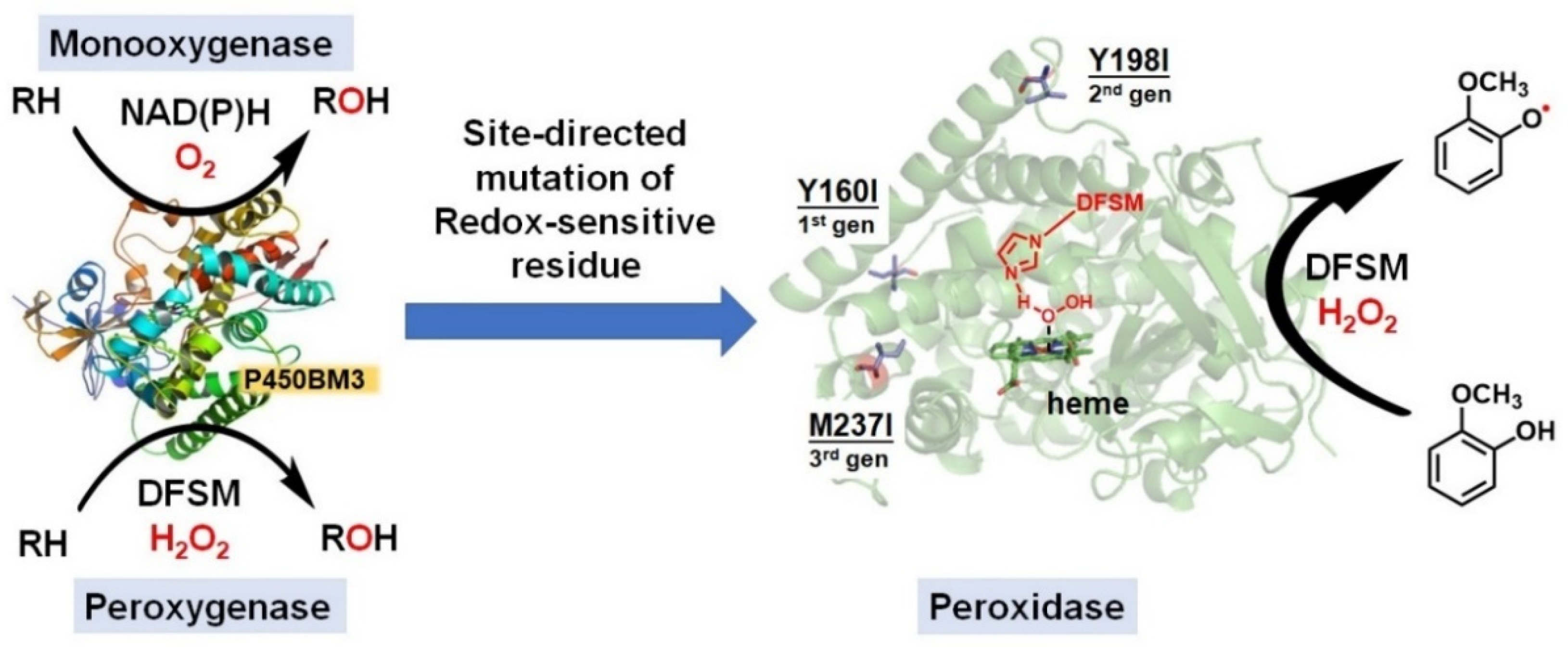
4. Switching Peroxidase Activity of the DFSM-Facilitated P450 Peroxygenase
5. Summary and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Ortiz de Montellano, P.R.O. Cytochrome P450: Structure, Mechanism, and Biochemistry, 3rd ed.; Kluwer Academic: New York, NY, USA; Plenum Publishers: New York, NY, USA, 2005. [Google Scholar]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta. Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R.; Urlacher, V.B. Cytochromes P450 as promising catalysts for biotechnological application: Chances and limitations. Appl. Microbiol. Biotech. 2014, 98, 6185–6203. [Google Scholar] [CrossRef] [PubMed]
- Fasan, R. Tuning P450 Enzymes as Oxidation Catalysts. ACS Catal. 2012, 2, 647–666. [Google Scholar] [CrossRef]
- de Montellano, P.R.O. Hydrocarbon Hydroxylation by Cytochrome P450 Enzymes. Chem. Rev. 2010, 110, 932–948. [Google Scholar] [CrossRef] [Green Version]
- Auclair, K.; Polic, V. Monoxygenase, Peroxidase and Peroxygenase Properties and Mechanisms of Cytochrome P450; Hrycay, E.G., Bandiera, S.M., Eds.; Springer: New YorK, NY, USA, 2015; pp. 209–228. [Google Scholar]
- Nastri, F.; Chino, M.; Maglio, O.; Bhagi-Damodaran, A.; Lu, Y.; Lombardi, A. Design and engineering of artificial oxygen-activating metalloenzymes. Chem. Soc. Rev. 2016, 45, 5020–5054. [Google Scholar] [CrossRef]
- Wang, T.T.; Fan, X.T.; Hou, C.X.; Liu, J.Q. Design of artificial enzymes by supramolecular strategies. Curr. Opin. Struct. Biol. 2018, 51, 19–27. [Google Scholar] [CrossRef]
- Wei, Y.F.; Ang, E.L.; Zhao, H.M. Recent developments in the application of P450 based biocatalysts. Curr. Opin. Chem. Biol. 2018, 43, 1–7. [Google Scholar] [CrossRef]
- McLntosh, J.A.; Farwell, C.C.; Arnold, F.H. Expanding P450 catalytic reaction space through evolution and engineering. Curr. Opin. Chem. Biol. 2014, 19, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Li, S.Y. Expansion of chemical space for natural products by uncommon P450 reactions. Nat. Prod. Rep. 2017, 34, 1061–1089. [Google Scholar] [CrossRef]
- Kumar, S.; Zhao, Y.; Sun, L.; Negi, S.S.; Halpert, J.R.; Muralidhara, B.K. Rational engineering of human cytochrome P450 2B6 for enhanced expression and stability: Importance of a Leu264 -> Phe substitution. Mol. Pharmacol. 2007, 72, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Rocha, C.G.; Gamble, C.G.; Lonsdale, R.; Li, A.; Nett, N.; Hoebenreich, S.; Lingnau, J.B.; Wirtz, C.; Fares, C.; Hinrichs, H.; et al. P450-Catalyzed Regio- and Diastereoselective Steroid Hydroxylation: Efficient Directed Evolution Enabled by Mutability Landscaping. ACS Catal. 2018, 8, 3395–3410. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Peng, Y.; Zhao, J.; Li, Q.; Yu, X.; Acevedo-Rocha, C.G.; Li, A. Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 2020, 7, 2. [Google Scholar] [CrossRef]
- Braunegg, G.; de Raadt, A.; Feichtenhofer, S.; Griengl, H.; Kopper, I.I.; Lehmann, A.; Weber, H.J. The Concept of Docking/Protecting Groups in Biohydroxylation. Angew. Chem. 1999, 38, 2763–2766. [Google Scholar] [CrossRef]
- de Raadt, A.; Griengl, H.; Weber, H. The concept of docking and protecting groups in biohydroxylation. Chem. Eur. J. 2001, 7, 27–31. [Google Scholar] [CrossRef]
- Munzer, D.F.; Meinhold, P.; Peters, M.W.; Feichtenhofer, S.; Griengl, H.; Arnold, F.H.; Glieder, A.; de Raadt, A. Stereoselective hydroxylation of an achiral cyclopentanecarboxylic acid derivative using engineered P450s BM-3. Chem. Commun. 2005, 20, 2597–2599. [Google Scholar] [CrossRef] [Green Version]
- Landwehr, M.; Hochrein, L.; Otey, C.R.; Kasrayan, A.; Bäckvall, J.-E.; Arnold, F.H. Enantioselective α-Hydroxylation of 2-Arylacetic Acid Derivatives and Buspirone Catalyzed by Engineered Cytochrome P450 BM-3. J. Am. Chem. Soc. 2006, 128, 6058–6059. [Google Scholar] [CrossRef] [Green Version]
- Narayan, A.R.H.; Jimenez-Oses, G.; Liu, P.; Negretti, S.; Zhao, W.X.; Gilbert, M.M.; Ramabhadran, R.O.; Yang, Y.F.; Furan, L.R.; Li, Z.; et al. Enzymatic hydroxylation of an unactivated methylene C-H bond guided by molecular dynamics simulations. Nat. Chem. 2015, 7, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Chaulagain, M.R.; Knauff, A.R.; Podust, L.M.; Montgomery, J.; Sherman, D.H. Selective oxidation of carbolide C-H bonds by an engineered macrolide P450 mono-oxygenase. Proc. Natl. Acad. Sci. USA 2009, 106, 18463–18468. [Google Scholar] [CrossRef] [Green Version]
- Negretti, S.; Narayan, A.R.H.; Chiou, K.C.; Kells, P.M.; Stachowski, J.L.; Hansen, D.A.; Podust, L.M.; Montgomery, J.; Sherman, D.H. Directing Group-Controlled Regioselectivity in an Enzymatic C-H Bond Oxygenation. J. Am. Chem. Soc. 2014, 136, 4901–4904. [Google Scholar] [CrossRef]
- Larsen, A.T.; May, E.M.; Auclair, K. Predictable Stereoselective and Chemoselective Hydroxylations and Epoxidations with P450 3A4. J. Am. Chem. Soc. 2011, 133, 7853–7858. [Google Scholar] [CrossRef]
- Polic, V.; Cheong, K.J.; Hammerer, F.; Auclair, K. Regioselective Epoxidations by Cytochrome P450 3A4 Using a Theobromine Chemical Auxiliary to Predictably Produce N-Protected β- or γ-Amino Epoxides. Adv. Synth. Catal. 2017, 359, 3983–3989. [Google Scholar] [CrossRef]
- Polic, V.; Auclair, K. Controlling substrate specificity and product regio- and stereo-selectivities of P450 enzymes without mutagenesis. Bioorg. Med. Chem. 2014, 22, 5547–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, O.; Watanabe, Y. Bringing out the Potential of Wild-type Cytochrome P450s using Decoy Molecules: Oxygenation of Nonnative Substrates by Bacterial Cytochrome P450s. Isr. J. Chem. 2015, 55, 32–39. [Google Scholar] [CrossRef]
- Shoji, O.; Watanabe, Y. Monooxygenation of Nonnative Substrates Catalyzed by Bacterial Cytochrome P450s Facilitated by Decoy Molecules. Chem. Lett. 2017, 46, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Shoji, O.; Aiba, Y.; Watanabe, Y. Hoodwinking Cytochrome P450BM3 into Hydroxylating Non-Native Substrates by Exploiting Its Substrate Misrecognition. Acc. Chem. Res. 2019, 52, 925–934. [Google Scholar] [CrossRef]
- Hara, M.; Ohkawa, H.; Narato, M.; Shirai, M.; Asada, Y.; Karube, I.; Miyake, J. Regeneration of NADPH by cactus chloroplasts: Coupling reaction with P450 monooxygenase. J. Ferment. Bioeng. 1997, 84, 324–329. [Google Scholar] [CrossRef]
- Lee, S.H.; Kwon, Y.C.; Kim, D.M.; Park, C.B. Cytochrome P450-catalyzed O-dealkylation coupled with photochemical NADPH regeneration. Biotechnol. Bioeng. 2013, 110, 383–390. [Google Scholar] [CrossRef]
- Cirino, P.C.; Arnold, F.H. Regioselectivity and activity of cytochrome P450 BM-3 and mutant F87A in reactions driven by hydrogen peroxide. Adv. Synth. Catal. 2002, 344, 932–937. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Zhou, L.; Cong, Z. Progress in cytochrome P450 monooxygenase driven by hydrogen peroxide. Biot. Resour. 2017, 39, 75–84. [Google Scholar]
- Hiroya, K.; Murakami, Y.; Shimizu, T.; Hatano, M.; Demontellano, P.R.O. Differential roles of Glu318 and Thr319 in cytochrome P450 1A2 catalysis supported by NADPH-cytochrome P450 reductase and tert-butyl hydroperoxide. Arch. Biochem. Biophys. 1994, 310, 397–401. [Google Scholar] [CrossRef]
- Cho, K.B.; Moreau, Y.; Kumar, D.; Rock, D.A.; Jones, J.P.; Shaik, S. Formation of the active species of cytochrome p450 by using iodosylbenzene: A case for spin-selective reactivity. Chem. Eur. J. 2007, 13, 4103–4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunham, N.P.; Arnold, F.H. Nature’s Machinery, Repurposed: Expanding the Repertoire of Iron-Dependent Oxygenases. ACS Catal. 2020, 10, 12239–12255. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, C.; Cong, Z. Strategies for Substrate-Regulated P450 Catalysis: From Substrate Engineering to Co-catalysis. Chem. Eur. J. 2019, 25, 6853–6863. [Google Scholar] [CrossRef]
- Shoji, O.; Watanabe, Y. Peroxygenase reactions catalyzed by cytochromes P450. J. Biol. Inorg. Chem. 2014, 19, 529–539. [Google Scholar] [CrossRef]
- Zhang, L.B.; Wang, Q. Harnessing P450 Enzyme for Biotechnology and Synthetic Biology. Chembiochem 2022, 23, e202100439. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.B.; Girhard, M. Cytochrome P450 Monooxygenases in Biotechnology and Synthetic Biology. Trends Biotechnol. 2019, 37, 882–897. [Google Scholar] [CrossRef]
- Munro, A.W.; McLean, K.J.; Grant, J.L.; Makris, T.M. Structure and function of the cytochrome P450 peroxygenase enzymes. Biochem. Soc. Trans. 2018, 46, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Bornscheuer, U.T.; Pohl, M. Improved biocatalysts by directed evolution and rational protein design. Curr. Opin. Chem. Biol. 2001, 5, 137–143. [Google Scholar] [CrossRef]
- Wang, X.D.; Saba, T.; Yiu, H.H.P.; Howe, R.F.; Anderson, J.A.; Shi, J.F. Cofactor NAD(P)H Regeneration Inspired by Heterogeneous Pathways. Chem 2017, 2, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Yamada, A.; Sugimoto, H.; Matsunaga, I.; Ogura, H.; Ichihara, K.; Adachi, S.; Park, S.Y.; Shiro, Y. Substrate recognition and molecular mechanism of fatty acid hydroxylation by cytochrome P450 from Bacillus subtilis. Crystallographic, spectroscopic, and mutational studies. J. Biol. Chem. 2003, 278, 9761–9767. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, I.; Yokotani, N.; Gotoh, O.; Kusunose, E.; Yamada, M.; Ichihara, K. Molecular Cloning and Expression of Fatty Acid α-Hydroxylase from Sphingomonas paucimobilis. J. Biol. Chem. 1997, 272, 23592–23596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujishiro, T.; Shoji, O.; Nagano, S.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Crystal structure of H2O2-dependent cytochrome P450SPalpha with its bound fatty acid substrate: Insight into the regioselective hydroxylation of fatty acids at the alpha position. J. Biol. Chem. 2011, 286, 29941–29950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rude, M.A.; Baron, T.S.; Brubaker, S.; Alibhai, M.; Del Cardayre, S.B.; Schirmer, A. Terminal olefin (1-alkene) biosynthesis by a novel p450 fatty acid decarboxylase from Jeotgalicoccus species. Appl. Environ. Microb. 2011, 77, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markel, U.; Lanvers, P.; Sauer, D.F.; Wittwer, M.; Dhoke, G.V.; Davari, M.D.; Schiffels, J.; Schwaneberg, U. A Photoclick-Based High-Throughput Screening for the Directed Evolution of Decarboxylase OleT. Chem. Eur. J. 2021, 27, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Girhard, M.; Schuster, S.; Dietrich, M.; Duerre, P.; Urlacher, V.B. Cytochrome P450 monooxygenase from Clostridium acetobutylicum: A new alpha-fatty acid hydroxylase. Biochem. Biophys. Res. Commun. 2007, 362, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Hrycay, E.G.; Gustafsson, J.A.; Ingelmansundberg, M.; Ernster, L. Sodium periodate, sodium chlorite, organic hydroperoxides, and H2O2 as hydroxylating agents in steroid hydroxylation reactions catalyzed by partially purified cytochrome P-450. Biochem. Biophys. Res. Commun. 1975, 66, 209–216. [Google Scholar] [CrossRef]
- Anari, M.R.; Josephy, P.D.; Henry, T.; Obrien, P.J. Hydrogen peroxide supports human and rat cytochrome P450 1A2-catalyzed 2-amino-3-methylimidazo 4,5-f quinoline bioactivation to mutagenic metabolites: Significance of cytochrome P450 peroxygenase. Chem. Res. Toxicol. 1997, 10, 582–588. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Li, Y.; Stearns, R.A.; de Montellano, P.R.O.; Baillie, T.A.; Tang, W. Cytochrome p450 3A4-mediated oxidative conversion of a cyano to an amide group in the metabolism of pinacidil. Biochemistry 2002, 41, 2712–2718. [Google Scholar] [CrossRef]
- Rabe, K.S.; Kiko, K.; Niemeyer, C.M. Characterization of the peroxidase activity of CYP119, a thermostable P450 from Sulfolobus acidocaldarius. Chembiochem 2008, 9, 420–425. [Google Scholar] [CrossRef]
- Goyal, S.; Banerjee, S.; Mazumdar, S. Oxygenation of monoenoic fatty acids by CYP175A1, an orphan cytochrome P450 from Thermus thermophilus HB27. Biochemistry 2012, 51, 7880–7890. [Google Scholar] [CrossRef]
- Gelb, M.H.; Heimbrook, D.C.; Malkonen, P.; Sligar, S.G. Stereochemistry and deuterium isotope effects in camphor hydroxylation by the cytochrome P450cam monoxygenase system. Biochemistry 1982, 21, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Cirino, P.C.; Arnold, F.H. A Self-Sufficient Peroxide-Driven Hydroxylation Biocatalyst. Angew. Chem. Int. Ed. 2003, 42, 3299–3301. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Chen, C.S.; Waxman, D.J.; Halpert, J.R. Directed evolution of mammalian cytochrome P4502B1. J. Biol. Chem. 2005, 280, 19569–19575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behera, R.K.; Goyal, S.; Mazumdar, S. Modification of the heme active site to increase the peroxidase activity of thermophilic cytochrome P450: A rational approach. J. Inorg. Biochem. 2010, 104, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.; Lin, Z.L.; Arnold, F.H. Laboratory evolution of peroxide-mediated cytochrome P450 hydroxylation. Nature 1999, 399, 670–673. [Google Scholar] [CrossRef]
- Gajhede, M.; Schuller, D.J.; Henriksen, A.; Smith, A.T.; Poulos, T.L. Crystal structure of horseradish peroxidase C at 2.15 angstrom resolution. Nat. Struct. Biol. 1997, 4, 1032–1038. [Google Scholar] [CrossRef]
- Piontek, K.; Strittmatter, E.; Ullrich, R.; Grobe, G.; Pecyna, M.J.; Kluge, M.; Scheibner, K.; Hofrichter, M.; Plattner, D.A. Structural basis of substrate conversion in a new aromatic peroxygenase: Cytochrome P450 functionality with benefits. J. Biol. Chem. 2013, 288, 34767–34776. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, S.I.; Roach, M.P.; Matsui, T.; Watanabe, Y. Investigations of the roles of the distal heme environment and the proximal heme iron ligand in peroxide activation by heme enzymes via molecular engineering of myoglobin. Acc. Chem. Res. 2001, 34, 818–825. [Google Scholar] [CrossRef]
- Watanabe, Y.; Ueno, T. Introduction of P450, Peroxidase, and Catalase Activities into Myoglobin by Site-Directed Mutagenesis: Diverse Reactivities of Compound I. Bull. Chem. Soc. Jpn. 2003, 76, 1309–1322. [Google Scholar] [CrossRef]
- Adachi, S.; Nagano, S.; Ishimori, K.; Watanabe, Y.; Morishima, I.; Egawa, T.; Kitagawa, T.; Makino, R. Roles of proximal ligand in heme proteins: Replacement of proximal histidine of human myoglobin with cysteine and tyrosine by site-directed mutagenesis as models for P-450, chloroperoxidase, and catalase. Biochemistry 1993, 32, 241–252. [Google Scholar] [CrossRef]
- Shoji, O.; Fujishiro, T.; Nishio, K.; Kano, Y.; Kimoto, H.; Chien, S.-C.; Onoda, H.; Muramatsu, A.; Tanaka, S.; Hori, A.; et al. A substrate-binding-state mimic of H2O2-dependent cytochrome P450 produced by one-point mutagenesis and peroxygenation of non-native substrates. J. Inorg. Biochem. 2016, 6, 5806–5811. [Google Scholar] [CrossRef] [Green Version]
- Podgorski, M.N.; Harbort, J.S.; Lee, J.H.Z.; Nguyen, G.T.H.; Bruning, J.B.; Donald, W.A.; Bernhardt, P.V.; Harmer, J.R.; Bell, S.G. An Altered Heme Environment in an Engineered Cytochrome P450 Enzyme Enables the Switch from Monooxygenase to Peroxygenase Activity. ACS Catal. 2022, 12, 1614–1625. [Google Scholar] [CrossRef]
- Whitehouse, C.J.C.; Bell, S.G.; Wong, L.-L. P450BM3 (CYP102A1): Connecting the dots. Chem. Soc. Rev. 2012, 41, 1218–1260. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Chen, Z.; Chen, J.; Chen, J.; Wang, C.; Zhou, H.; Yao, L.; Shoji, O.; Watanabe, Y.; Cong, Z. Dual-Functional Small Molecules for Generating an Efficient Cytochrome P450BM3 Peroxygenase. Angew. Chem. Int. Ed. 2018, 57, 7628–7633. [Google Scholar] [CrossRef]
- Willot, S.J.P.; Tieves, F.; Girhard, M.; Urlacher, V.B.; Hollmann, F.; de Gonzalo, G. P450BM3-Catalyzed Oxidations Employing Dual Functional Small Molecules. Catalysts 2019, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lan, D.; Durrani, R.; Hollmann, F. Peroxygenases en route to becoming dream catalysts. What are the opportunities and challenges? Curr. Opin. Chem. Biol. 2017, 37, 1–9. [Google Scholar] [CrossRef]
- Olmedo, A.; Aranda, C.; del Río, J.C.; Kiebist, J.; Scheibner, K.; Martínez, A.T.; Gutiérrez, A. From Alkanes to Carboxylic Acids: Terminal Oxygenation by a Fungal Peroxygenase. Angew. Chem. Int. Ed. 2016, 55, 12248–12251. [Google Scholar] [CrossRef]
- Imai, M.; Shimada, H.; Watanabe, Y.; Matsushima-Hibiya, Y.; Makino, R.; Koga, H.; Horiuchi, T.; Ishimura, Y. Uncoupling of the cytochrome P-450cam monooxygenase reaction by a single mutation, threonine-252 to alanine or valine: Possible role of the hydroxy amino acid in oxygen activation. Proc. Natl. Acad. Sci. USA 1989, 86, 7823–7827. [Google Scholar] [CrossRef] [Green Version]
- Martinis, S.A.; Atkins, W.M.; Stayton, P.S.; Sligar, S.G. A Conserved Residue of Cytochrome-P-450 Is Involved in Heme-Oxygen Stability and Activation. J. Am. Chem. Soc. 1989, 111, 9252–9253. [Google Scholar] [CrossRef]
- Truan, G.; Peterson, J.A. Thr268 in Substrate Binding and Catalysis in P450BM-31. Arch. Biochem. Biophy. 1998, 349, 53–64. [Google Scholar] [CrossRef]
- Rittle, J.; Green, M.T. Cytochrome P450 compound I: Capture, characterization, and C-H bond activation kinetics. Science 2010, 330, 933–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Jiang, Y.; Chen, Q.; Dong, S.; Feng, Y.; Cong, Z.; Shaik, S.; Wang, B. H-Bonding Networks Dictate the Molecular Mechanism of H2O2 Activation by P450. ACS Catal. 2021, 11, 8774–8785. [Google Scholar] [CrossRef]
- Haines, D.C.; Tomchick, D.R.; Machius, M.; Peterson, J.A. Pivotal role of water in the mechanism of P450BM-3. Biochemistry 2001, 40, 13456–13465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, Z.; Shoji, O.; Kasai, C.; Kawakami, N.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Activation of Wild-Type Cytochrome P450BM3 by the Next Generation of Decoy Molecules: Enhanced Hydroxylation of Gaseous Alkanes and Crystallographic Evidence. ACS Catal. 2014, 5, 150–156. [Google Scholar] [CrossRef]
- Shoji, O.; Yanagisawa, S.; Stanfield, J.K.; Suzuki, K.; Cong, Z.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Direct Hydroxylation of Benzene to Phenol by Cytochrome P450BM3 Triggered by Amino Acid Derivatives. Angew. Chem. Int. Ed. 2017, 56, 10324–10329. [Google Scholar] [CrossRef]
- Wang, B.; Li, C.; Dubey, K.D.; Shaik, S. Quantum mechanical/molecular mechanical calculated reactivity networks reveal how cytochrome P450cam and Its T252A mutant select their oxidation pathways. J. Am. Chem. Soc. 2015, 137, 7379–7390. [Google Scholar] [CrossRef]
- Ramirez-Escudero, M.; Molina-Espeja, P.; Gomez de Santos, P.; Hofrichter, M.; Sanz-Aparicio, J.; Alcalde, M. Structural Insights into the Substrate Promiscuity of a Laboratory-Evolved Peroxygenase. ACS Chem. Biol. 2018, 13, 3259–3268. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Micro. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef]
- Sigmund, M.-C.; Poelarends, G.J. Current state and future perspectives of engineered and artificial peroxygenases for the oxyfunctionalization of organic molecules. Nat. Catal. 2020, 3, 690–702. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R. Oxidations catalyzed by fungal peroxygenases. Curr. Opin. Chem. Biol. 2014, 19, 116–125. [Google Scholar] [CrossRef]
- Bormann, S.; Gomez Baraibar, A.; Ni, Y.; Holtmann, D.; Hollmann, F. Specific oxyfunctionalisations catalysed by peroxygenases: Opportunities, challenges and solutions. Catal. Sci. Technol. 2015, 5, 2038–2052. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, R.; Poraj-Kobielska, M.; Scholze, S.; Halbout, C.; Sandvoss, M.; Pecyna, M.J.; Scheibner, K.; Hofrichter, M. Side chain removal from corticosteroids by unspecific peroxygenase. J. Inorg. Biochem. 2018, 183, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Molina-Espeja, P.; Canellas, M.; Plou, F.J.; Hofrichter, M.; Lucas, F.; Guallar, V.; Alcalde, M. Synthesis of 1-Naphthol by a Natural Peroxygenase Engineered by Directed Evolution. ChemBioChem 2016, 17, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Gomez de Santos, P.; Cañellas, M.; Tieves, F.; Younes, S.H.H.; Molina-Espeja, P.; Hofrichter, M.; Hollmann, F.; Guallar, V.; Alcalde, M. Selective Synthesis of the Human Drug Metabolite 5′-Hydroxypropranolol by an Evolved Self-Sufficient Peroxygenase. ACS Catal. 2018, 8, 4789–4799. [Google Scholar] [CrossRef] [Green Version]
- Hagstrom, A.E.; Tornvall, U.; Nordblad, M.; Hatti-Kaul, R.; Woodley, J.M. Chemo-enzymatic epoxidation-process options for improving biocatalytic productivity. Biotechnol. Prog. 2011, 27, 67–76. [Google Scholar] [CrossRef]
- Wunschik, D.S.; Ingenbosch, K.N.; Suss, P.; Liebelt, U.; Quint, S.; Dyllick-Brenzinger, M.; Zuhse, R.; Menyes, U.; Hoffmann-Jacobsen, K.; Opwis, K.; et al. Enzymatic epoxidation of cyclohexene by peroxidase immobilization on a textile and an adapted reactor design. Enzyme. Microb. Technol. 2020, 136, 109512. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Fernandez-Fueyo, E.; Gomez Baraibar, A.; Ullrich, R.; Hofrichter, M.; Yanase, H.; Alcalde, M.; van Berkel, W.J.; Hollmann, F. Peroxygenase-Catalyzed Oxyfunctionalization Reactions Promoted by the Complete Oxidation of Methanol. Angew. Chem. Int. Ed. 2016, 55, 798–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, D.S.; Lee, H.; Tieves, F.; Lee, Y.W.; Son, E.J.; Zhang, W.; Shin, B.; Hollmann, F.; Park, C.B. Bias-Free in situ H2O2 Generation in a Photovoltaic-Photoelectrochemical Tandem Cell for Biocatalytic Oxyfunctionalization. ACS Catal. 2019, 9, 10562–10566. [Google Scholar] [CrossRef]
- Yayci, A.; Baraibar, A.G.; Krewing, M.; Fueyo, E.F.; Hollmann, F.; Alcalde, M.; Kourist, R.; Bandow, J.E. Plasma-Driven in situ Production of Hydrogen Peroxide for Biocatalysis. ChemSusChem 2020, 13, 2072–2079. [Google Scholar] [CrossRef]
- Zhao, P.; Chen, J.; Ma, N.; Chen, J.; Qin, X.; Liu, C.; Yao, F.; Yao, L.; Jin, L.; Cong, Z. Enabling highly (R)-enantioselective epoxidation of styrene by engineering unique non-natural P450 peroxygenases. Chem. Sci. 2021, 12, 6307–6314. [Google Scholar] [CrossRef]
- Chen, J.; Kong, F.; Ma, N.; Zhao, P.; Liu, C.; Wang, X.; Cong, Z. Peroxide-Driven Hydroxylation of Small Alkanes Catalyzed by an Artificial P450BM3 Peroxygenase System. ACS Catal. 2019, 9, 7350–7355. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, C.; Ma, N.; Chen, J.; Liu, C.; Wang, F.; Xu, J.; Cong, Z. Regioselective aromatic O-demethylation with an artificial P450BM3 peroxygenase system. Catal. Sci. Technol. 2020, 10, 1219–1223. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, J.; Ma, N.; Zhou, H.; Cong, Z. Selective hydroxylation of naphthalene using the H2O2-dependent engineered P450BM3 driven by dual-functional small molecules. J. Porphyr. Phthalocyanines. 2018, 22, 831–836. [Google Scholar] [CrossRef]
- Xu, G.; Mahmood, Q.; Lv, C.; Yang, R.; Zhou, L.; Wang, Q. Asymmetric kinetic resolution polymerization. Coord. Chem. Rev. 2020, 414, 213296. [Google Scholar] [CrossRef]
- Ren, W.-M.; Yue, T.-J.; Zhang, X.; Gu, G.-G.; Liu, Y.; Lu, X.-B. Stereoregular CO2 Copolymers from Epoxides with an Electron-Withdrawing Group: Crystallization and Unexpected Stereocomplexation. Macromolecules 2017, 50, 7062–7069. [Google Scholar] [CrossRef]
- Wu, G.-P.; Wei, S.-H.; Ren, W.-M.; Lu, X.-B.; Li, B.; Zu, Y.-P.; Darensbourg, D.J. Alternating copolymerization of CO2 and styrene oxide with Co(iii)-based catalyst systems: Differences between styrene oxide and propylene oxide. Energy Environ. Sci. 2011, 4, 5084–5092. [Google Scholar] [CrossRef]
- Wu, S.; Zhou, Y.; Li, Z. Biocatalytic selective functionalisation of alkenes via single-step and one-pot multi-step reactions. Chem. Commun. 2019, 55, 883–896. [Google Scholar] [CrossRef]
- Matsumoto, K.; Oguma, T.; Katsuki, T. Highly enantioselective epoxidation of styrenes catalyzed by proline-derived C1-symmetric titanium(salan) complexes. Angew. Chem. Int. Ed. 2009, 48, 7432–7435. [Google Scholar] [CrossRef]
- Wang, C.; Yamamoto, H. Asymmetric Epoxidation Using Hydrogen Peroxide as Oxidant. Chem.–Asian J. 2015, 10, 2056–2068. [Google Scholar] [CrossRef]
- Lin, H.; Xu, M.-Y.; Liu, Y.; Wu, Z.-L. Biocatalytic Epoxidation for Green Synthesis. In Green Biocatalysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 351–372. [Google Scholar]
- Wang, L.; Wei, S.; Pan, X.; Liu, P.; Du, X.; Zhang, C.; Pu, L.; Wang, Q. Enhanced Turnover for the P450 119 Peroxygenase-Catalyzed Asymmetric Epoxidation of Styrenes by Random Mutagenesis. Chem. Eur. J. 2018, 24, 2741–2749. [Google Scholar] [CrossRef]
- Li, A.; Liu, J.; Pham, S.Q.; Li, Z. Engineered P450pyr monooxygenase for asymmetric epoxidation of alkenes with unique and high enantioselectivity. Chem. Commun. 2013, 49, 11572–11574. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Wang, J.B.; Reetz, M.T. Exploiting Designed Oxidase-Peroxygenase Mutual Benefit System for Asymmetric Cascade Reactions. J. Am. Chem. Soc. 2019, 141, 5655–5658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, T.; Scholtissek, A.; Hofmann, S.; Koch, R.; Tischler, D. Accessing Enantiopure Epoxides and Sulfoxides: Related Flavin-Dependent Monooxygenases Provide Reversed Enantioselectivity. ChemCatChem 2020, 12, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Tang, Y.; Dong, S.; Lang, R.; Chen, H. A new monooxygenase from Herbaspirillum huttiense catalyzed highly enantioselective epoxidation of allylbenzenes and allylic alcohols. Catal. Sci. Technol. 2020, 10, 2145–2151. [Google Scholar] [CrossRef]
- van Schie, M.; Paul, C.E.; Arends, I.; Hollmann, F. Photoenzymatic epoxidation of styrenes. Chem. Commun. 2019, 55, 1790–1792. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.; Guo, C.; Lin, H.; Ding, Z.Y.; Wu, Z.L. Functional characterization of an (R)-selective styrene monooxygenase from streptomyces sp. NRRL S-31. Enzym. Microb. Technol. 2019, 132, 109391. [Google Scholar] [CrossRef]
- Xiao, H.; Dong, S.; Liu, Y.; Pei, X.-Q.; Lin, H.; Wu, Z.-L. A new clade of styrene monooxygenases for (R)-selective epoxidation. Catal. Sci. Technol. 2021, 11, 2195–2201. [Google Scholar] [CrossRef]
- Nesterov, D.S.; Nesterova, O.V.; Pombeiro, A.J.L. Homo- and heterometallic polynuclear transition metal catalysts for alkane CH bonds oxidative functionalization: Recent advances. Coord. Chem. Rev. 2018, 355, 199–222. [Google Scholar] [CrossRef]
- Soussan, L.; Pen, N.; Belleville, M.-P.; Marcano, J.S.; Paolucci-Jeanjean, D. Alkane biohydroxylation: Interests, constraints and future developments. J. Biotechnol. 2016, 222, 117–142. [Google Scholar] [CrossRef]
- Banerj Ee, R.; Proshlyakov, Y.; Lipscomb, J.D.; Proshlyakov, D.A. Structure of the key species in the enzymatic oxidation of methane to methanol. Nature 2015, 518, 431. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.R. Handbook of Bond Dissociation Energies in Organic Compounds; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Cooley, R.B.; Dubbel, B.L.; Sayavedra-Soto, L.A.; Bottomley, P.J.; Arp, D.J. Kinetic characterization of the soluble butane monooxygenase from Thauera butanivorans, formerly ‘Pseudomonas butanovora’. Microbiology 2009, 155, 2086–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, S.; Kinne, M.; Wang, X.S.; Ullrich, R.; Kayser, G.; Groves, J.T.; Hofrichter, M. Selective hydroxylation of alkanes by an extracellular fungal peroxygenase. FEBS J. 2011, 278, 3667–3675. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, N.; Shoji, O.; Watanabe, Y. Use of Perfluorocarboxylic Acids to Trick Cytochrome P450BM3 into Initiating the Hydroxylation of Gaseous Alkanes. Angew. Chem. Int. Ed. 2011, 50, 5315–5318. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, K.; Ariyasu, S.; Stanfield, J.K.; Suzuki, K.; Onoda, H.; Kasai, C.; Sugimoto, H.; Aiba, Y.; Watanabe, Y.; Shoji, O. Systematic Evolution of Decoy Molecules for the Highly Efficient Hydroxylation of Benzene and Small Alkanes Catalyzed by Wild-Type Cytochrome P450BM3. ACS Catal. 2020, 10, 9136–9144. [Google Scholar] [CrossRef]
- Ariyasu, S.; Kodama, Y.; Kasai, C.; Cong, Z.; Stanfield, J.K.; Aiba, Y.; Watanabe, Y.; Shoji, O. Development of a High-Pressure Reactor Based on Liquid-Flow Pressurisation to Facilitate Enzymatic Hydroxylation of Gaseous Alkanes. ChemCatChem 2019, 11, 4709–4714. [Google Scholar] [CrossRef]
- Zilly, F.E.; Acevedo, J.P.; Augustyniak, W.; Deege, A.; Haeusig, U.W.; Reetz, M.T. Tuning a P450 Enzyme for Methane Oxidation. Angew. Chem. Int. Ed. 2011, 50, 2720–2724. [Google Scholar] [CrossRef]
- Fasan, R.; Chen, M.M.; Crook, N.C.; Arnold, F.H. Engineered alkane-hydroxylating cytochrome P450(BM3) exhibiting nativelike catalytic properties. Angew. Chem. Int. Ed. 2007, 46, 8414–8418. [Google Scholar] [CrossRef]
- Patel, R.N.; Hou, C.T.; Laskin, A.I.; Felix, A. Microbial Oxidation of Hydrocarbons: Properties of a Soluble Methane Monooxygenase from a Facultative Methane-Utilizing Organism, Methylobacterium sp. Strain CRL-26. Appl. Environ. Microbiol. 1982, 44, 1130–1137. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Bell, S.G.; Lednik, J.; Insley, A.; Rao, Z.H.; Wong, L.L. The heme monooxygenase cytochrome P450(cam) can be engineered to oxidize ethane to ethanol. Angew. Chem. Int. Ed. 2005, 44, 4029–4032. [Google Scholar] [CrossRef]
- Trippe, K.M.; Wolpert, T.J.; Hyman, M.R.; Ciuffetti, L.M. RNAi silencing of a cytochrome P450 monoxygenase disrupts the ability of a filamentous fungus, Graphium sp., to grow on short-chain gaseous alkanes and ethers. Biodegradation 2014, 25, 137–151. [Google Scholar] [CrossRef]
- Gazi, S. Valorization of wood biomass-lignin via selective bond scission: A minireview. Appl. Catal. B Environ. 2019, 257, 117936. [Google Scholar] [CrossRef]
- Kamimura, N.; Takahashi, K.; Mori, K.; Araki, T.; Fujita, M.; Higuchi, Y.; Masai, E. Bacterial catabolism of lignin-derived aromatics: New findings in a recent decade: Update on bacterial lignin catabolism. Environ. Microbiol. Rep. 2017, 9, 679–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.C.; Paice, M.; Zhang, X. Enzymatic Oxidation of Lignin: Challenges and Barriers toward Practical Applications. ChemCatChem 2020, 12, 401–425. [Google Scholar] [CrossRef]
- Venkatesagowda, B. Enzymatic demethylation of lignin for potential biobased polymer applications. Fungal Biol. Rev. 2019, 33, 190–224. [Google Scholar] [CrossRef]
- Richter, N.; Zepeck, F.; Kroutil, W. Cobalamin-dependent enzymatic O-, N-, and S-demethylation. Trends Biotechnol. 2015, 33, 371–373. [Google Scholar] [CrossRef]
- Lanfranchi, E.; Trajkovic, M.; Barta, K.; de Vries, J.G.; Janssen, D.B. Exploring the Selective Demethylation of Aryl Methyl Ethers with a Pseudomonas Rieske Monooxygenase. ChemBioChem 2019, 20, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Mallinson, S.J.B.; Machovina, M.M.; Silveira, R.L.; Garcia-Borras, M.; Gallup, N.; Johnson, C.W.; Allen, M.D.; Skaf, M.S.; Crowley, M.F.; Neidle, E.L.; et al. A promiscuous cytochrome P450 aromatic O-demethylase for lignin bioconversion. Nat. Commun. 2018, 9, 2487. [Google Scholar] [CrossRef] [Green Version]
- Machovina, M.M.; Mallinson, S.J.B.; Knott, B.C.; Meyers, A.W.; Garcia-Borras, M.; Bu, L.; Gado, J.E.; Oliver, A.; Schmidt, G.P.; Hinchen, D.J.; et al. Enabling microbial syringol conversion through structure-guided protein engineering. Proc. Natl. Acad. Sci. USA 2019, 16, 13970–13976. [Google Scholar] [CrossRef] [Green Version]
- Ellis, E.S.; Hinchen, D.J.; Bleem, A.; Bu, L.; Mallinson, S.J.B.; Allen, M.D.; Streit, B.R.; Machovina, M.M.; Doolin, Q.V.; Michener, W.E.; et al. Engineering a Cytochrome P450 for Demethylation of Lignin-Derived Aromatic Aldehydes. JACS Au 2021, 1, 252–261. [Google Scholar] [CrossRef]
- Dardas, A.; Gal, D.; Barrelle, M.; Sauretignazi, G.; Sterjiades, R.; Pelmont, J. The demethylation of guaiacol by a new bacterial cytochrome P-450. Arch. Biochem. Biophys. 1985, 236, 585–592. [Google Scholar] [CrossRef]
- Eltis, L.D.; Karlson, U.; Timmis, K.N. Purification and characterization of cytochrome P450RR1 from Rhodococcus rhodochrous. Eur. J. Biochem. 1993, 213, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.G.; Zhou, R.; Yang, W.; Tan, A.B.H.; Gentleman, A.S.; Wong, L.-L.; Zhou, W. Investigation of the Substrate Range of CYP199A4: Modification of the Partition between Hydroxylation and Desaturation Activities by Substrate and Protein Engineering. Chem. Eur. J. 2012, 18, 16677–16688. [Google Scholar] [CrossRef] [PubMed]
- Leveson-Gower, R.B.; Mayer, C.; Roelfes, G. The importance of catalytic promiscuity for enzyme design and evolution. Nat. Rev. Chem. 2019, 3, 687–705. [Google Scholar] [CrossRef]
- Sandoval, B.A.; Hyster, T.K. Emerging strategies for expanding the toolbox of enzymes in biocatalysis. Curr. Opin. Chem. Biol. 2020, 55, 45–51. [Google Scholar] [CrossRef]
- Bornscheuer, U.T.; Kazlauskas, R.J. Catalytic promiscuity in biocatalysis: Using old enzymes to form new bonds and follow new pathways. Angew. Chem. Int. Ed. 2004, 43, 6032–6040. [Google Scholar] [CrossRef]
- Hrycay, E.G.; Bandiera, S.M. The monooxygenase, peroxidase, and peroxygenase properties of cytochrome P450. Arch. Biochem. Biophys. 2012, 522, 71–89. [Google Scholar] [CrossRef]
- Depillis, G.D.; Sishta, B.P.; Mauk, A.G.; Demontellano, P.R.O. Small substrates and cytochrome c are oxidized at different sites of cytochrome c peroxidase. J. Biol. Chem. 1991, 266, 19334–19341. [Google Scholar] [CrossRef]
- Zhang, P.; Yuan, H.; Xu, J.K.; Wang, X.J.; Gao, S.Q.; Tan, X.S.; Lin, Y.W. A Catalytic Binding Site Together with a Distal Tyr in Myoglobin Affords Catalytic Efficiencies Similar to Natural Peroxidases. ACS Catal. 2020, 10, 891–896. [Google Scholar] [CrossRef]
- Yamada, M.; Hashimoto, Y.; Kumano, T.; Tsujimura, S.; Kobayashi, M. New function of aldoxime dehydratase: Redox catalysis and the formation of an expected product. PLoS ONE 2017, 12, e0175846. [Google Scholar] [CrossRef] [Green Version]
- Chien, S.C.; Shoji, O.; Morimoto, Y.; Watanabe, Y. Use of apomyoglobin to gently remove heme from a H2O2-dependent cytochrome P450 and allow its reconstitution. New J. Chem. 2017, 41, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.N.; Fang, W.H.; Liu, C.A.F.; Qin, X.Q.; Wang, X.L.; Jin, L.Y.; Wang, B.J.; Cong, Z.Q. Switching an Artificial P450 Peroxygenase into Peroxidase via Mechanism-Guided Protein Engineering. ACS Catal. 2021, 11, 8449–8455. [Google Scholar] [CrossRef]
- Xu, J.; Cen, Y.X.; Singh, W.; Fan, J.J.; Wu, L.; Lin, X.F.; Zhou, J.H.; Huang, M.L.; Reetz, M.T.; Wu, Q. Stereodivergent Protein Engineering of a Lipase To Access All Possible Stereoisomers of Chiral Esters with Two Stereocenters. J. Am. Chem. Soc. 2019, 141, 7934–7945. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Wu, Q.; Reetz, M.T. Focused rational iterative site-specific mutagenesis (FRISM). In Enzyme Engineering and Evolution: General Methods; Methods in Enzymology; Tawfik, D.S., Ed.; Academic Press: New York, NY, USA, 2020; Volume 643, pp. 225–242. [Google Scholar]
- Vidal-Limon, A.; Aguila, S.; Ayala, M.; Batista, C.V.; Vazquez-Duhalt, R. Peroxidase activity stabilization of cytochrome P450BM3 by rational analysis of intramolecular electron transfer. J. Inorg. Biochem. 2013, 122, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Mate, M.J.; Romero, A.; Martinez, M.J.; Martinez, A.T.; Ruiz-Duenas, F.J. Two oxidation sites for low redox potential substrates: A directed mutagenesis, kinetic, and crystallographic study on Pleurotus eryngii versatile peroxidase. J. Biol. Chem. 2012, 287, 41053–410675. [Google Scholar] [CrossRef] [Green Version]
- Mandelman, D.; Jamal, J.; Poulos, T.L. Identification of two electron-transfer sites in ascorbate peroxidase using chemical modification, enzyme kinetics, and crystallography. Biochemistry 1998, 37, 17610–17617. [Google Scholar] [CrossRef]
- Murphy, E.J.; Metcalfe, C.L.; Nnamchi, C.; Moody, P.C.E.; Raven, E.L. Crystal structure of guaiacol and phenol bound to a heme peroxidase. FEBS J. 2012, 279, 1632–1639. [Google Scholar] [CrossRef]
- Li, Q.S.; Ogawa, J.; Shimizu, S. Critical role of the residue size at position 87 in H2O2-dependent substrate hydroxylation activity and H2O2 inactivation of cytochrome P450BM-3. Biochem. Biophys. Res. Commun. 2001, 280, 1258–1261. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Makris, T.M. Expanding the substrate scope and reactivity of cytochrome P450 OleT. Biochem. Biophys. Res. Commun. 2016, 476, 462–466. [Google Scholar] [CrossRef]
- Roberts, A.G.; Katayama, J.; Kaspera, R.; Ledwitch, K.V.; Trong, I.L.; Stenkamp, R.E.; Thompson, J.A.; Totah, R.A. The role of cytochrome P450 BM3 phenylalanine-87 and threonine-268 in binding organic hydroperoxides. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2016, 1860, 669–677. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Enzyme | Alkanes | Final Product | PFR a | TTN b | Ref. |
|---|---|---|---|---|---|---|
| 1 | BM3_F87A/T268I/A184I/Im-C6-Phe | Propane | 2-Propanol | 630 | 1775 | [93] |
| 2 | BM3_F87A/T268I/A82T/Im-C6-Phe | n-Butane | 2-Butanol | 1042 | 2253 | [93] |
| 3 | BM3/PFC10 | Propane | 2-Propanol | 70 | 700 | [117] |
| 4 | BM3/PFC9-L-Leu | Propane | 2-Propanol | 256 | 2560 | [76] |
| 5 | BM3/3CCPA-Pip-Phe | Propane | 2-Propanol | 615 | - | [118] |
| 6 | BM3/PFC9 | n-Butane | 2-Butanol | 110 | 1100 | [120] |
| 7 | BM3/PFC11 | Propane | 2-Propanol | - | 1021 | [120] |
| 8 | BM3/PFC7 | n-Butane | 2-Butanol | - | 3632 | [120] |
| 9 | BM3/C7AM-Pip-Phe | Ethane | Ethanol | 82.7 | - | [118] |
| 10 | P450cam_EB | n-Butane | 2-Butanol | 520 | - | [123] |
| 11 | P450cam_EB_L294M/T185M/L1358P/G248A | Propane | 2-Propanol | 505 | - | [123] |
| 12 | P450cam_EB_L294M/T185M/L1358P/G248A | Ethane | Ethanol | 78.2 | - | [123] |
| 13 | P450PMOR1 | Propane | 2-Propanol | 455 | 35,600 | [121] |
| 14 | P450PMOR2 | Propane | 2-Propanol | 370 | 45,800 | [121] |
| 15 c | CYP52L1 | Propane | 1-Propanol | - | - | [124] |
| 16 | sMMO | Methane | Methanol | 78 | - | [122] |
| 17 | sMMO | Ethane | Ethanol | 45.6 | - | [122] |
| 18 | sMMO | Propane | 2-Propanol | 33–58.8 | - | [122] |
| 19 | sMMO | n-Butane | 2-Butanol | 7.2–28.8 | - | [122] |
| 20 | AaeUPO | Propane | 2-Propanol | 17 | 999 | [116] |
| 21 | AaeUPO | n-Butane | 2-Butanol | 21 | 1258 | [116] |
 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Substrate | Enzyme System | Product | TON | |||||
| No. | R1 | R2 | R3 | No. | R2 | R3 | ||
| 3a | Me | H | H | BM3_F87A/T268I/Im-C5-Phe | 4a | H | H | 486 |
| 3b | Me | Me | H | BM3_F87A/T268I/Im-C6-Phe | 4b | Me | H | 356 |
| 3c | Me | OH | H | BM3_F87A/Im-C6-Phe | 4c | OH | H | 539 |
| 3d | Et | OH | H | BM3_F87G/T268G/Im-C5-Phe | 4d | OH | H | 99 |
| 3e | Me | OMe | H | BM3_F87A/T268I/Im-C6-Phe | 4e | OMe | H | 287 |
| 3f | H | OMe | OMe | BM3_F87G/T268V/Im-C5-Phe | 4f | OH | OMe | 165 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di, S.; Fan, S.; Jiang, F.; Cong, Z. A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective. Antioxidants 2022, 11, 529. https://doi.org/10.3390/antiox11030529
Di S, Fan S, Jiang F, Cong Z. A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective. Antioxidants. 2022; 11(3):529. https://doi.org/10.3390/antiox11030529
Chicago/Turabian StyleDi, Siyu, Shengxian Fan, Fengjie Jiang, and Zhiqi Cong. 2022. "A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective" Antioxidants 11, no. 3: 529. https://doi.org/10.3390/antiox11030529
APA StyleDi, S., Fan, S., Jiang, F., & Cong, Z. (2022). A Unique P450 Peroxygenase System Facilitated by a Dual-Functional Small Molecule: Concept, Application, and Perspective. Antioxidants, 11(3), 529. https://doi.org/10.3390/antiox11030529