
Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fish Maintenance
2.3. Experimental Design
2.4. Biochemical Determinations
2.5. Statistical Analysis
3. Results
3.1. AChE Activity
3.2. Biotransformation Enzymes
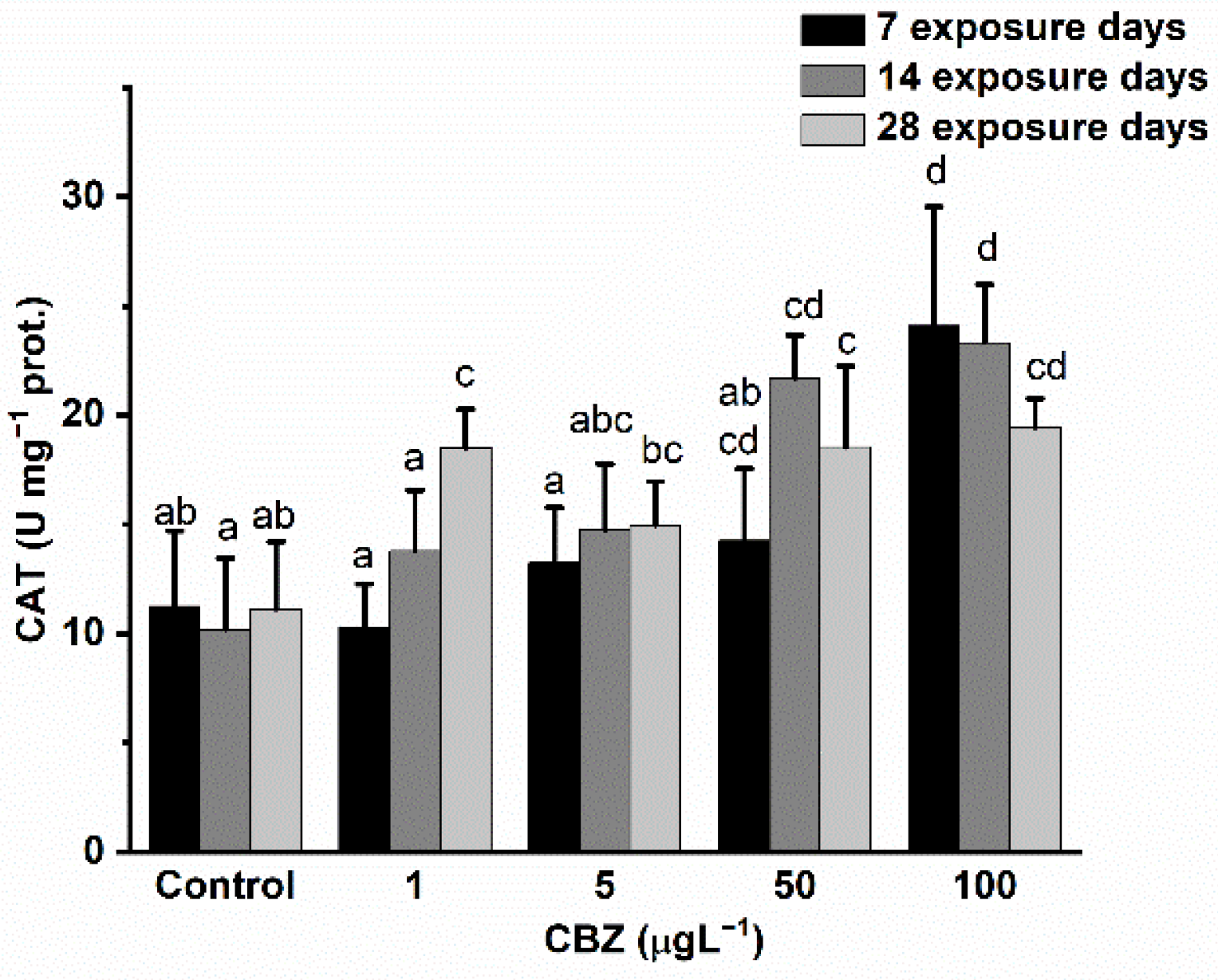
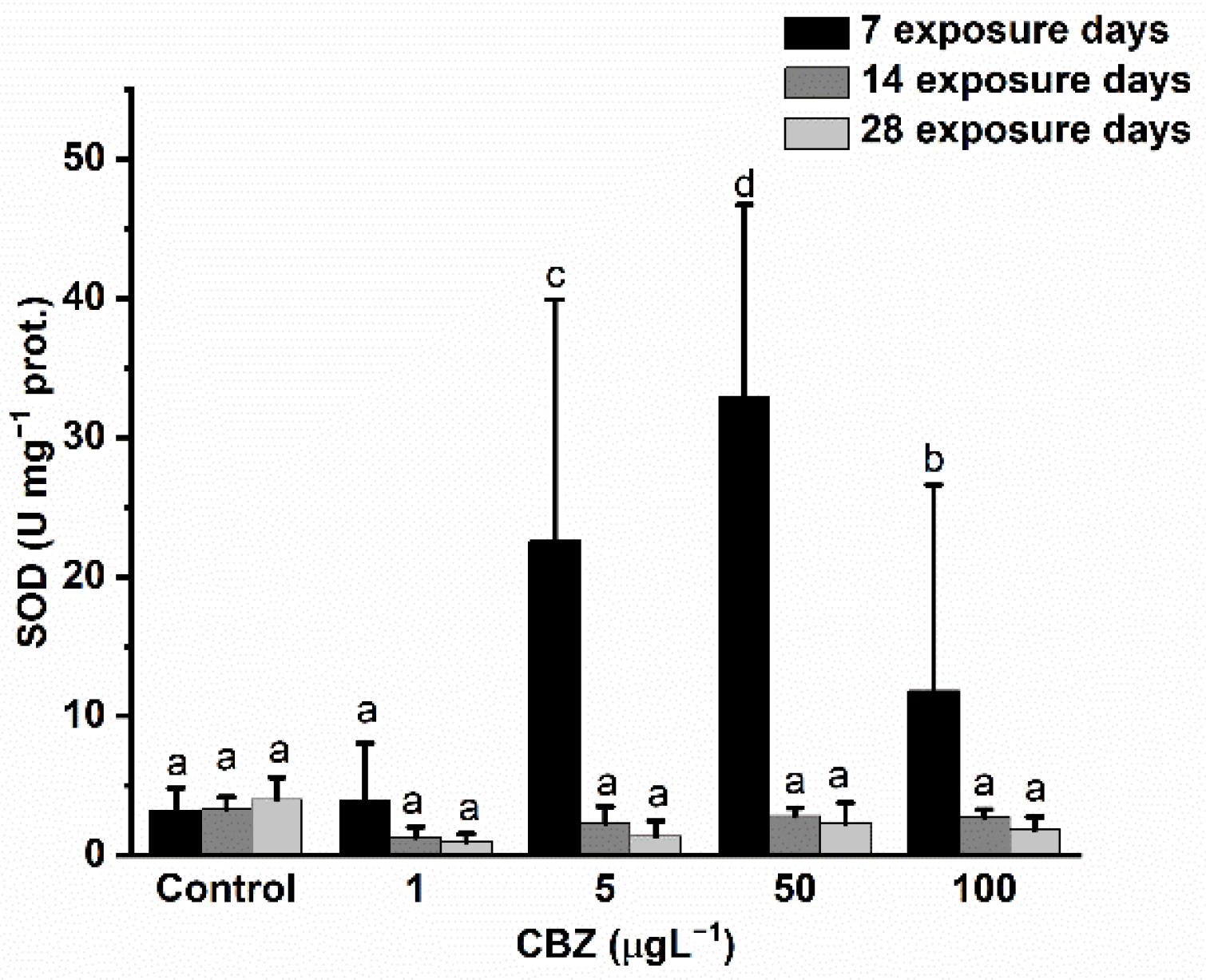
3.3. Antioxidant Defence
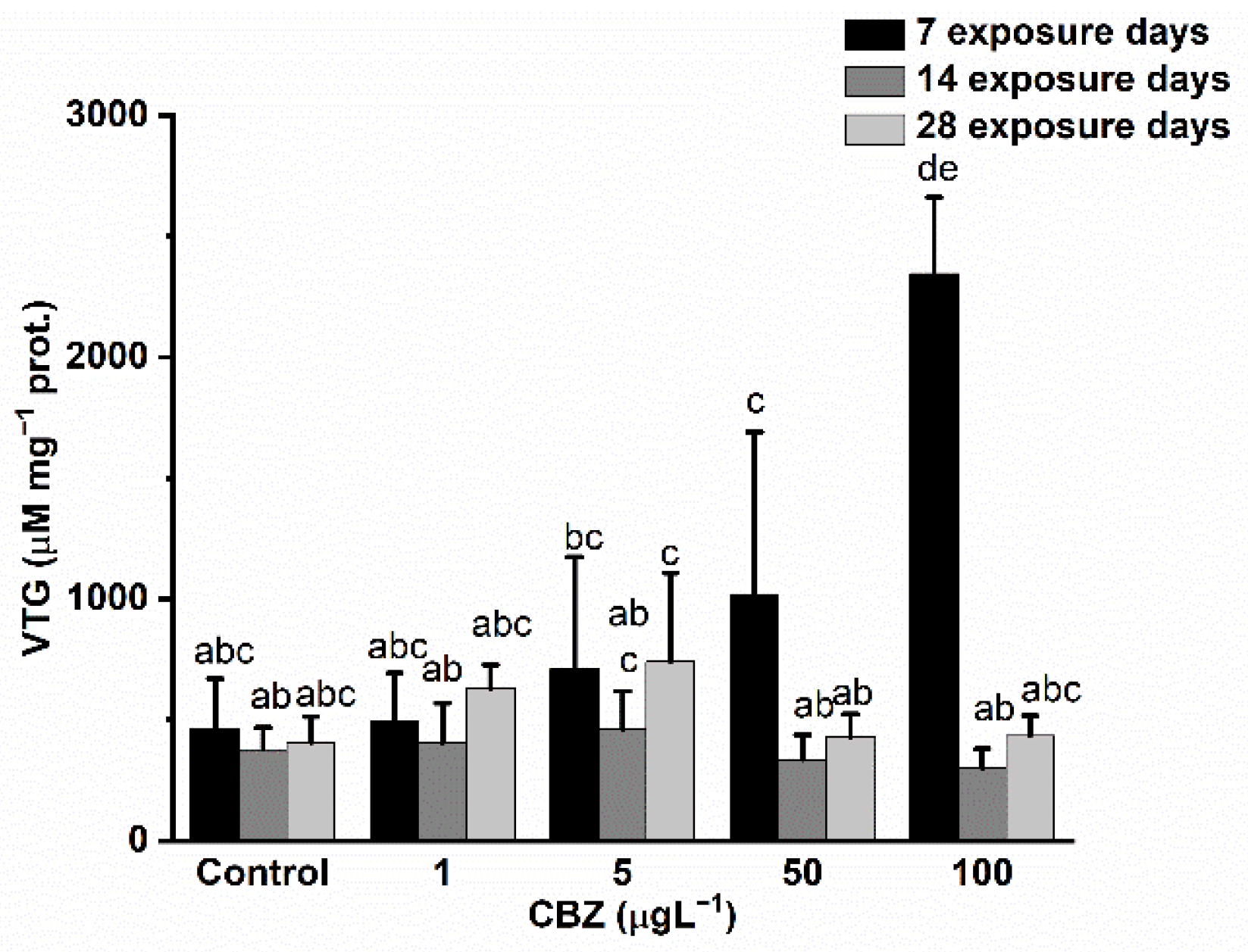
3.4. Damage Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santos, L.H.M.L.M.; Araújo, A.N.; Fachini, A.; Pena, A.; Delerue-Matos, C.; Montenegro, M.C.B.S.M. Ecotoxicological Aspects Related to the Presence of Pharmaceuticals in the Aquatic Environment. J. Hazard. Mater. 2010, 175, 45–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, N.; Pereira, R.; Abrantes, N.; Pereira, J.; Gonçalves, F.; Marques, C.R. Ecotoxicological Effects of Ciprofloxacin on Freshwater Species: Data Integration and Derivation of Toxicity Thresholds for Risk Assessment. Ecotoxicology 2012, 21, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Valdés, M.E.; Huerta, B.; Wunderlin, D.A.; Bistoni, M.A.; Barceló, D.; Rodriguez-Mozaz, S. Bioaccumulation and Bioconcentration of Carbamazepine and Other Pharmaceuticals in Fish under Field and Controlled Laboratory Experiments. Evidences of Carbamazepine Metabolization by Fish. Sci. Total Environ. 2016, 557–558, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Vernouillet, G.; Eullaffroy, P.; Lajeunesse, A.; Blaise, C.; Gagné, F.; Juneau, P. Toxic Effects and Bioaccumulation of Carbamazepine Evaluated by Biomarkers Measured in Organisms of Different Trophic Levels. Chemosphere 2010, 80, 1062–1068. [Google Scholar] [CrossRef]
- Kreke, N.; Dietrich, D.R. Physiological Endpoints for Potential SSRI Interactions in Fish. Crit. Rev. Toxicol. 2008, 38, 215–247. [Google Scholar] [CrossRef] [Green Version]
- LaLone, C.A.; Villeneuve, D.L.; Burgoon, L.D.; Russom, C.L.; Helgen, H.W.; Berninger, J.P.; Tietge, J.E.; Severson, M.N.; Cavallin, J.E.; Ankley, G.T. Molecular Target Sequence Similarity as a Basis for Species Extrapolation to Assess the Ecological Risk of Chemicals with Known Modes of Action. Aquat. Toxicol. 2013, 144–145, 141–154. [Google Scholar] [CrossRef]
- Da Silva Santos, N.; Oliveira, R.; Lisboa, C.A.; Mona e Pinto, J.; Sousa-Moura, D.; Camargo, N.S.; Perillo, V.; Oliveira, M.; Grisolia, C.K.; Domingues, I. Chronic Effects of Carbamazepine on Zebrafish: Behavioral, Reproductive and Biochemical Endpoints. Ecotoxicol. Environ. Saf. 2018, 164, 297–304. [Google Scholar] [CrossRef]
- Breton, H.; Cociglio, M.; Bressolle, F.; Peyriere, H.; Blayac, J.P.; Hillaire-Buys, D. Liquid Chromatography-Electrospray Mass Spectrometry Determination of Carbamazepine, Oxcarbazepine and Eight of Their Metabolites in Human Plasma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 828, 80–90. [Google Scholar] [CrossRef]
- Clara, M.; Strenn, B.; Kreuzinger, N. Carbamazepine as a Possible Anthropogenic Marker in the Aquatic Environment: Investigations on the Behaviour of Carbamazepine in Wastewater Treatment and during Groundwater Infiltration. Water Res. 2004, 38, 947–954. [Google Scholar] [CrossRef]
- Zhang, Y.; Geißen, S.U.; Gal, C. Carbamazepine and Diclofenac: Removal in Wastewater Treatment Plants and Occurrence in Water Bodies. Chemosphere 2008, 73, 1151–1161. [Google Scholar] [CrossRef]
- Wilkinson, J.L.; Boxall, A.B.A.; Kolpin, D.W.; Leung, K.M.Y.; Lai, R.W.S.; Wong, D.; Ntchantcho, R.; Pizarro, J.; Mart, J.; Echeverr, S.; et al. Pharmaceutical Pollution of the World’s Rivers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113947119. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, V.L.; Perino, C.; D’Aco, V.J.; Hartmann, A.; Bechter, R. Human Health Risk Assessment of Carbamazepine in Surface Waters of North America and Europe. Regul. Toxicol. Pharmacol. 2010, 56, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zha, J.; Liang, X.; Li, J.; Wang, Z. Effects of the Human Antiepileptic Drug Carbamazepine on the Behavior, Biomarkers, and Heat Shock Proteins in the Asian Clam Corbicula Fluminea. Aquat. Toxicol. 2014, 155, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Brandão, F.P.; Rodrigues, S.; Castro, B.B.; Gonçalves, F.; Antunes, S.C.; Nunes, B. Short-Term Effects of Neuroactive Pharmaceutical Drugs on a Fish Species: Biochemical and Behavioural Effects. Aquat. Toxicol. 2013, 144–145, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Sim, W.J.; Lee, J.W.; Lee, E.S.; Shin, S.K.; Hwang, S.R.; Oh, J.E. Occurrence and Distribution of Pharmaceuticals in Wastewater from Households, Livestock Farms, Hospitals and Pharmaceutical Manufactures. Chemosphere 2011, 82, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.; Gawlik, B.M.; Locoro, G.; Rimaviciute, E.; Contini, S.; Bidoglio, G. EU-Wide Survey of Polar Organic Persistent Pollutants in European River Waters. Environ. Pollut. 2009, 157, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Staszny, A.; Dobosy, P.; Maasz, G.; Szalai, Z.; Jakab, G.; Pirger, Z.; Szeberenyi, J.; Molnar, E.; Pap, L.O.; Juhasz, V.; et al. Effects of Pharmaceutically Active Compounds (PhACs) on Fish Body and Scale Shape in Natural Waters. PeerJ 2021, 9, e10642. [Google Scholar] [CrossRef]
- Kondor, A.C.; Molnár, É.; Vancsik, A.; Filep, T.; Szeberényi, J.; Szabó, L.; Maász, G.; Pirger, Z.; Weiperth, A.; Ferincz, Á.; et al. Occurrence and Health Risk Assessment of Pharmaceutically Active Compounds in Riverbank Filtrated Drinking Water. J. Water Process Eng. 2021, 41, 102039. [Google Scholar] [CrossRef]
- Ayano, G. Bipolar Disorders and Carbamazepine: Pharmacokinetics, Pharmacodynamics, Therapeutic Effects and Indications of Carbamazepine: Review of Articles. J. Neuropsychopharmacol. Ment. Health 2016, 1, 112. [Google Scholar] [CrossRef] [Green Version]
- Brodie, M.J.; Richens, A.; Yuen, A.; UK Lamotrigine/Carbamazepine Monotherapy Trial Group. Double-Blind Comparison of Lamotrigine and Carbamazepine in Newly Diagnosed Epilepsy. Lancet 1995, 345, 476–479. [Google Scholar] [CrossRef]
- Xin, X.; Huang, G.; Liu, X.; An, C.; Yao, Y.; Weger, H.; Zhang, P.; Chen, X. Molecular Toxicity of Triclosan and Carbamazepine to Green Algae Chlorococcum Sp.: A Single Cell View Using Synchrotron-Based Fourier Transform Infrared Spectromicroscopy. Environ. Pollut. 2017, 226, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Oropesa, A.L.; Floro, A.M.; Palma, P. Assessment of the Effects of the Carbamazepine on the Endogenous Endocrine System of Daphnia Magna. Environ. Sci. Pollut. Res. 2016, 23, 17311–17321. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Li, P.; Rodina, M.; Randak, T. Effect of Human Pharmaceutical Carbamazepine on the Quality Parameters and Oxidative Stress in Common Carp (Cyprinus carpio L.) Spermatozoa. Chemosphere 2010, 80, 530–534. [Google Scholar] [CrossRef]
- Galus, M.; Rangarajan, S.; Lai, A.; Shaya, L.; Balshine, S.; Wilson, J.Y. Effects of Chronic, Parental Pharmaceutical Exposure on Zebrafish (Danio rerio) Offspring. Aquat. Toxicol. 2014, 151, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Zlabek, V.; Velisek, J.; Grabic, R.; Machova, J.; Kolarova, J.; Li, P.; Randak, T. Acute Toxicity of Carbamazepine to Juvenile Rainbow Trout (Oncorhynchus mykiss): Effects on Antioxidant Responses, Hematological Parameters and Hepatic EROD. Ecotoxicol. Environ. Saf. 2011, 74, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Gasca-Pérez, E.; Galar-Martínez, M.; García-Medina, S.; Pérez-Coyotl, I.A.; Ruiz-Lara, K.; Cano-Viveros, S.; Pérez-Pastén Borja, R.; Gómez-Oliván, L.M. Short-Term Exposure to Carbamazepine Causes Oxidative Stress on Common Carp (Cyprinus carpio). Environ. Toxicol. Pharmacol. 2019, 66, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Karan, V.; Vitorović, S.; Tutundžić, V.; Poleksić, V. Functional Enzymes Activity and Gill Histology of Carp after Copper Sulfate Exposure and Recovery. Ecotoxicol. Environ. Saf. 1998, 40, 49–55. [Google Scholar] [CrossRef]
- Oruç, E.Ö.; Üner, N. Marker Enzyme Assesment in the Liver of Cyprinus carpio (L.) Exposed to 2,4-D and Azinphosmethyl. J. Biochem. Mol. Toxicol. 2002, 16, 182–188. [Google Scholar] [CrossRef]
- Saucedo-Vence, K.; Dublán-García, O.; López-Martínez, L.X.; Morachis-Valdes, G.; Galar-Martínez, M.; Islas-Flores, H.; Gómez-Oliván, L.M. Short and Long-Term Exposure to Diclofenac Alter Oxidative Stress Status in Common Carp Cyprinus carpio. Ecotoxicology 2015, 24, 527–539. [Google Scholar] [CrossRef]
- Hai, F.I.; Yang, S.; Asif, M.B.; Sencadas, V.; Shawkat, S.; Sanderson-Smith, M.; Gorman, J.; Xu, Z.Q.; Yamamoto, K. Carbamazepine as a Possible Anthropogenic Marker in Water: Occurrences, Toxicological Effects, Regulations and Removal by Wastewater Treatment Technologies. Water 2018, 10, 107. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wills, E.D. Evaluation of Lipid Peroxidation in Lipids and Biological Membranes. In Biochemical Toxicology: A Practical Approach; Snell, K., Mullock, B., Eds.; IRL Press: Washington, DC, USA, 1987; pp. 127–152. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Guilhermino, L.; Lopes, M.C.; Carvalho, A.P.; Soares, A.M.V.M. Inhibition of Acetylcholinesterase Activity as Effect Criterion in Acute Tests with Juvenile Daphnia Magna. Chemosphere 1996, 32, 727–738. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S Transferases. The First Enzymatic Step in Mercapturic Acid Formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. Purification and Characterization of the Flavoenzyme Glutathione Reductase from Rat Liver. J. Biol. Chem. 1975, 250, 5475–5480. [Google Scholar] [CrossRef]
- Vassault, A. Lactate Dehydrogenase. In Methods of Enzymatic Analysis; Bergmeye, H.U., Bergmeyer, J., Gra, I.M., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 118–126. [Google Scholar]
- Diamantino, T.C.; Almeida, E.; Soares, A.M.V.M.; Guilhermino, L. Lactate Dehydrogenase Activity as an Effect Criterion in Toxicity Tests with Daphnia Magna Straus. Chemosphere 2001, 45, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Burke, M.D.; Mayer, R.T. Ethoxyresorufin: Direct Fluorimetric Assay of a Microsomal O Dealkylation Which Is Preferentially Inducible by 3 Methylcholanthrene. Drug Metab. Dispos. 1974, 2, 583–588. [Google Scholar]
- Olive, P.L. DNA Precipitation Assay: A Rapid and Simple Method for Detecting DNA Damage in Mammalian Cells. Environ. Mol. Mutagen. 1988, 11, 487–495. [Google Scholar] [CrossRef]
- Blaise, C.; Gagné, F.; Pellerin, J.; Hansen, P.D. Determination of Vitellogenin-like Properties in Mya arenaria Hemolymph (Saguenay Fjord, Canada): A Potential Biomarker for Endocrine Disruption. Environ. Toxicol. 1999, 14, 455–465. [Google Scholar] [CrossRef]
- Stanton, M.G. Colorimetric Determination of Inorganic Phosphate in the Presence of Biological Material and Adenosine Triphosphate. Anal. Biochem. 1968, 22, 27–34. [Google Scholar] [CrossRef]
- Crapo, J.D.; McCord, J.M.; Fridovich, I. Preparation and Assay of Superioxide Dismutases. In Methods in Enzymology; Academic Press: New York, NY, USA, 1978; Volume 53, pp. 382–393. [Google Scholar]
- Nkoom, M.; Lu, G.; Liu, J.; Dong, H. Biological Uptake, Depuration and Biochemical Effects of Diclofenac and Carbamazepine in Carassius carassius. Ecotoxicol. Environ. Saf. 2020, 205, 111106. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Martínez, G.V.; DelValls, T.A.; Martín-Díaz, M.L. General Stress, Detoxification Pathways, Neurotoxicity and Genotoxicity Evaluated in Ruditapes philippinarum Exposed to Human Pharmaceuticals. Ecotoxicol. Environ. Saf. 2016, 124, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.S.; Kim, B.M.; Jeong, C.B.; Park, H.G.; Leung, K.M.Y.; Lee, Y.M.; Lee, J.S. Effect of Pharmaceuticals Exposure on Acetylcholinesterase (AchE) Activity and on the Expression of AchE Gene in the Monogonont Rotifer, Brachionus koreanus. Comp. Biochem. Physiol.-C Toxicol. Pharmacol. 2013, 158, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, S.; Schiedek, D.; Dippner, J.W. Effect of Temperature and Salinity on Acetylcholinesterase Activity, a Common Pollution Biomarker, in Mytilus Sp. from the South-Western Baltic Sea. J. Exp. Mar. Biol. Ecol. 2005, 320, 93–103. [Google Scholar] [CrossRef]
- Ferreira, G.K.; Carvalho-Silva, M.; Gonçalves, C.L.; Vieira, J.S.; Scaini, G.; Ghedim, F.V.; Deroza, P.F.; Zugno, A.I.; Pereira, T.C.B.; Oliveira, G.M.T.; et al. L-Tyrosine Administration Increases Acetylcholinesterase Activity in Rats. Neurochem. Int. 2012, 61, 1370–1374. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Yang, L.; Yu, L.; Lin, B.; Hou, Y.; Wu, J.; Huang, Q.; Han, Y.; Guo, L.; Ouyang, Q.; et al. Acetylcholinesterase Is Associated with Apoptosis in β Cells and Contributes to Insulin-Dependent Diabetes Mellitus Pathogenesis. Acta Biochim. Biophys. Sin. 2012, 44, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.J.; Yang, L.; Zhao, Q.; Caen, J.P.; He, H.Y.; Jin, Q.H.; Guo, L.H.; Alemany, M.; Zhang, L.Y.; Shi, Y.F. Induction of Acetylcholinesterase Expression during Apoptosis in Various Cell Types. Cell Death Differ. 2002, 9, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Chen, R.; Wang, M.; Zha, J. Carbamazepine at Environmentally Relevant Concentrations Caused DNA Damage and Apoptosis in the Liver of Chinese Rare Minnows (Gobiocypris rarus) by the Ras/Raf/ERK/P53 Signaling Pathway. Environ. Pollut. 2021, 270, 116245. [Google Scholar] [CrossRef]
- Tõugu, V.; Kesvatera, T. Role of Ionic Interactions in Cholinesterase Catalysis. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1996, 1298, 12–30. [Google Scholar] [CrossRef]
- Jönsson, E.M.; Abrahamson, A.; Brunström, B.; Brandt, I. Cytochrome P4501A Induction in Rainbow Trout Gills and Liver Following Exposure to Waterborne Indigo, Benzo[a]Pyrene and 3,3′,4,4′,5-Pentachlorobiphenyl. Aquat. Toxicol. 2006, 79, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative Stress and Antioxidant Defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish Bioaccumulation and Biomarkers in Environmental Risk Assessment: A Review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Viarengo, A.; Canesi, L.; Garcia Martinez, P.; Peters, L.D.; Livingstone, D.R. Pro-Oxidant Processes and Antioxidant Defence Systems in the Tissues of the Antarctic Scallop (Adamussium colbecki) Compared with the Mediterranean Scallop (Pecten jacobaeus). Comp. Biochem. Physiol.-Part B Biochem. 1995, 111, 119–126. [Google Scholar] [CrossRef]
- Li, Z.H.; Velisek, J.; Zlabek, V.; Grabic, R.; Machova, J.; Kolarova, J.; Randak, T. Hepatic Antioxidant Status and Hematological Parameters in Rainbow Trout, Oncorhynchus mykiss, after Chronic Exposure to Carbamazepine. Chem. Biol. Interact. 2010, 183, 98–104. [Google Scholar] [CrossRef]
- Oakes, K.D.; Van Der Kraak, G.J. Utility of the TBARS Assay in Detecting Oxidative Stress in White Sucker (Catostomus commersoni) Populations Exposed to Pulp Mill Effluent. Aquat. Toxicol. 2003, 63, 447–463. [Google Scholar] [CrossRef]
- Bolognesi, C.; Hayashi, M. Micronucleus Assay in Aquatic Animals. Mutagenesis 2011, 26, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Pacheco, M.; Santos, M.A. Cytochrome P4501A, Genotoxic and Stress Responses in Golden Grey Mullet (Liza aurata) Following Short-Term Exposure to Phenanthrene. Chemosphere 2007, 66, 1284–1291. [Google Scholar] [CrossRef]
- Osman, A.G.M.; Reheem, A.-E.; El Baset, M.A.; AbuelFadl, K.Y.; Rab, A.G.G. Enzymatic and Histopathologic Biomarkers as Indicators of Aquatic Pollution in Fishes. Nat. Sci. 2010, 2, 1302–1311. [Google Scholar] [CrossRef] [Green Version]
- Malarvizhi, A.; Kavitha, C.; Saravanan, M.; Ramesh, M. Carbamazepine (CBZ) Induced Enzymatic Stress in Gill, Liver and Muscle of a Common Carp, Cyprinus carpio. J. King Saud Univ.-Sci. 2012, 24, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Emmerson, J.; Korsgaard, B.; Petersen, I. Dose Response Kinetics of Serum Vitellogenin, Liver DNA, RNA, Protein and Lipid after Induction by Estradiol-17 Beta in Male Flounders (Platichthys flesus L.). Comp. Biochem. Physiol. 1979, 63, 1–6. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Csenki, Z.; Ivánovics, B.; Bock, I.; Csorbai, B.; Molnár, J.; Vásárhelyi, E.; Griffitts, J.; Ferincz, Á.; Urbányi, B.; et al. Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio). Antioxidants 2022, 11, 1136. https://doi.org/10.3390/antiox11061136
Liang X, Csenki Z, Ivánovics B, Bock I, Csorbai B, Molnár J, Vásárhelyi E, Griffitts J, Ferincz Á, Urbányi B, et al. Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio). Antioxidants. 2022; 11(6):1136. https://doi.org/10.3390/antiox11061136
Chicago/Turabian StyleLiang, Xinyue, Zsolt Csenki, Bence Ivánovics, Illés Bock, Balázs Csorbai, József Molnár, Erna Vásárhelyi, Jeffrey Griffitts, Árpád Ferincz, Béla Urbányi, and et al. 2022. "Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio)" Antioxidants 11, no. 6: 1136. https://doi.org/10.3390/antiox11061136
APA StyleLiang, X., Csenki, Z., Ivánovics, B., Bock, I., Csorbai, B., Molnár, J., Vásárhelyi, E., Griffitts, J., Ferincz, Á., Urbányi, B., & Ács, A. (2022). Biochemical Marker Assessment of Chronic Carbamazepine Exposure at Environmentally Relevant Concentrations in Juvenile Common Carp (Cyprinus carpio). Antioxidants, 11(6), 1136. https://doi.org/10.3390/antiox11061136