
Profiling Distinctive Inflammatory and Redox Responses to Hydrogen Sulfide in Stretched and Stimulated Lung Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Cell Stretch Experiments
2.3. Co-Culture Experiments
2.4. Cytokine Measurements
2.5. Detection of Reactive Oxygen Species
2.6. Detection of Glutathione
2.7. Detection of Mitochondrial Activity
2.8. Actin Fluorescence Staining
2.9. Neutrophil Transmigration Assay
2.10. Neutrophil Chemotaxis Assay
2.11. Statistical Analysis
3. Results
3.1. Effects of Cyclic Stretch and Hydrogen Sulfide on Lung Epithelial Cells
3.2. Effects of Cyclic Stretch and Hydrogen Sulfide on Inflammatory Signaling between Epithelial Cells and Macrophages
3.3. Effects of Cyclic Stretch and Hydrogen Sulfide on Macrophages
3.4. Effects of Cyclic Stretch and NADPH Oxidase Inhibition on Reactive Oxygen Species and Inflammation on Macrophages
3.5. Effects of Cyclic Stretch and Hydrogen Sulfide on Inflammation and Reactive Oxygen Species in Endothelial Cells
3.6. Effects of Cyclic Stretch and Hydrogen Sulfide on Endothelial Cell Integrity
3.7. Effects of Hydrogen Sulfide and ROS Scavenging on Neutrophil Transmigration through Endothelial Monolayers
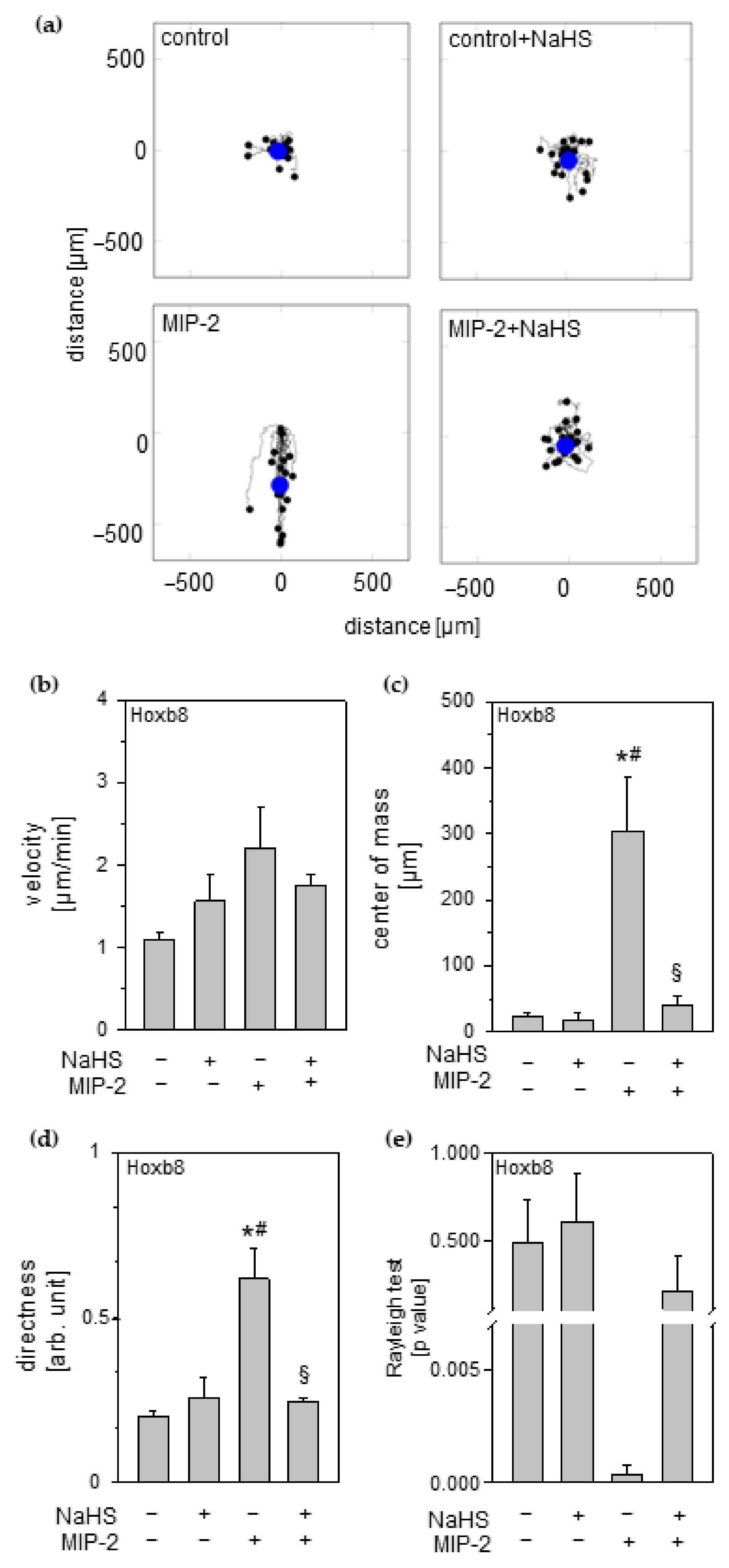
3.8. Effect of Hydrogen Sulfide on Neutrophil Chemotaxis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neto, A.S.; Simonis, F.D.; Barbas, C.S.; Biehl, M.; Determann, R.M.; Elmer, J.; Friedman, G.; Gajic, O.; Goldstein, J.N.; Linko, R.; et al. Lung-Protective Ventilation With Low Tidal Volumes and the Occurrence of Pulmonary Complications in Patients Without Acute Respiratory Distress Syndrome: A Systematic Review and Individual Patient Data Analysis. Crit. Care Med. 2015, 10, 2155–2163. [Google Scholar] [CrossRef] [PubMed]
- Serpa, N.A.; Hemmes, S.N.; Barbas, C.S.; Beiderlinden, M.; Biehl, M.; Binnekade, J.M.; Canet, J.; Fernandez-Bustamante, A.; Futier, E.; Gajic, O.; et al. Protective versus Conventional Ventilation for Surgery: A Systematic Review and Individual Patient Data Meta-analysis. Anesthesiology 2015, 1, 66–78. [Google Scholar] [CrossRef] [PubMed]
- The Acute Respiratory Distress Syndrome Network. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. N. Engl. J. Med. 2000, 18, 1301–1308. [Google Scholar]
- McGuinness, G.; Zhan, C.; Rosenberg, N.; Azour, L.; Wickstrom, M.; Mason, D.M.; Thomas, K.M.; Moore, W.H. Increased Incidence of Barotrauma in Patients with COVID-19 on Invasive Mechanical Ventilation. Radiology 2020, 2, E252–E262. [Google Scholar] [CrossRef]
- Davidovich, N.; DiPaolo, B.C.; Lawrence, G.G.; Chhour, P.; Yehya, N.; Margulies, S.S. Cyclic stretch-induced oxidative stress increases pulmonary alveolar epithelial permeability. Am. J. Respir. Cell Mol. Biol. 2013, 1, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Lionetti, V.; Recchia, F.A.; Ranieri, V.M. Overview of ventilator-induced lung injury mechanisms. Curr. Opin. Crit Care 2005, 1, 82–86. [Google Scholar] [CrossRef]
- Vlahakis, N.E.; Hubmayr, R.D. Cellular stress failure in ventilator-injured lungs. Am. J. Respir. Crit Care Med. 2005, 12, 1328–1342. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Kita, M.; Kimura, I.; Oseko, F.; Terauchi, R.; Takahashi, K.; Kubo, T.; Kanamura, N. Mechanical stress induces expression of cytokines in human periodontal ligament cells. Oral Dis. 2006, 2, 171–175. [Google Scholar] [CrossRef]
- Oudin, S.; Pugin, J. Role of MAP kinase activation in interleukin-8 production by human BEAS-2B bronchial epithelial cells submitted to cyclic stretch. Am. J. Respir. Cell Mol. Biol. 2002, 1, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Spassov, S.G.; Faller, S.; Hummel, M.; Helo, K.; Ihle, A.; Ryter, S.W.; Strosing, K.M.; Hoetzel, A. Hydrogen Sulfide Confers Lung Protection During Mechanical Ventilation via Cyclooxygenase 2, 15-deoxy Delta12,14-Prostaglandin J2, and Peroxisome Proliferator-Activated Receptor Gamma. Crit Care Med. 2017, 8, e849–e857. [Google Scholar] [CrossRef]
- Spassov, S.G.; Kessler, C.; Jost, R.; Schumann, S. Ventilation-Like Mechanical Strain Modulates the Inflammatory Response of BEAS2B Epithelial Cells. Oxid. Med. Cell Longev. 2019, 2019, 2769761. [Google Scholar] [CrossRef] [Green Version]
- Imanaka, H.; Shimaoka, M.; Matsuura, N.; Nishimura, M.; Ohta, N.; Kiyono, H. Ventilator-induced lung injury is associated with neutrophil infiltration, macrophage activation, and TGF-beta 1 mRNA upregulation in rat lungs. Anesth. Analg. 2001, 2, 428–436. [Google Scholar]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 3, 293–307. [Google Scholar] [CrossRef]
- Faller, S.; Ryter, S.W.; Choi, A.M.; Loop, T.; Schmidt, R.; Hoetzel, A. Inhaled hydrogen sulfide protects against ventilator-induced lung injury. Anesthesiology 2010, 1, 104–115. [Google Scholar] [CrossRef] [Green Version]
- Aslami, H.; Heinen, A.; Roelofs, J.J.; Zuurbier, C.J.; Schultz, M.J.; Juffermans, N.P. Suspended animation inducer hydrogen sulfide is protective in an in vivo model of ventilator-induced lung injury. Intensive Care Med. 2010, 11, 1946–1952. [Google Scholar] [CrossRef] [Green Version]
- Francis, R.C.; Vaporidi, K.; Bloch, K.D.; Ichinose, F.; Zapol, W.M. Protective and Detrimental Effects of Sodium Sulfide and Hydrogen Sulfide in Murine Ventilator-induced Lung Injury. Anesthesiology 2011, 5, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Spassov, S.; Pfeifer, D.; Strosing, K.; Ryter, S.; Hummel, M.; Faller, S.; Hoetzel, A. Genetic targets of hydrogen sulfide in ventilator-induced lung injury--a microarray study. PLoS ONE 2014, 7, e102401. [Google Scholar] [CrossRef]
- Schmidt, E.P.; Lee, W.L.; Zemans, R.L.; Yamashita, C.; Downey, G.P. On, around, and through: Neutrophil-endothelial interactions in innate immunity. Physiology 2011, 5, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Hattori, H.; Subramanian, K.K.; Sakai, J.; Jia, Y.; Li, Y.; Porter, T.F.; Loison, F.; Sarraj, B.; Kasorn, A.; Jo, H.; et al. Small-molecule screen identifies reactive oxygen species as key regulators of neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2010, 8, 3546–3551. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, M.; Li, L.; Rose, P.; Tan, C.H.; Parkinson, D.B.; Moore, P.K. The effect of hydrogen sulfide donors on lipopolysaccharide-induced formation of inflammatory mediators in macrophages. Antioxid. Redox. Signal. 2010, 10, 1147–1154. [Google Scholar] [CrossRef]
- Pozzi, G.; Masselli, E.; Gobbi, G.; Mirandola, P.; Taborda-Barata, L.; Ampollini, L.; Carbognani, P.; Micheloni, C.; Corazza, F.; Galli, D.; et al. Hydrogen Sulfide Inhibits TMPRSS2 in Human Airway Epithelial Cells: Implications for SARS-CoV-2 Infection. Biomedicines 2021, 9, 1273. [Google Scholar] [CrossRef]
- Faller, S.; Spassov, S.G.; Zimmermann, K.K.; Ryter, S.W.; Buerkle, H.; Loop, T.; Schmidt, R.; Strosing, K.M.; Hoetzel, A. Hydrogen sulfide prevents hyperoxia-induced lung injury by downregulating reactive oxygen species formation and angiopoietin-2 release. Curr. Pharm. Des. 2013, 15, 2715–2721. [Google Scholar] [CrossRef]
- Spassov, S.G.; Donus, R.; Ihle, P.M.; Engelstaedter, H.; Hoetzel, A.; Faller, S. Hydrogen Sulfide Prevents Formation of Reactive Oxygen Species through PI3K/Akt Signaling and Limits Ventilator-Induced Lung Injury. Oxid. Med.Cell Longev. 2017, 2017, 3715037. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 7, 671–675. [Google Scholar] [CrossRef]
- Pepperell, E.E.; Watt, S.M. A novel application for a 3-dimensional timelapse assay that distinguishes chemotactic from chemokinetic responses of hematopoietic CD133(+) stem/progenitor cells. Stem Cell Res. 2013, 2, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Pendyala, S.; Usatyuk, P.V.; Gorshkova, I.A.; Garcia, J.G.; Natarajan, V. Regulation of NADPH oxidase in vascular endothelium: The role of phospholipases, protein kinases, and cytoskeletal proteins. Antioxid. Redox. Signal. 2009, 4, 841–860. [Google Scholar] [CrossRef] [Green Version]
- Jufri, N.F.; Mohamedali, A.; Avolio, A.; Baker, M.S. Mechanical stretch: Physiological and pathological implications for human vascular endothelial cells. Vasc. Cell. 2015, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Stroka, K.M.; Aranda-Espinoza, H. Endothelial cell substrate stiffness influences neutrophil transmigration via myosin light chain kinase-dependent cell contraction. Blood 2011, 6, 1632–1640. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Li, H.; Gong, Y.; Zheng, H.; Zhao, D. Hydrogen sulfide ameliorated lipopolysaccharide-induced acute lung injury by inhibiting autophagy through PI3K/Akt/mTOR pathway in mice. Biochem. Biophys. Res. Commun. 2018, 1, 514–518. [Google Scholar] [CrossRef]
- Pugin, J.; Dunn, I.; Jolliet, P.; Tassaux, D.; Magnenat, J.L.; Nicod, L.P.; Chevrolet, J.C. Activation of human macrophages by mechanical ventilation in vitro. Am. J. Physiol. 1998, 1 Pt 6, L1040–L1050. [Google Scholar] [CrossRef]
- Herold, S.; Mayer, K.; Lohmeyer, J. Acute lung injury: How macrophages orchestrate resolution of inflammation and tissue repair. Front. Immunol. 2011, 2, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peteranderl, C.; Morales-Nebreda, L.; Selvakumar, B.; Lecuona, E.; Vadász, I.; Morty, R.E.; Schmoldt, C.; Bespalowa, J.; Wolff, T.; Pleschka, S.; et al. Macrophage-epithelial paracrine crosstalk inhibits lung edema clearance during influenza infection. J. Clin. Investig. 2016, 4, 1566–1580. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Jiang, Q.; Yang, S.; Huang, S.; Han, X.; Duan, J.; Pan, S.; Zhao, M.; Guo, S. GYY4137 attenuates LPS-induced acute lung injury via heme oxygenase-1 modulation. Pulm. Pharmacol. Ther. 2019, 54, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Rios, E.C.; Soriano, F.G.; Olah, G.; Gero, D.; Szczesny, B.; Szabo, C. Hydrogen sulfide modulates chromatin remodeling and inflammatory mediator production in response to endotoxin, but does not play a role in the development of endotoxin tolerance. J. Inflamm. 2016, 13, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.L.; Ren, J.D.; Huang, Z.; Wang, T.; Xiang, K.; Cheng, L.; Tang, L.J. The Role of Exogenous Hydrogen Sulfide in Free Fatty Acids Induced Inflammation in Macrophages. Cell Physiol. Biochem. 2017, 4, 1635–1644. [Google Scholar] [CrossRef] [PubMed]
- Muschter, D.; Beiderbeck, A.S.; Spath, T.; Kirschneck, C.; Schroder, A.; Grassel, S. Sensory Neuropeptides and their Receptors Participate in Mechano-Regulation of Murine Macrophages. Int. J. Mol. Sci. 2019, 3, 503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelblanco, M.; Lugrin, J.; Ehirchiou, D.; Nasi, S.; Ishii, I.; So, A.; Martinon, F.; Busso, N. Hydrogen sulfide inhibits NLRP3 inflammasome activation and reduces cytokine production both in vitro and in a mouse model of inflammation. J. Biol. Chem. 2018, 7, 2546–2557. [Google Scholar] [CrossRef] [Green Version]
- Naik, E.; Dixit, V.M. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. J. Exp. Med. 2011, 3, 417–420. [Google Scholar] [CrossRef]
- Carnesecchi, S.; Pache, J.C.; Barazzone-Argiroffo, C. NOX enzymes: Potential target for the treatment of acute lung injury. Cell Mol. Life Sci. 2012, 14, 2373–2385. [Google Scholar] [CrossRef]
- Chapman, K.E.; Sinclair, S.E.; Zhuang, D.; Hassid, A.; Desai, L.P.; Waters, C.M. Cyclic mechanical strain increases reactive oxygen species production in pulmonary epithelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 5, L834–L841. [Google Scholar] [CrossRef]
- Katsuyama, M. NOX/NADPH oxidase, the superoxide-generating enzyme: Its transcriptional regulation and physiological roles. J. Pharmacol. Sci. 2010, 2, 134–146. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Goto, Y.; Kimura, H. Hydrogen sulfide increases glutathione production and suppresses oxidative stress in mitochondria. Antioxid. Redox. Signal. 2010, 1, 1–13. [Google Scholar] [CrossRef]
- Boshtam, M.; Kouhpayeh, S.; Amini, F.; Azizi, Y.; Najaflu, M.; Shariati, L.; Khanahmad, H. Anti-inflammatory effects of apocynin: A narrative review of the evidence. All Life 2021, 1, 997–1010. [Google Scholar] [CrossRef]
- Kim, S.Y.; Moon, K.A.; Jo, H.Y.; Jeong, S.; Seon, S.H.; Jung, E.; Cho, Y.S.; Chun, E.; Lee, K.Y. Anti-inflammatory effects of apocynin, an inhibitor of NADPH oxidase, in airway inflammation. Immunol. Cell Biol. 2012, 4, 441–448. [Google Scholar] [CrossRef]
- Iwaki, M.; Ito, S.; Morioka, M.; Iwata, S.; Numaguchi, Y.; Ishii, M.; Kondo, M.; Kume, H.; Naruse, K.; Sokabe, M.; et al. Mechanical stretch enhances IL-8 production in pulmonary microvascular endothelial cells. Biochem. Biophys. Res. Commun. 2009, 3, 531–536. [Google Scholar] [CrossRef]
- Li, H.D.; Zhang, Z.R.; Zhang, Q.X.; Qin, Z.C.; He, D.M.; Chen, J.S. Treatment with exogenous hydrogen sulfide attenuates hyperoxia-induced acute lung injury in mice. Eur. J. Appl. Physiol. 2013, 6, 1555–1563. [Google Scholar] [CrossRef]
- Zimmermann, K.K.; Spassov, S.G.; Strosing, K.M.; Ihle, P.M.; Engelstaedter, H.; Hoetzel, A.; Faller, S. Hydrogen Sulfide Exerts Anti-oxidative and Anti-inflammatory Effects in Acute Lung Injury. Inflammation 2018, 1, 249–259. [Google Scholar] [CrossRef]
- Tian, D.; Dong, J.; Jin, S.; Teng, X.; Wu, Y. Endogenous hydrogen sulfide-mediated MAPK inhibition preserves endothelial function through TXNIP signaling. Free Radic. Biol. Med. 2017, 110, 291–299. [Google Scholar] [CrossRef]
- Tian, Y.; Gawlak, G.; O’Donnell, J.J., III; Mambetsariev, I.; Birukova, A.A. Modulation of Endothelial Inflammation by Low and High Magnitude Cyclic Stretch. PLoS ONE 2016, 4, e0153387. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Teng, X.; Jin, S.; Dong, J.; Guo, Q.; Tian, D.; Wu, Y. Hydrogen sulfide improves endothelial dysfunction by inhibiting the vicious cycle of NLRP3 inflammasome and oxidative stress in spontaneously hypertensive rats. J. Hypertens. 2019, 8, 1633–1643. [Google Scholar] [CrossRef]
- Breton-Romero, R.; Acin-Perez, R.; Rodriguez-Pascual, F.; Martinez-Molledo, M.; Brandes, R.P.; Rial, E.; Enriquez, J.A.; Lamas, S. Laminar shear stress regulates mitochondrial dynamics, bioenergetics responses and PRX3 activation in endothelial cells. Biochim. Biophys. Acta 2014, 11, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shi, S.; Dong, S.; Wu, J.; Song, M.; Zhong, X.; Liu, Y. Sodium hydrosulfide attenuates hyperhomocysteinemia rat myocardial injury through cardiac mitochondrial protection. Mol. Cell Biochem. 2015, 1, 189–200. [Google Scholar] [CrossRef]
- Anwar, M.A.; Shalhoub, J.; Lim, C.S.; Gohel, M.S.; Davies, A.H. The effect of pressure-induced mechanical stretch on vascular wall differential gene expression. J. Vasc. Res. 2012, 6, 463–478. [Google Scholar] [CrossRef]
- Zhang, L.J.; Tao, B.B.; Wang, M.J.; Jin, H.M.; Zhu, Y.C. PI3K p110alpha isoform-dependent Rho GTPase Rac1 activation mediates H2S-promoted endothelial cell migration via actin cytoskeleton reorganization. PLoS ONE 2012, 9, e44590. [Google Scholar]
- Faller, S.; Hausler, F.; Goeft, A.; von Itter, M.A.; Gyllenram, V.; Hoetzel, A.; Spassov, S.G. Hydrogen sulfide limits neutrophil transmigration, inflammation, and oxidative burst in lipopolysaccharide-induced acute lung injury. Sci. Rep. 2018, 1, 14676. [Google Scholar] [CrossRef] [Green Version]
- Matthay, M.A.; Zemans, R.L. The acute respiratory distress syndrome: Pathogenesis and treatment. Annu. Rev. Pathol. 2011, 6, 147–163. [Google Scholar] [CrossRef] [Green Version]
- Dal-Secco, D.; Cunha, T.M.; Freitas, A.; Alves-Filho, J.C.; Souto, F.O.; Fukada, S.Y.; Grespan, R.; Alencar, N.M.; Neto, A.F.; Rossi, M.A.; et al. Hydrogen sulfide augments neutrophil migration through enhancement of adhesion molecule expression and prevention of CXCR2 internalization: Role of ATP-sensitive potassium channels. J. Immunol. 2008, 6, 4287–4298. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.L.; Liu, X.H.; Gong, Q.H.; Wu, D.; Zhu, Y.Z. Hydrogen sulfide attenuated tumor necrosis factor-alpha-induced inflammatory signaling and dysfunction in vascular endothelial cells. PLoS ONE 2011, 5, e19766. [Google Scholar]
- Giustarini, G.; Vrisekoop, N.; Kruijssen, L.; Wagenaar, L.; van Staveren, S.; van Roest, M.; Bleumink, R.; Bol-Schoenmakers, M.; Weaver, R.J.; Koenderman, L.; et al. Trovafloxacin-Induced Liver Injury: Lack in Regulation of Inflammation by Inhibition of Nucleotide Release and Neutrophil Movement. Toxicol. Sci. 2019, 2, 385–396. [Google Scholar] [CrossRef]
- Redecke, V.; Wu, R.; Zhou, J.; Finkelstein, D.; Chaturvedi, V.; High, A.A.; Hacker, H. Hematopoietic progenitor cell lines with myeloid and lymphoid potential. Nat. Methods 2013, 8, 795–803. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Plate Coating | Medium | H2S | Elongation | ||||
|---|---|---|---|---|---|---|---|---|
| Volume | Supplements | Source | Conc. | Shape | [%] | [h] | ||
| A549 | Collagen IV | 1 mL | - | NaHS | 1 mM | Square | 20.5 | 4 |
| BEAS-2B | Collagen I | 1 mL | + | NaHS | 1 mM | Square | 20.5 | 6 |
| RAW264.7 | Collagen I | 1 mL | - | NaHS | 2 mM | ½ Sinus | 20.5 | 6 |
| HUVEC | Collagen IV | 2 mL | + | GYY | 1 mM | Sinus | 20.0 | 24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spassov, S.G.; Faller, S.; Goeft, A.; Von Itter, M.-N.A.; Birkigt, A.; Meyerhoefer, P.; Ihle, A.; Seiler, R.; Schumann, S.; Hoetzel, A. Profiling Distinctive Inflammatory and Redox Responses to Hydrogen Sulfide in Stretched and Stimulated Lung Cells. Antioxidants 2022, 11, 1001. https://doi.org/10.3390/antiox11051001
Spassov SG, Faller S, Goeft A, Von Itter M-NA, Birkigt A, Meyerhoefer P, Ihle A, Seiler R, Schumann S, Hoetzel A. Profiling Distinctive Inflammatory and Redox Responses to Hydrogen Sulfide in Stretched and Stimulated Lung Cells. Antioxidants. 2022; 11(5):1001. https://doi.org/10.3390/antiox11051001
Chicago/Turabian StyleSpassov, Sashko G., Simone Faller, Andreas Goeft, Marc-Nicolas A. Von Itter, Andreas Birkigt, Peter Meyerhoefer, Andreas Ihle, Raphael Seiler, Stefan Schumann, and Alexander Hoetzel. 2022. "Profiling Distinctive Inflammatory and Redox Responses to Hydrogen Sulfide in Stretched and Stimulated Lung Cells" Antioxidants 11, no. 5: 1001. https://doi.org/10.3390/antiox11051001
APA StyleSpassov, S. G., Faller, S., Goeft, A., Von Itter, M. -N. A., Birkigt, A., Meyerhoefer, P., Ihle, A., Seiler, R., Schumann, S., & Hoetzel, A. (2022). Profiling Distinctive Inflammatory and Redox Responses to Hydrogen Sulfide in Stretched and Stimulated Lung Cells. Antioxidants, 11(5), 1001. https://doi.org/10.3390/antiox11051001