
Effects of Dietary Yeast β-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection
2.3. Carcass Traits
2.4. Meat Quality
2.5. Texture Characteristics
2.6. Muscle Fatty Acid Profiles
2.7. Plasma and Skeletal Muscle Biochemical Parameters
2.8. RNA Extraction and qRT-PCR Analysis
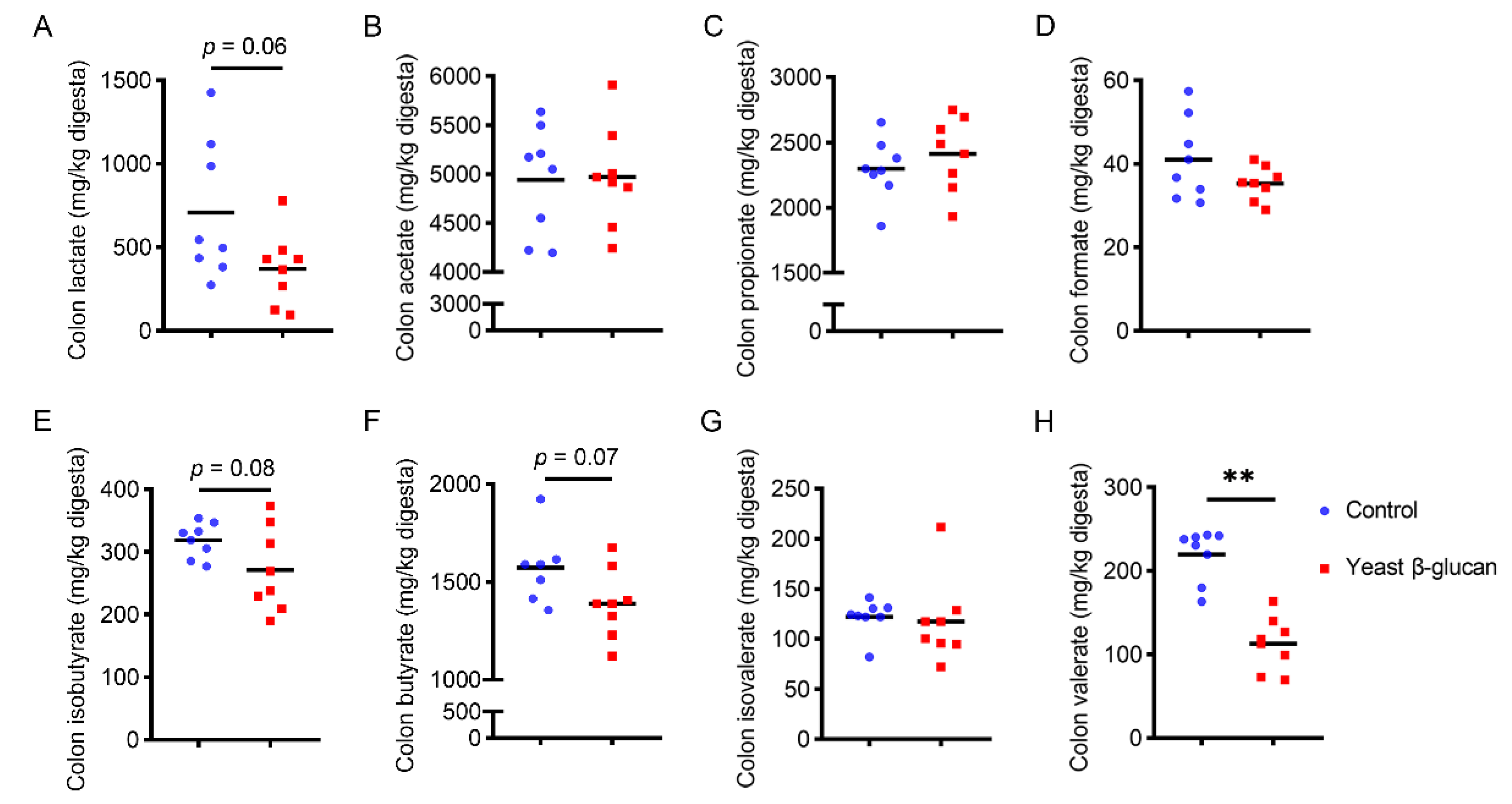
2.9. SCFAs Concentration in Colonic Digesta
2.10. DNA Extraction and PCR Amplification
2.11. DNA Illumina miSeq and Sequence Data Processing
2.12. Statistical Analysis
3. Results
3.1. Immunological Stress
3.2. Carcass Traits and Meat Quality
3.3. Texture Profile Analysis and Fatty Acid Composition of Fresh Meat
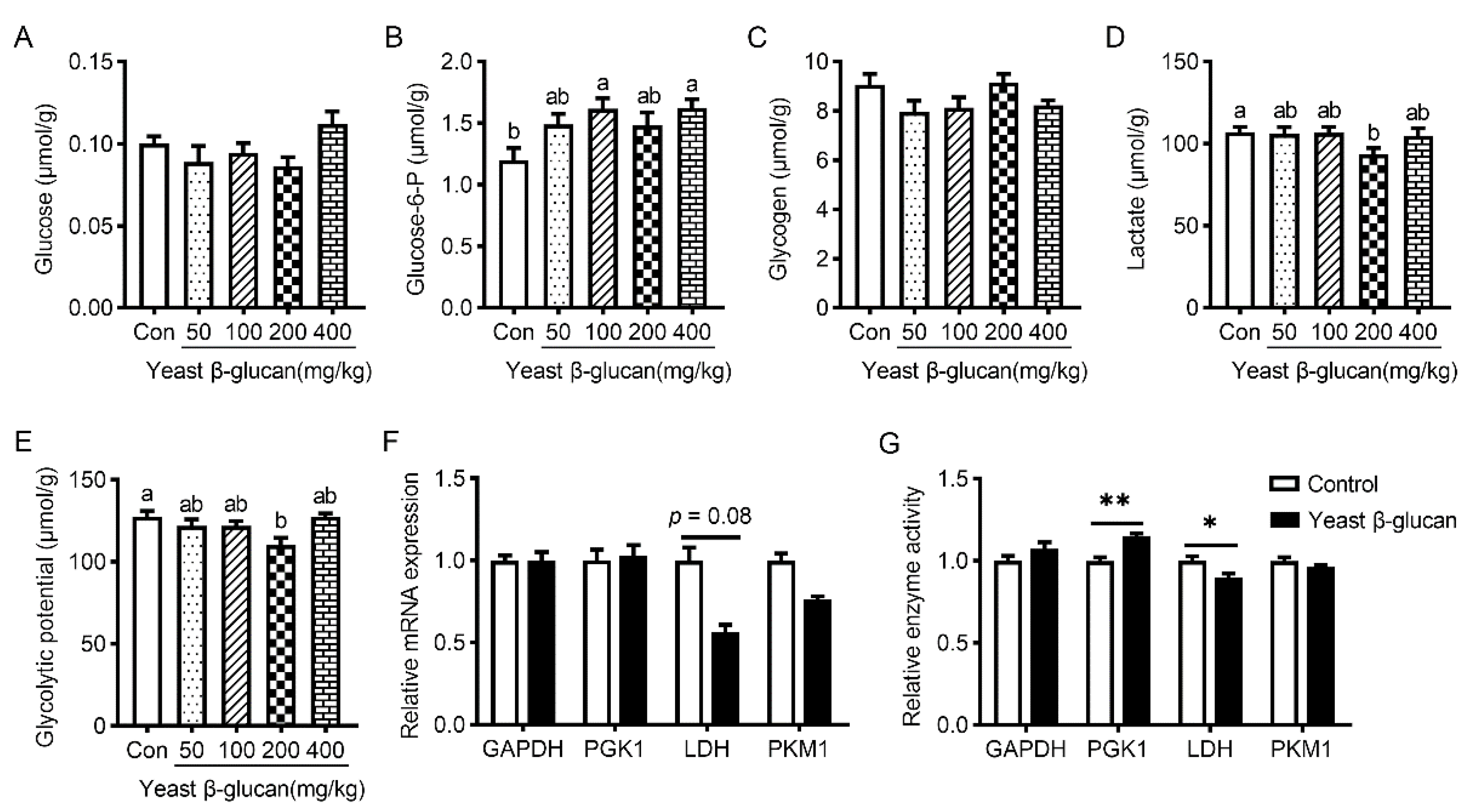
3.4. Muscle Glycolytic Potential
3.5. Muscle Fiber Characteristics
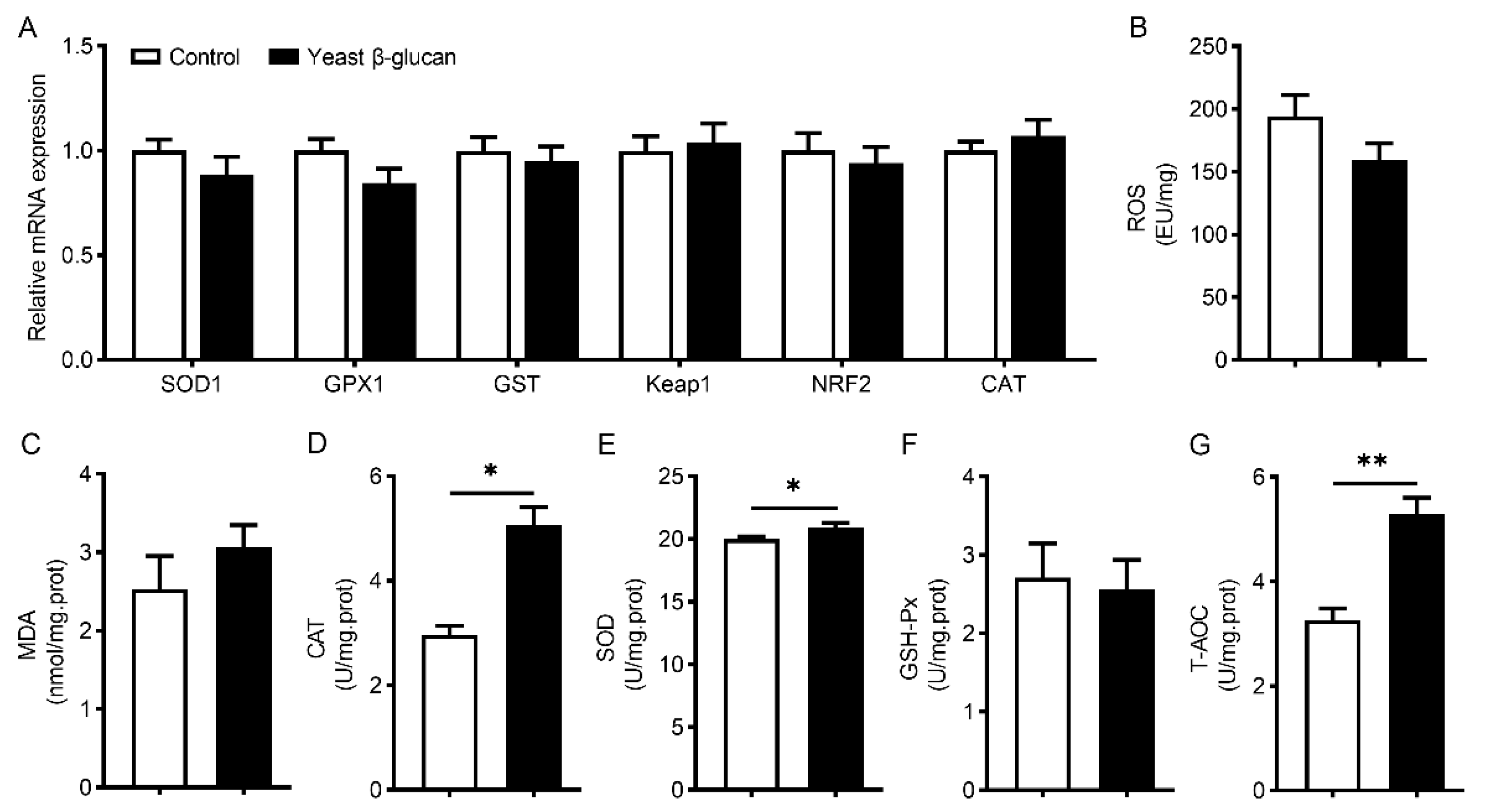
3.6. Muscle Antioxidant Activities
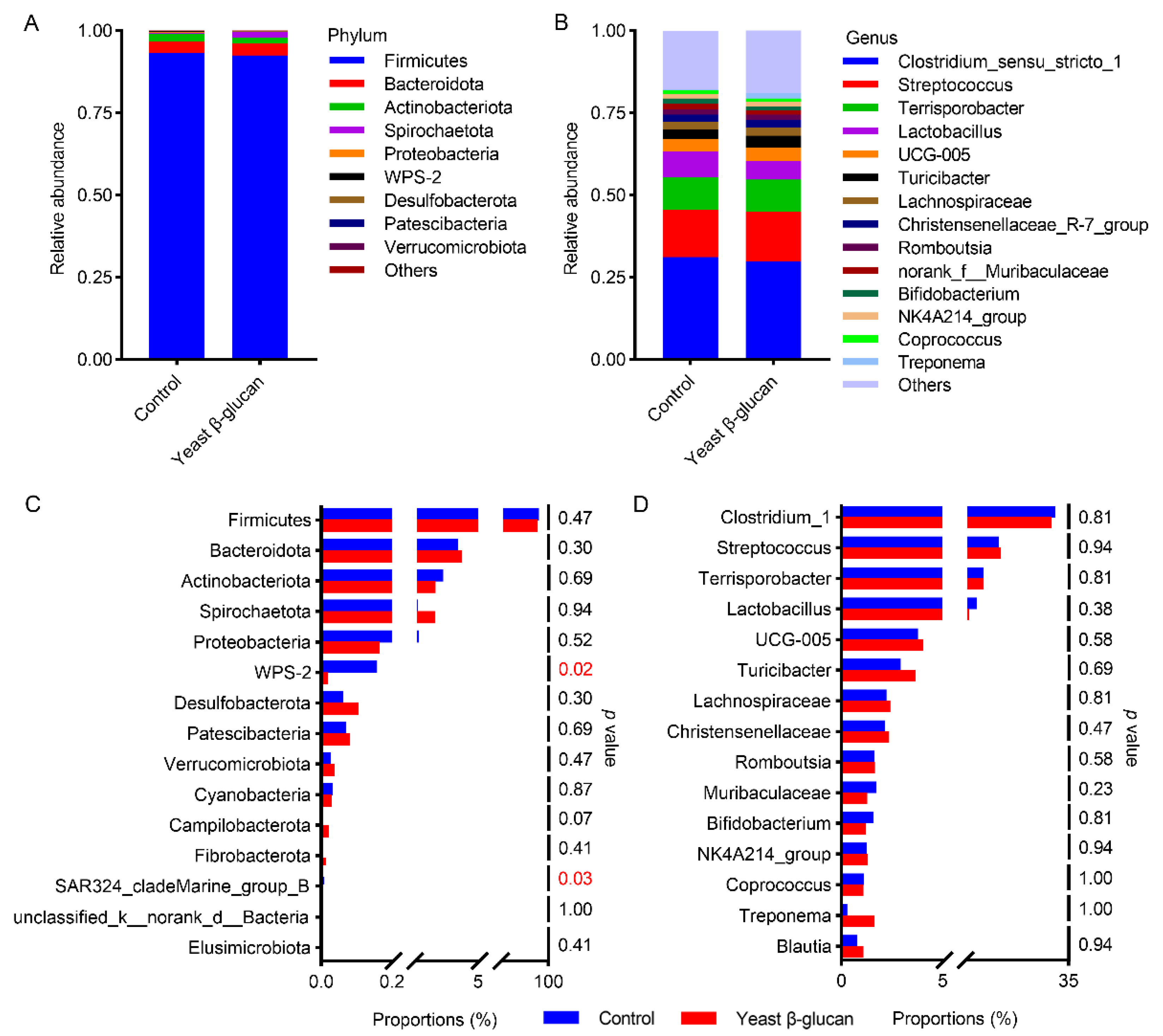
3.7. Responses of Colonic Microbiota to Dietary Yeast β-Glucan Supplementation
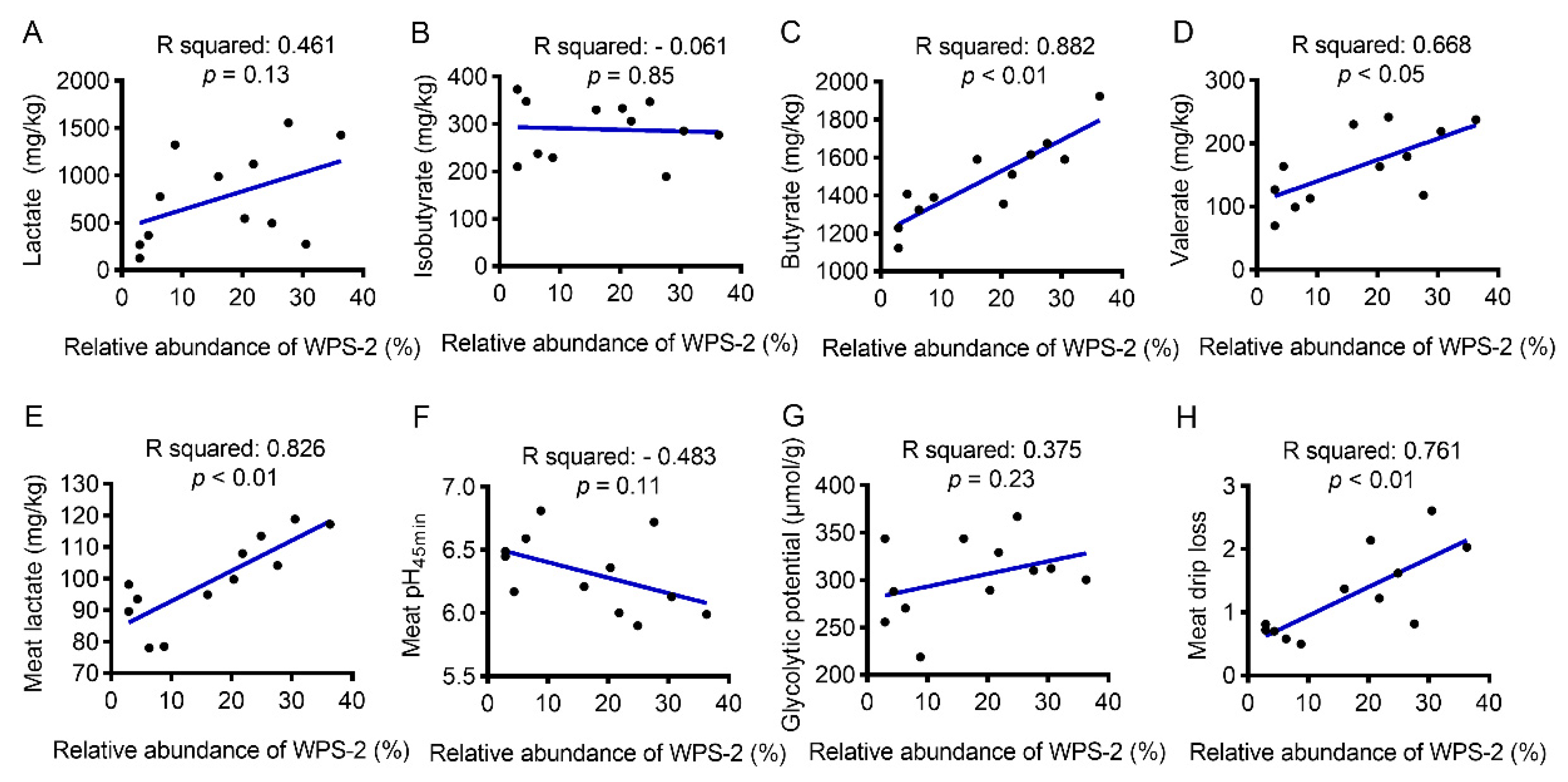
3.8. Correlation between WPS-2 Abundance and SCFAs Concentration or Meat Drip-Loss-Related Biochemical Indices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Song, S.; Cheng, H.; Jung, E.Y.; Joo, S.T.; Kim, G.D. Muscle fiber characteristics on chop surface of pork loin (M. longissimus thoracis et lumborum) associated with muscle fiber pennation angle and their relationships with pork loin quality. Food Sci. Anim. Resour. 2020, 40, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Fischer, K. Drip loss in pork: Influencing factors and relation to further meat quality traits. J. Anim. Breed Genet. 2007, 124, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhou, L.; Zhang, J.; Liu, X.; Zhang, Y.; Cai, L.; Zhang, W.; Cui, L.; Yang, J.; Ji, J.; et al. A large-scale comparison of meat quality and intramuscular fatty acid composition among three Chinese indigenous pig breeds. Meat Sci. 2020, 168, 108182. [Google Scholar] [CrossRef] [PubMed]
- Rossi, R.; Pastorelli, G.; Cannata, S.; Tavaniello, S.; Maiorano, G.; Corino, C. Effect of long term dietary supplementation with plant extract on carcass characteristics meat quality and oxidative stability in pork. Meat Sci. 2013, 95, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Monin, G.; Sellier, P. Pork of low technological quality with a normal rate of muscle pH fall in the immediate post-mortem period: The case of the Hampshire breed. Meat Sci. 1985, 13, 49–63. [Google Scholar] [CrossRef]
- Joo, S.T.; Kauffman, R.G.; Kim, B.C.; Park, G.B. The relationship of sarcoplasmic and myofibrillar protein solubility to colour and water-holding capacity in porcine longissimus muscle. Meat Sci. 1999, 52, 291–297. [Google Scholar] [CrossRef]
- England, E.M.; Scheffler, T.L.; Kasten, S.C.; Matarneh, S.K.; Gerrard, D.E. Exploring the unknowns involved in the transformation of muscle to meat. Meat Sci. 2013, 95, 837–843. [Google Scholar] [CrossRef]
- Hamilton, D.N.; Miller, K.D.; Ellis, M.; McKeith, F.K.; Wilson, E.R. Relationships between longissimus glycolytic potential and swine growth performance, carcass traits, and pork quality. J. Anim. Sci. 2003, 81, 2206–2212. [Google Scholar] [CrossRef] [Green Version]
- Asghar, A. Perspectives on warmed-over flavor. Food Technol. 1988, 42, 102–108. [Google Scholar]
- Domínguez, R.; Pateiro, M.; Gagaoua, M.; Barba, F.J.; Zhang, W.; Lorenzo, J.M. A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 2019, 8, 429. [Google Scholar] [CrossRef] [Green Version]
- Cavazos, D.A.; deGraffenried, M.J.; Apte, S.A.; Bowers, L.W.; Whelan, K.A.; deGraffenried, L.A. Obesity promotes aerobic glycolysis in prostate cancer cells. Nutr. Cancer 2014, 66, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Chen, D.; Zhang, F.; Willsky, G.R.; Crans, D.C.; Ding, W. Effects of vanadium (III, IV, V)-chlorodipicolinate on glycolysis and antioxidant status in the liver of STZ-induced diabetic rats. J. Inorg. Biochem. 2014, 136, 47–56. [Google Scholar] [CrossRef] [PubMed]
- De Marco Castro, E.; Calder, P.C.; Roche, H.M. β-1,3/1,6-glucans and immunity: State of the art and future directions. Mol. Nutr. Food Res. 2021, 65, e1901071. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ali, R.A.; Qureshi, M.A. The influence of beta-glucan on immune responses in broiler chicks. Immunopharmacol Immunotoxicol 2003, 25, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by beta-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Kayali, H.; Ozdag, M.F.; Kahraman, S.; Aydin, A.; Gonul, E.; Sayal, A.; Odabasi, Z.; Timurkaynak, E. The antioxidant effect of beta-Glucan on oxidative stress status in experimental spinal cord injury in rats. Neurosurg. Rev. 2005, 28, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R.; Nowacka-Jechalke, N.; Juda, M.; Malm, A. The preliminary study of prebiotic potential of polish wild mushroom polysaccharides: The stimulation effect on Lactobacillus strains growth. Eur. J. Nutr. 2018, 57, 1511–1521. [Google Scholar] [CrossRef] [Green Version]
- Machová, E.; Bystrický, S. Antioxidant capacities of mannans and glucans are related to their susceptibility of free radical degradation. Int. J. Biol. Macromol. 2013, 61, 308–311. [Google Scholar] [CrossRef]
- Dietrich-Muszalska, A.; Olas, B.; Kontek, B.; Rabe-Jabłońska, J. Beta-glucan from Saccharomyces cerevisiae reduces plasma lipid peroxidation induced by haloperidol. Int. J. Biol. Macromol. 2011, 49, 113–116. [Google Scholar] [CrossRef]
- Yu, C.; Chen, H.; Du, D.; Lv, W.; Li, S.; Li, D. β-Glucan from Saccharomyces cerevisiae alleviates oxidative stress in LPS-stimulated RAW264.7 cells via Dectin-1/Nrf2/HO-1 signaling pathway. Cell Stress Chaperones. 2021, 26, 629–637. [Google Scholar] [CrossRef]
- Wu, C.; Xu, Q.; Wang, R.; Qin, L.; Peng, X.; Hu, L.; Liu, Y.; Fang, Z.; Lin, Y.; Xu, S.; et al. Effects of dietary β-glucan supplementation on growth performance and immunological and metabolic parameters of weaned pigs administered with Escherichia coli lipopolysaccharide. Food Funct. 2018, 9, 3338–3343. [Google Scholar] [CrossRef] [PubMed]
- Hahn, T.W.; Lohakare, J.D.; Lee, S.L.; Moon, W.K.; Chae, B.J. Effects of supplementation of beta-glucans on growth performance, nutrient digestibility, and immunity in weanling pigs. J. Anim. Sci. 2006, 84, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ou, J.; Luo, Z.; Kim, I.H. Effect of dietary β-1,3-glucan supplementation and heat stress on growth performance, nutrient digestibility, meat quality, organ weight, ileum microbiota, and immunity in broilers. Poult. Sci. 2020, 99, 4969–4977. [Google Scholar] [CrossRef]
- Luo, J.; Zeng, D.; Cheng, L.; Mao, X.; Yu, J.; Yu, B.; Chen, D. Dietary β-glucan supplementation improves growth performance, carcass traits and meat quality of finishing pigs. Anim. Nutr. 2019, 5, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]
- Lahiri, S.; Kim, H.; Garcia-Perez, I.; Reza, M.M.; Martin, K.A.; Kundu, P.; Cox, L.M.; Selkrig, J.; Posma, J.M.; Zhang, H.; et al. The gut microbiota influences skeletal muscle mass and function in mice. Sci. Transl. Med. 2019, 11, eaan5662. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Lyu, W.; Hong, Q.; Zhang, X.; Yang, H.; Xiao, Y. Gut microbiota influence lipid metabolism of skeletal muscle in pigs. Front. Nutr. 2021, 8, 675445. [Google Scholar] [CrossRef]
- Chen, C.; Fang, S.; Wei, H.; He, M.; Fu, H.; Xiong, X.; Zhou, Y.; Wu, J.; Gao, J.; Yang, H.; et al. Prevotella copri increases fat accumulation in pigs fed with formula diets. Microbiome 2021, 9, 175. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, B.; Yu, J.; Zheng, P.; Huang, Z.; Luo, Y.; Luo, J.; Mao, X.; Yan, H.; He, J.; et al. Butyrate promotes slow-twitch myofiber formation and mitochondrial biogenesis in finishing pigs via inducing specific microRNAs and PGC-1α expression1. J. Anim. Sci. 2019, 97, 3180–3192. [Google Scholar] [CrossRef]
- Jiao, A.; Diao, H.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, Y.; Luo, J.; Wang, Q.; Wang, H.; et al. Infusion of short chain fatty acids in the ileum improves the carcass traits, meat quality and lipid metabolism of growing pigs. Anim. Nutr. 2021, 7, 94–100. [Google Scholar] [CrossRef]
- de Vries, H.; Geervliet, M.; Jansen, C.A.; Rutten, V.; van Hees, H.; Groothuis, N. Impact of yeast-derived β-glucans on the porcine gut microbiota and immune system in early life. Microorganisms 2020, 8, 1573. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shao, Y.; Guo, Y.; Yuan, J. Enhancement of peripheral blood CD8+ T cells and classical swine fever antibodies by dietary beta-1,3/1,6-glucan supplementation in weaned piglets. Transbound. Emerg. Dis. 2008, 55, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, M.; Yan, E.; Wang, Y.; Ma, C.; Zhang, P.; Yin, J. Dietary malic acid supplementation induces skeletal muscle fiber-type transition of weaned piglets and further improves meat quality of finishing pigs. Front. Nutr. 2021, 8, 825495. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, X.; Zhu, Z.; Jiao, N.; Qiu, K.; Yin, J. Surplus dietary isoleucine intake enhanced monounsaturated fatty acid synthesis and fat accumulation in skeletal muscle of finishing pigs. J. Anim. Sci. Biotechnol. 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, C.; Zhao, J.; Xu, H.; Hou, Q.; Zhang, H. Lactobacillus casei Zhang and vitamin K2 prevent intestinal tumorigenesis in mice via adiponectin-elevated different signaling pathways. Oncotarget 2017, 8, 24719–24727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wasielewska, M.; Banel, A.; Zygmunt, B. Determination of selected organic acids in animal farm water samples by ion chromatography. Int. J. Chem. Eng. Appl. 2012, 3, 178–181. [Google Scholar]
- Wu, Y.; Ma, N.; Song, P.; He, T.; Levesque, C.; Bai, Y.; Zhang, A.; Ma, X. Grape seed proanthocyanidin affects lipid metabolism via changing gut microflora and enhancing propionate production in weaned pigs. J. Nutr. 2019, 149, 1523–1532. [Google Scholar] [CrossRef]
- Cheng, Q.; Sun, D.W. Factors affecting the water holding capacity of red meat products: A review of recent research advances. Crit. Rev. Food Sci. Nutr. 2008, 48, 137–159. [Google Scholar] [CrossRef]
- Aaslyng, M.D.; Hviid, M. Meat quality in the Danish pig population anno 2018. Meat Sci. 2020, 163, 108034. [Google Scholar] [CrossRef]
- Xu, D.; Wang, Y.; Jiao, N.; Qiu, K.; Zhang, X.; Wang, L.; Wang, L.; Yin, J. The coordination of dietary valine and isoleucine on water holding capacity, pH value and protein solubility of fresh meat in finishing pigs. Meat Sci. 2020, 163, 108074. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Chen, X.; Huang, Z.; Chen, D.; Li, M.; He, J. Effects of dietary grape seed proanthocyanidin extract supplementation on meat quality, muscle fiber characteristics and antioxidant capacity of finishing pigs. Food Chem. 2022, 367, 130781. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, J.; Shao, Y.; Yao, W.; Xia, J.; He, Q. The effect of dietary garcinol supplementation on oxidative stability, muscle postmortem glycolysis and meat quality in pigs. Meat Sci. 2020, 161, 107998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Luo, J.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X. Dietary resveratrol supplementation improves meat quality of finishing pigs through changing muscle fiber characteristics and antioxidative status. Meat Sci. 2015, 102, 15–21. [Google Scholar] [CrossRef]
- Coronado, S.A.; Trout, G.R.; Dunshea, F.R.; Shah, N.P. Antioxidant effects of rosemary extract and whey powder on the oxidative stability of wiener sausages during 10 months frozen storage. Meat Sci. 2002, 62, 217–224. [Google Scholar] [CrossRef]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the molecular structure and function relationship of β-glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P.; Yoon, I.; Zhen, Y.G. Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs. J. Anim. Sci. 2009, 87, 2614–2624. [Google Scholar] [CrossRef]
- Ryu, Y.C.; Kim, B.C. Comparison of histochemical characteristics in various pork groups categorized by postmortem metabolic rate and pork quality. J. Anim. Sci. 2006, 84, 894–901. [Google Scholar] [CrossRef]
- Matarneh, S.K.; England, E.M.; Scheffler, T.L.; Oliver, E.M.; Gerrard, D.E. Net lactate accumulation and low buffering capacity explain low ultimate pH in the longissimus lumborum of AMPKγ3 R200Q mutant pigs. Meat Sci. 2015, 110, 189–195. [Google Scholar] [CrossRef]
- Choe, J.H.; Choi, Y.M.; Lee, S.H.; Shin, H.G.; Ryu, Y.C.; Hong, K.C.; Kim, B.C. The relation between glycogen, lactate content and muscle fiber type composition, and their influence on postmortem glycolytic rate and pork quality. Meat Sci. 2008, 80, 355–362. [Google Scholar] [CrossRef]
- Luo, J.; Shen, Y.L.; Lei, G.H.; Zhu, P.K.; Jiang, Z.Y.; Bai, L.; Li, Z.M.; Tang, Q.G.; Li, W.X.; Zhang, H.S.; et al. Correlation between three glycometabolic-related hormones and muscle glycolysis, as well as meat quality, in three pig breeds. J. Sci. Food Agric. 2017, 97, 2706–2713. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Shan, T.; Wu, T.; Zhu, L.N.; Ren, Y.; An, S.; Wang, Y. Comparisons of different muscle metabolic enzymes and muscle fiber types in Jinhua and Landrace pigs. J. Anim. Sci. 2011, 89, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, S.T.; Kim, G.D.; Hwang, Y.H.; Ryu, Y.C. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Sci. 2013, 95, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Reggiani, C. Myosin isoforms in mammalian skeletal muscle. J. Appl. Physiol. (1985) 1994, 77, 493–501. [Google Scholar] [CrossRef]
- Du, M.; Shen, Q.W.; Zhu, M.J. Role of beta-adrenoceptor signaling and AMP-activated protein kinase in glycolysis of postmortem skeletal muscle. J. Agric. Food Chem. 2005, 53, 3235–3239. [Google Scholar] [CrossRef]
- Shen, Q.W.; Means, W.J.; Thompson, S.A.; Underwood, K.R.; Zhu, M.J.; McCormick, R.J.; Ford, S.P.; Du, M. Pre-slaughter transport, AMP-activated protein kinase, glycolysis, and quality of pork loin. Meat Sci. 2006, 74, 388–395. [Google Scholar] [CrossRef]
- Cain, D.W.; Cidlowski, J.A. Immune regulation by glucocorticoids. Nat. Rev. Immunol. 2017, 17, 233–247. [Google Scholar] [CrossRef]
- Yang, H.; Xia, L.; Chen, J.; Zhang, S.; Martin, V.; Li, Q.; Lin, S.; Chen, J.; Calmette, J.; Lu, M.; et al. Stress-glucocorticoid-TSC22D3 axis compromises therapy-induced antitumor immunity. Nat. Med. 2019, 25, 1428–1441. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Suzuki, K.; Coombes, J.S. The influence of antioxidant supplementation on markers of inflammation and the relationship to oxidative stress after exercise. J. Nutr. Biochem. 2007, 18, 357–371. [Google Scholar] [CrossRef]
- Stentz, F.B.; Umpierrez, G.E.; Cuervo, R.; Kitabchi, A.E. Proinflammatory cytokines, markers of cardiovascular risks, oxidative stress, and lipid peroxidation in patients with hyperglycemic crises. Diabetes 2004, 53, 2079–2086. [Google Scholar] [CrossRef] [Green Version]
- McAdam, B.F.; Mardini, I.A.; Habib, A.; Burke, A.; Lawson, J.A.; Kapoor, S. Effect of regulated expression of human cyclooxygenase isoforms on eicosanoid and isoeicosanoid production in inflammation. J. Clin. Investig. 2000, 105, 1473–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, V.R.; Marchini, J.S.; Cortiella, J. Assessment of protein nutritional status. J. Nutr. 1990, 120, 1496–1502. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M. Muscle wasting: The gut microbiota as a new therapeutic target? Int. J. Biochem. Cell. Biol. 2013, 45, 2186–2190. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Williams, T.J.; Montgomery, K.; Wong, H.L.; Zaugg, J.; Berengut, J.F.; Bissett, A.; Chuvochina, M.; Hugenholtz, P.; Ferrari, B.C. Candidatus Eremiobacterota, a metabolically and phylogenetically diverse terrestrial phylum with acid-tolerant adaptations. ISME J. 2021, 15, 2692–2707. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Content, % | Nutrient Levels | Content |
|---|---|---|---|
| Corn | 81.00 | Analyzed nutrient levels | |
| Soybean meal | 11.50 | Crude protein | 12.37 |
| Wheat bran | 3.20 | Lysine | 0.84 |
| Soybean oil | 1.20 | Methionine + cysteine | 0.44 |
| L-Lysine·HCl, % | 0.39 | Threonine | 0.53 |
| DL-Methionine, 98.0% | 0.03 | Tryptophan | 0.14 |
| L-Threonine, 98.5% | 0.11 | Isoleucine | 0.45 |
| L-Tryptophan, % | 0.03 | Leucine | 1.13 |
| L-Valine, % | 0.02 | Valine | 0.59 |
| Limestone | 0.70 | Calculated nutrient levels | |
| Dicalcium phosphate | 0.90 | Digestible energy, MJ/kg | 14.37 |
| Salt | 0.34 | Metabolizable energy, MJ/kg | 14.01 |
| 50% Choline chloride | 0.08 | Standardized ileal digestible amino acids | |
| Premix 1 | 0.50 | Lysine | 0.75 |
| Total | 100.00 | Methionine + cysteine | 0.42 |
| Threonine | 0.46 | ||
| Tryptophan | 0.13 | ||
| Isoleucine | 0.40 | ||
| Leucine | 1.03 | ||
| Valine | 0.48 | ||
| Items | Yeast β-Glucan Levels (mg/kg) | SEM | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | ANOVA | Linear | Quadratic | ||
| Albumin, g/L | 32.50 b | 32.43 b | 33.61 ab | 34.95 a | 35.34 a | 0.63 | <0.01 | <0.01 | 0.05 |
| Globulin, g/L | 36.86 b | 39.22 ab | 39.75 ab | 41.30 a | 37.18 b | 1.15 | 0.05 | 0.83 | <0.01 |
| Totel protein, g/L | 72.20 | 74.76 | 73.36 | 73.73 | 71.23 | 1.31 | 0.37 | 0.25 | 0.21 |
| A/G | 0.97 a | 0.90 ab | 0.86 ab | 0.79 b | 0.88 ab | 0.03 | 0.02 | 0.01 | <0.01 |
| Cortisol, ng/mL | 145.10 a | 122.08 b | 127.10 ab | 122.17 b | 136.08 ab | 6.19 | 0.04 | 0.92 | 0.01 |
| Items | Yeast β-Glucan Levels (mg/kg) | SEM | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | ANOVA | Linear | Quadratic | ||
| Live body weight, kg | 110.11 | 112.28 | 111.16 | 110.44 | 110.37 | 1.53 | 0.86 | 0.70 | 0.80 |
| Carcass weight, kg | 83.37 | 84.23 | 82.61 | 82.00 | 81.08 | 0.93 | 0.17 | 0.02 | 0.79 |
| Carcass length, cm | 80.70 | 78.76 | 79.38 | 79.29 | 79.13 | 0.77 | 0.47 | 0.41 | 0.40 |
| Dressing percentage, % | 74.20 | 74.97 | 74.35 | 74.36 | 74.58 | 0.33 | 0.50 | 0.86 | 0.94 |
| Back fat depth, mm | |||||||||
| Shoulder fat thickness | 39.77 | 39.58 | 43.22 | 38.70 | 40.68 | 1.11 | 0.06 | 0.96 | 0.93 |
| Last rib fat thickness | 22.06 | 23.40 | 25.27 | 24.26 | 23.50 | 0.78 | 0.08 | 0.48 | 0.02 |
| Lumbosacral fat thickness | 17.49 | 17.27 | 18.10 | 16.94 | 16.59 | 0.73 | 0.65 | 0.27 | 0.77 |
| 6th to 7th rib fat thickness | 28.27 | 28.74 | 30.99 | 26.46 | 28.25 | 1.36 | 0.24 | 0.52 | 0.84 |
| 10th rib fat thickness | 20.33 ab | 20.12 b | 23.03 ab | 20.39 ab | 23.51 a | 0.79 | 0.01 | 0.01 | 0.67 |
| Average back-fat depth | 26.58 | 26.36 | 29.51 | 25.80 | 26.67 | 1.10 | 0.95 | 0.69 | 0.69 |
| Loin eye area, cm2 | 43.56 | 45.24 | 42.24 | 39.78 | 42.24 | 1.88 | 0.36 | 0.30 | 0.21 |
| Fat-free lean index, % | 49.10 a | 48.77 ab | 47.71 b | 48.68 ab | 48.33 ab | 0.32 | 0.04 | 0.27 | 0.20 |
| Items | Yeast β-Glucan Levels (mg/kg) | SEM | p Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 50 | 100 | 200 | 400 | ANOVA | Linear | Quadratic | ||
| Meat quality | |||||||||
| pH45 min | 6.16 b | 6.23 b | 6.34 ab | 6.56 a | 6.42 ab | 0.07 | <0.01 | <0.01 | <0.01 |
| pH24 h | 5.61 | 5.63 | 5.61 | 5.65 | 5.61 | 0.03 | 0.90 | 0.98 | 0.49 |
| Flesh color score | 1.69 | 1.84 | 1.80 | 2.07 | 1.93 | 0.11 | 0.15 | 0.10 | 0.09 |
| L*45 min | 44.28 | 42.50 | 42.61 | 41.01 | 40.81 | 1.03 | 0.14 | 0.02 | 0.25 |
| a*45 min | 14.01 ab | 14.12 ab | 13.83 b | 14.68 a | 14.66 a | 0.24 | 0.05 | 0.01 | 0.63 |
| b*45 min | 10.00 | 9.55 | 9.45 | 9.16 | 9.49 | 0.34 | 0.54 | 0.38 | 0.14 |
| Drip loss, % | 1.62 a | 1.27 ab | 1.33 ab | 0.86 b | 1.26 ab | 0.15 | 0.03 | 0.11 | 0.01 |
| Cooking loss, % | 26.75 ab | 26.55 ab | 30.20 a | 24.57 b | 24.53 b | 0.95 | <0.01 | 0.01 | 0.50 |
| Shear force, n | 71.07 a | 60.74 b | 62.94 ab | 67.32 ab | 67.64 ab | 2.62 | 0.06 | 0.64 | 0.20 |
| Marbling score | 1.66 | 1.78 | 1.79 | 1.94 | 1.69 | 0.09 | 0.28 | 0.93 | 0.03 |
| Proximate analysis | |||||||||
| Moisture, % | 73.21 | 74.01 | 73.22 | 73.30 | 73.27 | 0.43 | 0.64 | 0.66 | 0.98 |
| Crude protein, % | 23.17 | 22.75 | 22.70 | 23.05 | 22.83 | 0.28 | 0.72 | 0.77 | 0.78 |
| Intramuscular fat, % | 4.00 | 3.50 | 3.88 | 4.05 | 3.69 | 0.24 | 0.48 | 0.80 | 0.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Guo, J.; Wang, Y.; Wang, L.; Xu, D.; Yan, E.; Zhang, X.; Yin, J. Effects of Dietary Yeast β-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs. Antioxidants 2022, 11, 1340. https://doi.org/10.3390/antiox11071340
He L, Guo J, Wang Y, Wang L, Xu D, Yan E, Zhang X, Yin J. Effects of Dietary Yeast β-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs. Antioxidants. 2022; 11(7):1340. https://doi.org/10.3390/antiox11071340
Chicago/Turabian StyleHe, Linjuan, Jianxin Guo, Yubo Wang, Lu Wang, Doudou Xu, Enfa Yan, Xin Zhang, and Jingdong Yin. 2022. "Effects of Dietary Yeast β-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs" Antioxidants 11, no. 7: 1340. https://doi.org/10.3390/antiox11071340
APA StyleHe, L., Guo, J., Wang, Y., Wang, L., Xu, D., Yan, E., Zhang, X., & Yin, J. (2022). Effects of Dietary Yeast β-Glucan Supplementation on Meat Quality, Antioxidant Capacity and Gut Microbiota of Finishing Pigs. Antioxidants, 11(7), 1340. https://doi.org/10.3390/antiox11071340