
Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Procurement and Processing of Macroalgae
2.3. Estimation of the Chlorophyll Content
2.4. Antimicrobial Activity
Percentage Inhibition of Macroalgae Extracts
2.5. Analysis of Phytochemical Content and Antioxidant Activity
2.6. Bioaccessibility of Macroalgae Phenols and Polyphenols
2.7. Cytotoxicity of Macroalgae Extracts by an MTT Assay
2.7.1. Preparation of Mouse Lymphocytes
2.7.2. Determination of Cytotoxicity
2.8. Liquid Chromatography–Mass Spectrometry LC-MS/MS Analysis
2.9. Statistical Analyses
3. Results
3.1. Chlorophyll Content
3.2. Antimicrobial Activity
3.3. Phytochemical Content and Antioxidant Activity
3.3.1. Phytochemical Analysis
3.3.2. Antioxidant Activity
3.4. Bioaccessibility of Macroalgae Polyphenols
3.5. Cytotoxicity Assay
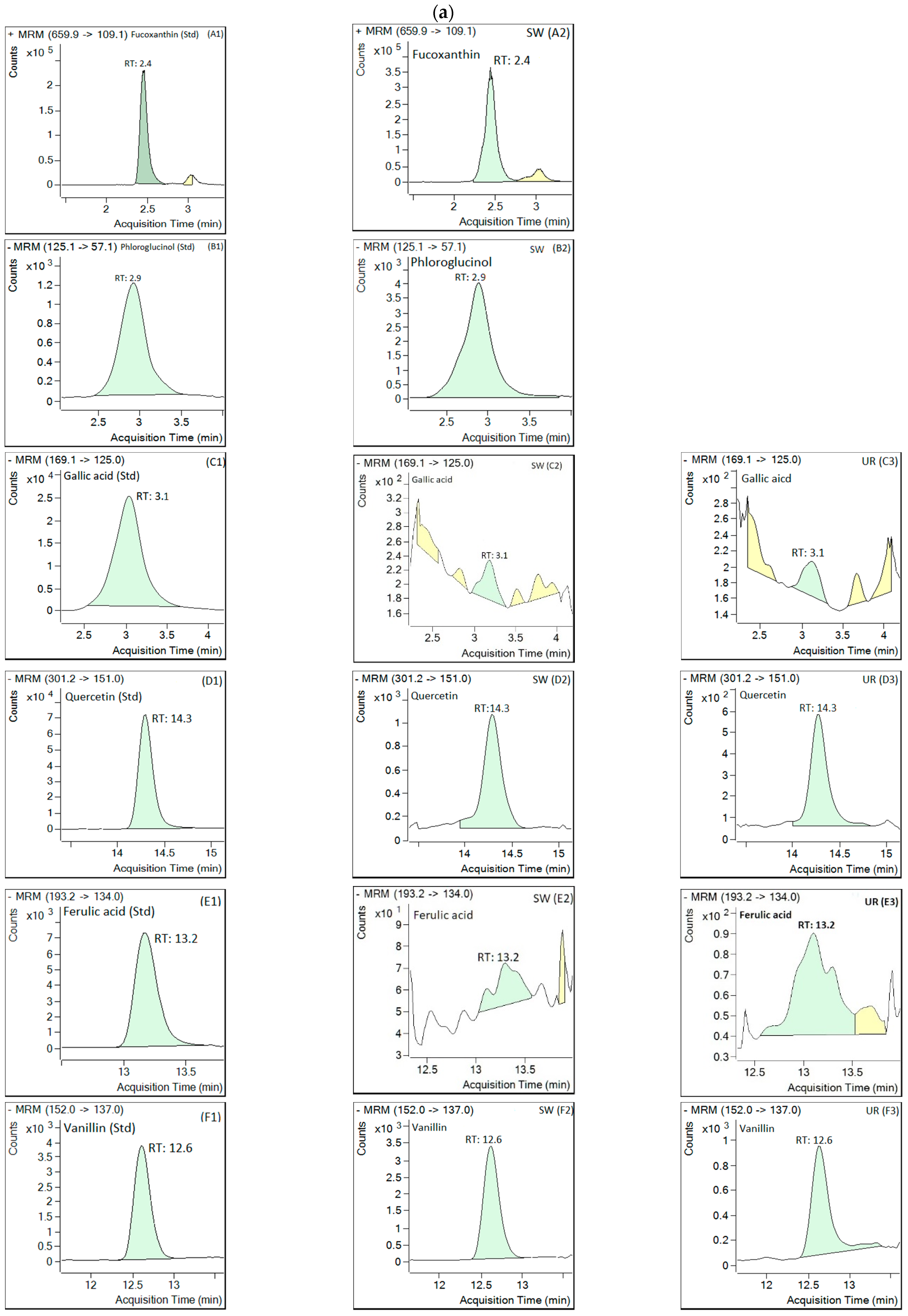
3.6. LC-MS/MS Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.; Sahoo, D.; Levine, I. Assessment of nutritional value in a brown macroalgae Sargassum wightii and their seasonal variations. Algal Res. 2015, 9, 117–125. [Google Scholar] [CrossRef]
- Santos, F.; Monteiro, J.P.; Duarte, D.; Melo, T.; Lopes, D.; da Costa, E.; Domingues, M.R. Unraveling the lipidome and antioxidant activity of native Bifurcaria bifurcata and invasive Sargassum muticum seaweeds: A lipid perspective on how systemic intrusion may present an opportunity. Antioxidants 2020, 9, 642. [Google Scholar] [CrossRef] [PubMed]
- Bansemir, A.; Blume, M.; Schröder, S.; Lindequist, U. Screening of cultivated macroalgaes for antibacterial activity against fish pathogenic bacteria. Aquaculture 2006, 252, 79–84. [Google Scholar] [CrossRef]
- Corsetto, P.A.; Montorfano, G.; Zava, S.; Colombo, I.; Ingadottir, B.; Jonsdottir, R.; Sveinsdottir, K.; Rizzo, A.M. Characterization of antioxidant potential of seaweed extracts for enrichment of convenience food. Antioxidants 2020, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Kumar, Y.; Tarafdar, A.; Badgujar, P.C. Macroalgae as a source of natural antioxidants: Therapeutic activity and food applications. J. Food Qual. 2021, 2021, 5753391. [Google Scholar] [CrossRef]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish macroalgaes. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Gomez-Zavaglia, A.; Prieto Lage, M.A.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The potential of seaweeds as a source of functional ingredients of prebiotic and antioxidant value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef] [Green Version]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Identification and characterization of phenolic antioxidant compounds from brown Irish macroalgae Himanthalia elongata using LC-DAD–ESI-MS/MS. Innov. Food Sci. Emerg. Technol. 2016, 37, 261–268. [Google Scholar] [CrossRef]
- Rajivgandhi, G.N.; Kanisha, C.C.; Ramachandran, G.; Manoharan, N.; Mothana, R.A.; Siddiqui, N.A.; Al-Rehaily, A.J.; Ullah, R.; Almarfadi, O.M. Phytochemical screening and anti-oxidant activity of Sargassum wightii enhances the anti-bacterial activity against Pseudomonas aeruginosa. Saudi J. Biol. Sci. 2021, 28, 1763–1769. [Google Scholar] [CrossRef]
- Guner, A. In vitro risk assessment of Padina pavonica (Linnaeus) (Brown algae). Food Health 2020, 7, 31–38. [Google Scholar] [CrossRef]
- Herath, K.H.I.N.M.; Cho, J.; Kim, A.; Kim, H.S.; Han, E.J.; Kim, H.J.; Kim, M.S.; Ahn, G.; Jeon, Y.J.; Jee, Y. Differential modulation of immune response and cytokine profiles of Sargassum horneri ethanol extract in murine spleen with or without Concanavalin A stimulation. Biomed. Pharmacother. 2019, 110, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, Q.; Hu, K.; Zhang, R.; Yuan, Y.; He, S.; Zeng, Q.; Su, D. Effects of in vitro simulated digestion on the free and bound phenolic content and antioxidant activity of seven species of macroalgaes. Int. J. Food Sci. 2020, 56, 2365–2374. [Google Scholar] [CrossRef]
- Kim, J.; Choi, J.H.; Ko, G.; Jo, H.; Oh, T.; Ahn, B.; Unno, T. Anti-inflammatory properties and gut microbiota modulation of Porphyra tenera extracts in dextran sodium sulfate-induced colitis in mice. Antioxidants 2020, 9, 988. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Jaiswal, A.K.; Abu-Gannam, N.; Gupta, S. Antimicrobial, antioxidant and free radical-scavenging capacity of brown macroalgae Himanthalia elongata from western coast of Ireland. J. Food Biochem. 2013, 37, 322–335. [Google Scholar] [CrossRef]
- Airanthi, M.W.A.; Hosokawa, M.; Miyashita, K. Comparative antioxidant activity of edible Japanese brown macroalgaes. J. Food Sci. 2011, 76, C104–C111. [Google Scholar] [CrossRef]
- Yildiz, G.; Celikler, S.; Vatan, O.; Dere, S. Determination of the anti-oxidative capacity and bioactive compounds in green macroalgae Ulva rigida C. Agardh. Int. J. Food Prop. 2012, 15, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Syad, A.N.; Shunmugiah, K.P.; Kasi, P.D. Macroalgaes as nutritional supplements: Analysis of nutritional profile, physicochemical properties and proximate composition of G. acerosa and S. wightii. Biomed. Prev. Nutr. 2013, 3, 139–144. [Google Scholar] [CrossRef]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, É. Assessment of the antioxidant and antibacterial activities of three species of edible macroalgaes. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Wang, T.; Jonsdottir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic macroalgaes. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Kumar, Y.; Tarafdar, A.; Kumar, D.; Badgujar, P.C. Effect of Indian brown macroalgae Sargassum wightii as a functional ingredient on the phytochemical content and antioxidant activity of coffee beverage. J. Food Sci. Technol. 2019, 56, 4516–4525. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Mishra, A.; Tarafdar, A.; Kumar, Y.; Verma, K.; Aluko, R.; Trajkovska, B.; Badgujar, P.C. In vitro bioaccessibility and characterisation of spent hen meat hydrolysate powder prepared by spray and freeze-drying techniques. Process. Biochem. 2021, 105, 128–136. [Google Scholar] [CrossRef]
- Badgujar, P.C.; Jain, S.K.; Singh, A.; Punia, J.S.; Gupta, R.P.; Chandratre, G.A. Immunotoxic effects of imidacloprid following 28 days of oral exposure in BALB/c mice. Environ. Toxicol. Pharmacol. 2013, 35, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Gandomi, H.; Derakhshandeh, A.; Misaghi, A.; Noori, N. Phytochemical composition and in vitro safety evaluation of Ziziphora clinopodioides Lam. ethanolic extract: Cytotoxicity, genotoxicity and mutagenicity assessment. J. Ethnopharmacol. 2021, 266, 113428. [Google Scholar] [CrossRef]
- Agregan, R.; Munekata, P.E.; Franco, D.; Dominguez, R.; Carballo, J.; Lorenzo, J.M. Phenolic compounds from three brown macroalgae species using LC-DAD–ESI-MS/MS. Food Res. Int. 2017, 99, 979–985. [Google Scholar] [CrossRef]
- Kumar, Y.; Singhal, S.; Tarafdar, A.; Pharande, A.; Ganesan, M.; Badgujar, P.C. Ultrasound assisted extraction of selected edible macroalgae: Effect on antioxidant activity and quantitative assessment of polyphenols by liquid chromatography with tandem mass spectrometry (LC-MS/MS). Algal Res. 2020, 52, 102114. [Google Scholar] [CrossRef]
- Sakthivel, R.; Devi, K.P. Evaluation of physicochemical properties, proximate and nutritional composition of Gracilaria edulis collected from Palk Bay. Food Chem. 2015, 174, 68–74. [Google Scholar] [CrossRef]
- Dere, Ş.; Güneş, T.; Sivaci, R. Spectrophotometric determination of chlorophyll-A, B and total carotenoid contents of some algae species using different solvents. Turk. J. Bot. 1998, 22, 13–18. [Google Scholar]
- Garcia-Perez, P.; Lourenço-Lopes, C.; Silva, A.; Pereira, A.G.; Fraga-Corral, M.; Zhao, C.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A. Pigment Composition of Nine Brown Algae from the Iberian Northwestern Coastline: Influence of the Extraction Solvent. Mar. Drugs. 2022, 20, 113. [Google Scholar] [CrossRef]
- Suganya, S.; Ishwarya, R.; Jayakumar, R.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-anbr, A.N.; Vaseeharan, B. New insecticides and antimicrobials derived from Sargassum wightii and Halimeda gracillis macroalgaes: Toxicity against mosquito vectors and antibiofilm activity against microbial pathogens. S. Afr. J. Bot. 2019, 125, 466–480. [Google Scholar] [CrossRef]
- El Fayoumy, R.A.; Pendleton, P.; El-Fallal, A.A.; Abou-Dobara, M.I.; El-Sayed, A.K. Allyl isothiocyanate release from edible Laminaria japonica for time-dependent growth deactivation of foodborne pathogens: I: Micrococcus luteus, Bacillus subtilis, and Listeria monocytogenes. Food Bioproc. Tech. 2017, 10, 1562–1573. [Google Scholar] [CrossRef]
- Kadam, S.U.; Pankaj, S.K.; Tiwari, B.K.; Cullen, P.J.; O’Donnell, C.P. Development of biopolymer-based gelatin and casein films incorporating brown macroalgae Ascophyllum nodosum extract. Food Packag. Shelf Life 2015, 6, 68–74. [Google Scholar] [CrossRef]
- Cabral, E.M.; Oliveira, M.; Mondala, J.R.; Curtin, J.; Tiwari, B.K.; Garcia-Vaquero, M. Antimicrobials from macroalgaes for food applications. Mar. Drugs 2021, 19, 211. [Google Scholar] [CrossRef] [PubMed]
- Stévant, P.; Ólafsdóttir, A.; Déléris, P.; Dumay, J.; Fleurence, J.; Ingadóttir, B.; Jonsdottir, R.; Ragueneau, E.; Rebours, C.; Rustad, T. Semi-dry storage as a maturation process for improving the sensory characteristics of the edible red seaweed dulse (Palmaria palmata). Algal Res. 2020, 51, 102048. [Google Scholar] [CrossRef]
- Dang, T.T.; Bowyer, M.C.; van Altena, I.A.; Scarlett, C.J. Comparison of chemical profile and antioxidant properties of the brown algae. Int. J. Food Sci. Technol. 2018, 53, 174–181. [Google Scholar] [CrossRef]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K.; Ming, C.H. Antioxidant activities and phenolics content of eight species of macroalgaes from north Borneo. J. Appl. Phycol. 2008, 20, 367. [Google Scholar] [CrossRef]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.-H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug. Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Lee, H.S.; Kang, I.J.; Won, M.H.; You, S. Antioxidant properties of extract and fractions from Enteromorpha prolifera, a type of green macroalgae. Food Chem. 2011, 127, 999–1006. [Google Scholar] [CrossRef]
- Francisco, J.; Horta, A.; Pedrosa, R.; Afonso, C.; Cardoso, C.; Bandarra, N.M.; Gil, M.M. Bioaccessibility of antioxidants and fatty acids from Fucus spiralis. Foods 2020, 9, 440. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, F.B.; Cermeño, M.; Kleekayai, T.; Harnedy, P.A.; FitzGerald, R.J.; Alves, R.C.; Oliveira, M.B.P.P. Effect of in vitro simulated gastrointestinal digestion on the antioxidant activity of the red macroalgae Porphyra dioica. Food Res. Int. 2020, 136, 109309. [Google Scholar] [CrossRef]
- Su, D.; Li, N.; Chen, M.; Yuan, Y.; He, S.; Wang, Y.; Wu, Q.; Li, L.; Yang, H.; Zeng, Q. Effects of in vitro digestion on the composition of flavonoids and antioxidant activities of the lotus leaf at different growth stages. Int. J. Food Sci. 2018, 53, 1631–1639. [Google Scholar] [CrossRef]
- Cassani, L.; Gomez-Zavaglia, A.; Jimenez-Lopez, C.; Lourenço-Lopes, C.; Prieto, M.A.; Simal-Gandara, J. Macroalgae-based natural ingredients: Stability of phlorotannins during extraction, storage, passage through the gastrointestinal tract and potential incorporation into functional foods. Food Res. Int. 2020, 137, 109676. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Badgujar, P.C.; Mishra, V.; Sehrawat, R.; Babar, O.A.; Upadhyay, A. Effect of microfluidization on cholesterol, thermal properties and in vitro and in vivo protein digestibility of milk. LWT-Food Sci. Technol. 2019, 116, 108523. [Google Scholar] [CrossRef]
- Rushdi, M.I.; Abdel-Rahman, I.A.; Saber, H.; Attia, E.Z.; Abdelraheem, W.M.; Madkour, H.A.; Hassan, H.M.; Elmaidomy, A.H.; Abdelmohsen, U.R. Pharmacological and natural products diversity of the brown algae genus Sargassum. RSC Adv. 2020, 10, 24951–24972. [Google Scholar] [CrossRef]
- Rodeiro, I.; Hernández, I.; Herrera, J.A.; Riera, M.; Donato, M.T.; Tolosa, L.; Gonzalez, K.; Ansoar, Y.; Gomez-Lechon, M.J.; Berghe, W.V.; et al. Assessment of the cytotoxic potential of an aqueous-ethanolic extract from Thalassia testudinum angiosperm marine grown in the Caribbean sea. J. Pharm. Pharmacol. 2018, 70, 1553–1560. [Google Scholar] [CrossRef]
- Rajauria, G. Optimization and validation of reverse phase HPLC method for qualitative and quantitative assessment of polyphenols in macroalgae. J. Pharm. Biomed. Anal. 2018, 148, 230–237. [Google Scholar] [CrossRef]
- Raguraman, V.; Mubarak Ali, D.; Narendrakumar, G.; Thirugnanasambandam, R.; Kirubagaran, R.; Thajuddin, N. Unraveling rapid extraction of fucoxanthin from Padina tetrastromatica: Purification, characterization and biomedical application. Process Biochem. 2018, 73, 211–219. [Google Scholar] [CrossRef]
- Nunes, N.; Leça, J.M.; Pereira, A.C.; Pereira, V.; Ferraz, S.; Barreto, M.C.; Marques, J.C.; MAA Pinhero de Carvalho. Evaluation of fucoxanthin contents in macroalgae biomass by vortex-assisted solid-liquid microextraction using high-performance liquid chromatography with photodiode array detection. Algal Res. 2019, 42, 101603. [Google Scholar] [CrossRef]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Characterization of dietary fucoxanthin from Himanthalia elongata brown macroalgae. Food Res. Int. 2017, 99, 995–1001. [Google Scholar] [CrossRef]
- Jaswir, I.; Noviendri, D.; Salleh, H.M.; Taher, M.; Miyashita, K.; Ramli, N. Analysis of fucoxanthin content and purification of all-trans-fucoxanthin from Turbinaria turbinata and Sargassum plagyophyllum by SiO2 open column chromatography and reversed phase-HPLC. J. Liq. Chromatogr. Relat. Technol. 2013, 36, 1340–1354. [Google Scholar] [CrossRef]
- Abd Ghafar, S.Z.; Mediani, A.; Ramli, N.S.; Abas, F. Antioxidant, α-glucosidase, and nitric oxide inhibitory activities of Phyllanthus acidus and LC–MS/MS profile of the active extract. Food Biosci. 2018, 25, 134–140. [Google Scholar] [CrossRef]
- Olate-Gallegos, C.; Barriga, A.; Vergara, C.; Fredes, C.; García, P.; Giménez, B.; Robert, P. Identification of polyphenols from Chilean brown macroalgaes extracts by LC-DAD-ESI-MS/MS. J. Aquat. Food Prod. Technol. 2019, 28, 375–391. [Google Scholar] [CrossRef]
- Pawłowska, K.A.; Strawa, J.; Tomczyk, M.; Granica, S. Changes in the phenolic contents and composition of Persicariaodorata fresh and dried leaves. J. Food Compos. Anal. 2020, 91, 103507. [Google Scholar] [CrossRef]
- Khallouki, F.; Ricarte, I.; Breuer, A.; Owen, R.W. Characterization of phenolic compounds in mature Moroccan Medjool date palm fruits (Phoenix dactylifera) by HPLC-DAD-ESI-MS. J. Food Compos. Anal. 2018, 70, 63–71. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macroalgae | Chlorophyll a (µg/g fw) | Chlorophyll b (µg/g fw) |
|---|---|---|
| S. wightii | 6.41 ± 0.02 | ND |
| U. rigida | 12.22 ± 0.28 * | 13.50 ± 0.14 * |
| Percent Inhibition | ||||||||
|---|---|---|---|---|---|---|---|---|
| S. aureus1 | E. coli1 | S. typhimurium1 | B. subtilis2 | |||||
| Concentration (mg/mL) | SW | UR | SW | UR | SW | UR | SW | UR |
| 0.6 | 62.96 ± 0.37 e | 30.02 ± 0.32 i | 67.36 ± 0.23 d | 31.99 ± 0.32 h | 45.06 ± 0.03 g | 30.73 ± 0.14 j | 67.39 ±1.83 d | 36.03 ± 0.22 h |
| 0.8 | 65.60 ± 0.23 d | 36.44 ± 0.21 h | 66.79 ± 0.13 e | 31.51 ± 0.16 i | 59.80 ± 0.01 d | 38.94 ± 1.56 h | 69.43 ± 0.37 cd | 40.46 ± 3.50 g |
| 1 | 71.19 ± 0.14 c | 37.29 ± 0.21 g | 68.51 ± 0.07 c | 36.05 ± 0.13 g | 64.06 ± 0.01 c | 33.56 ± 0.08 i | 72.02 ± 0.29 c | 44.57 ± 0.29 f |
| 2 | 74.97 ± 0.18 b | 37.42 ± 0.21 g | 74.39 ± 0.17 b | 46.65 ± 0.17 f | 79.81 ± 0.18 b | 49.94 ± 0.22 f | 76.73 ± 0.21 b | 47.45 ± 2.11 f |
| 4 | 88.07 ± 0.18 a | 48.01 ± 0.36 f | 87.32 ± 0.24 a | 66.51 ± 0.19 e | 91.15 ± 0.18 a | 51.30 ± 1.35 e | 85.01 ± 0.37 a | 55.97 ± 3.39 e |
| S. wightii | U. rigida | |||
|---|---|---|---|---|
| Bioaccessibility (%) | SGD | SGID | SGD | SGID |
| TPC | 10.59 ± 0.98 c | 39.91 ± 4.01 a | 6.88 ± 0.94 c | 26.17 ± 1.10 b |
| FRAP | 12.43 ± 0.74 c | 25.64 ± 1.73 a | 10.70 ± 0.18 c | 20.92 ± 0.69 b |
| Bioactive Compounds | S. wightii (mg/kg Dry Weight) | U. rigida (mg/kg Dry Weight) |
|---|---|---|
| Fucoxanthin | 9.27 ± 2.28 | ND |
| Phloroglucinol | 17.96 ± 2.80 | ND |
| Gallic acid | 0.07 ± 0.00 | 0.07 ± 0.00 |
| Quercetin | 0.04 ± 0.02 | 0.04 ± 0.00 |
| Ferulic acid | 1.05 ± 0.00 | 1.07 ± 0.00 |
| Vanillin | 1.55 ± 0.31 | 1.23 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, Y.; Tarafdar, A.; Kumar, D.; Saravanan, C.; Badgujar, P.C.; Pharande, A.; Pareek, S.; Fawole, O.A. Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient. Antioxidants 2022, 11, 993. https://doi.org/10.3390/antiox11050993
Kumar Y, Tarafdar A, Kumar D, Saravanan C, Badgujar PC, Pharande A, Pareek S, Fawole OA. Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient. Antioxidants. 2022; 11(5):993. https://doi.org/10.3390/antiox11050993
Chicago/Turabian StyleKumar, Yogesh, Ayon Tarafdar, Deepak Kumar, Chakkaravarthi Saravanan, Prarabdh C. Badgujar, Aparna Pharande, Sunil Pareek, and Olaniyi Amos Fawole. 2022. "Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient" Antioxidants 11, no. 5: 993. https://doi.org/10.3390/antiox11050993
APA StyleKumar, Y., Tarafdar, A., Kumar, D., Saravanan, C., Badgujar, P. C., Pharande, A., Pareek, S., & Fawole, O. A. (2022). Polyphenols of Edible Macroalgae: Estimation of In Vitro Bio-Accessibility and Cytotoxicity, Quantification by LC-MS/MS and Potential Utilization as an Antimicrobial and Functional Food Ingredient. Antioxidants, 11(5), 993. https://doi.org/10.3390/antiox11050993