
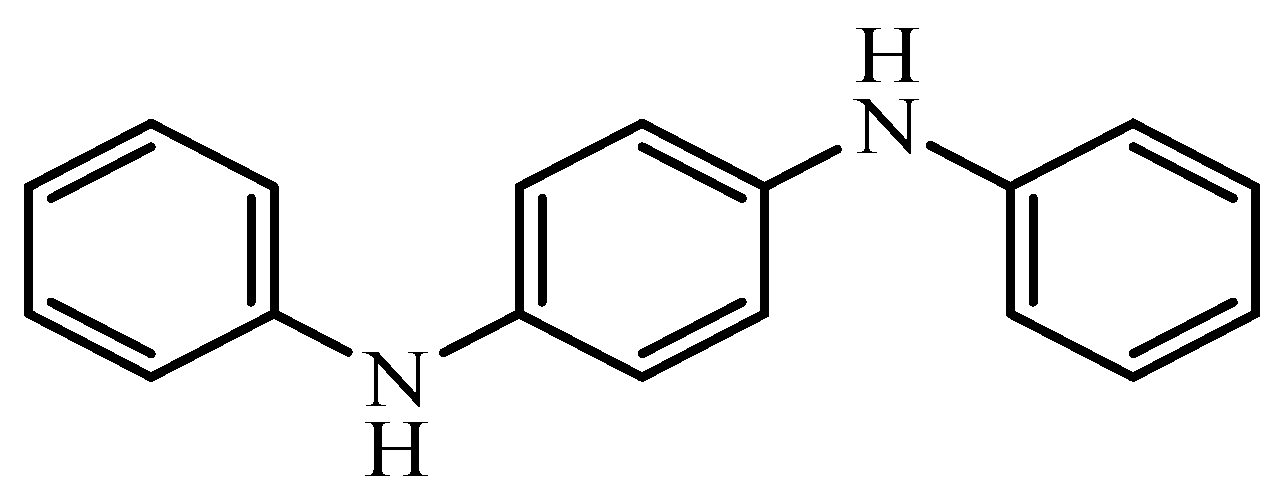
N,N′-Diphenyl-1,4-phenylenediamine Antioxidant’s Potential Role in Enhancing the Pancreatic Antioxidant, Immunomodulatory, and Anti-Apoptotic Therapeutic Capabilities of Adipose-Derived Stem Cells in Type I Diabetic Rats

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. AD-MSCs Isolation and Preparation
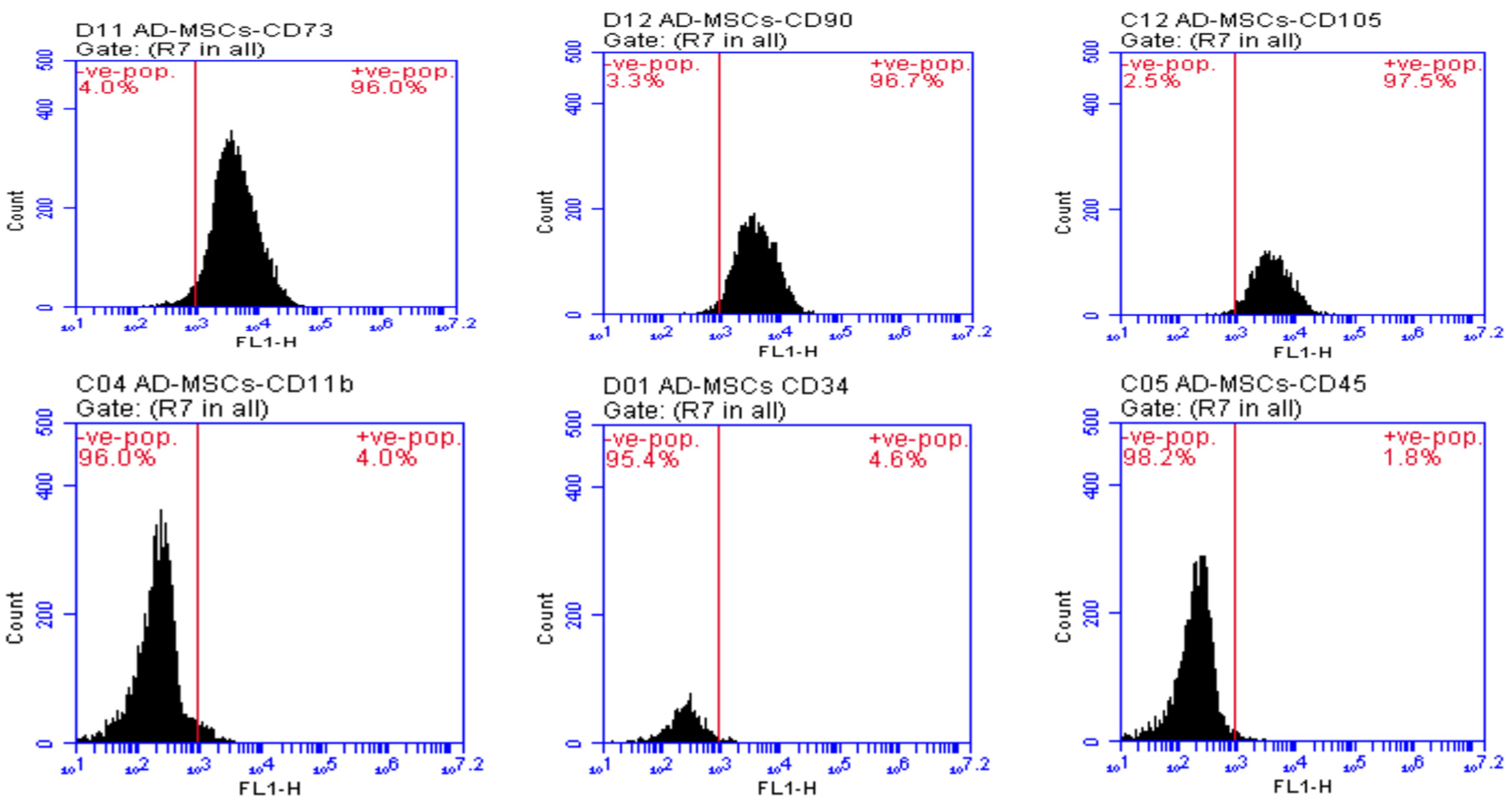
2.3. AD-MSCs Characterization
2.4. AD-MSCs Counting
2.5. Animals’ Maintenance and Grouping
- Control group: received an intraperitoneal (i.p.) sodium citrate (Na3C6H5O7) buffer (pH 4.5, single dose) once.
- DPPD group: one-time injection with DPPD (250 mg/kg, i.p., single dose) daily for four weeks.
- Diabetic (D) untreated group: one-time injection with STZ (45 mg/kg, i.p., single dose) dissolved in Na3C6H5O7 buffer (pH 4.5).
- Diabetic DPPD treated group: Injected with DPPD (250 mg/kg, i.p., single dose) daily for four weeks, following diabetes induction confirmation.
- Diabetic AD-MSCs treated group: Injected once with an intravenous (i.v., single dose) of AD-MSCs (1 × 106 cell/rat), following diabetes induction confirmation.
- Diabetic AD-MSCs+DPPD treated group: Injected once at day one with AD-MSCs (1 × 106 cell/rat, i.v., single dose) following diabetes induction confirmation, then treated with DPPD (250 mg/kg, i.p., single dose) daily for four weeks.
2.6. Samples Collection
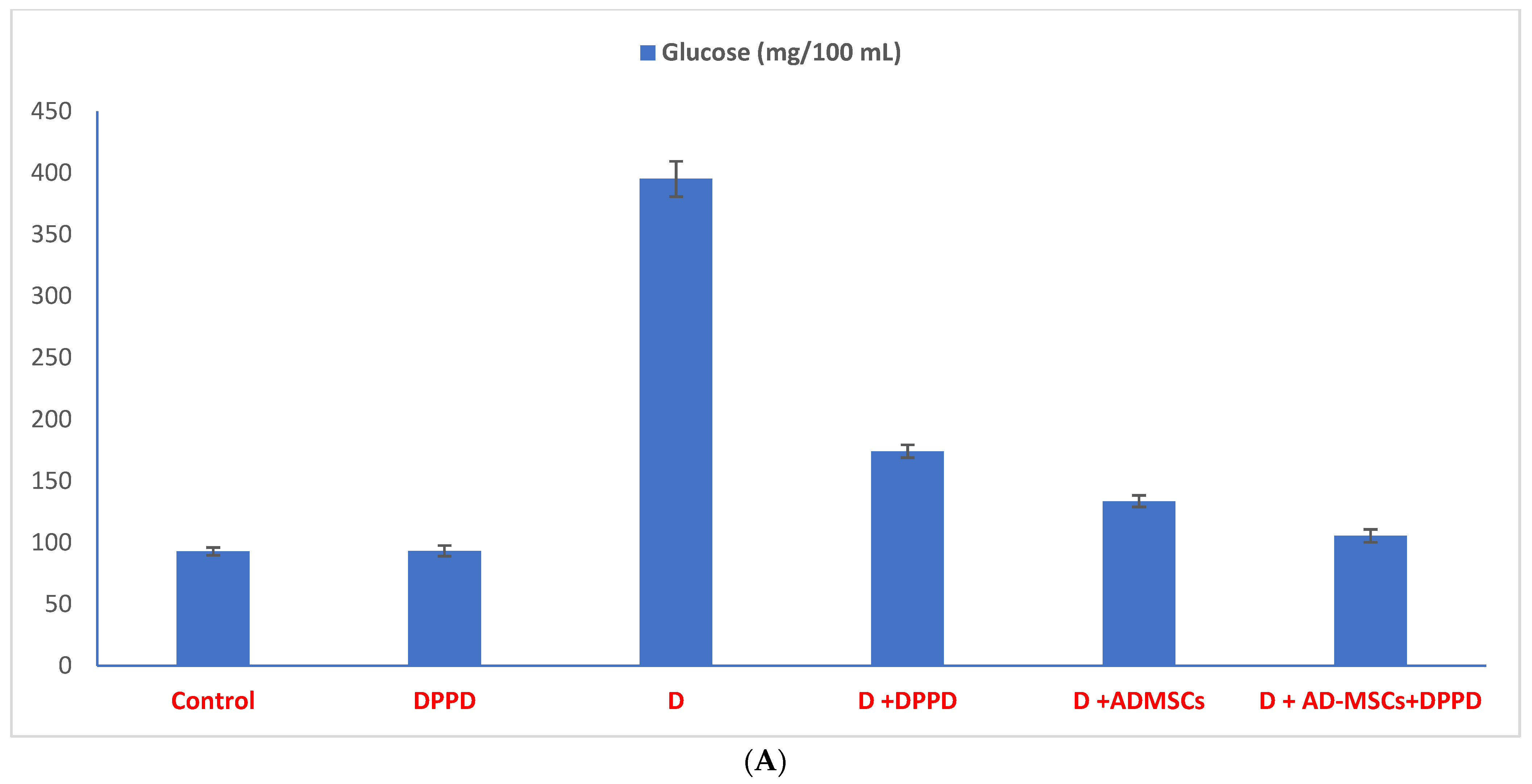
2.7. Biochemical Assays
2.8. Preparation and Staining of Pancreatic Cells for Flow Cytometric Analysis
2.9. Statistical Analysis
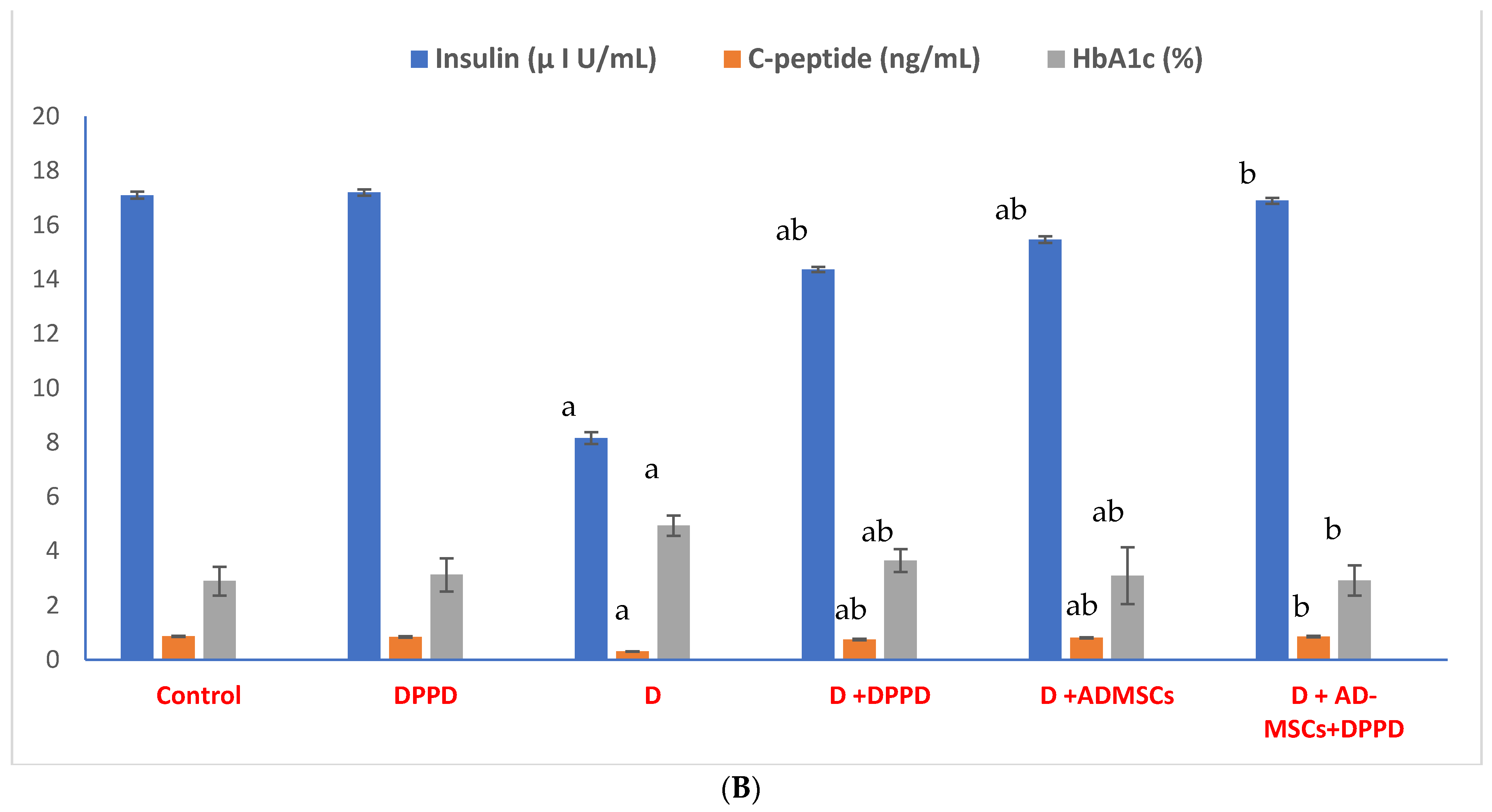
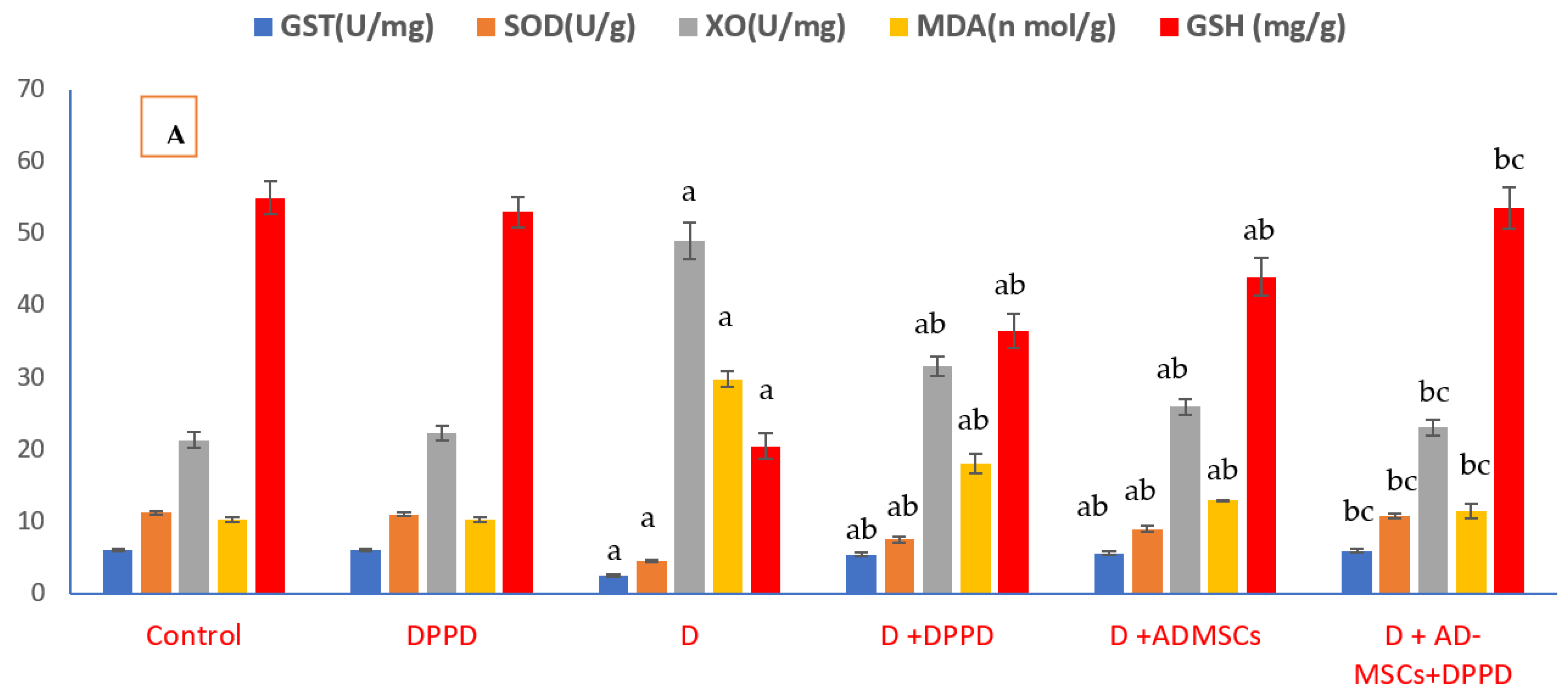
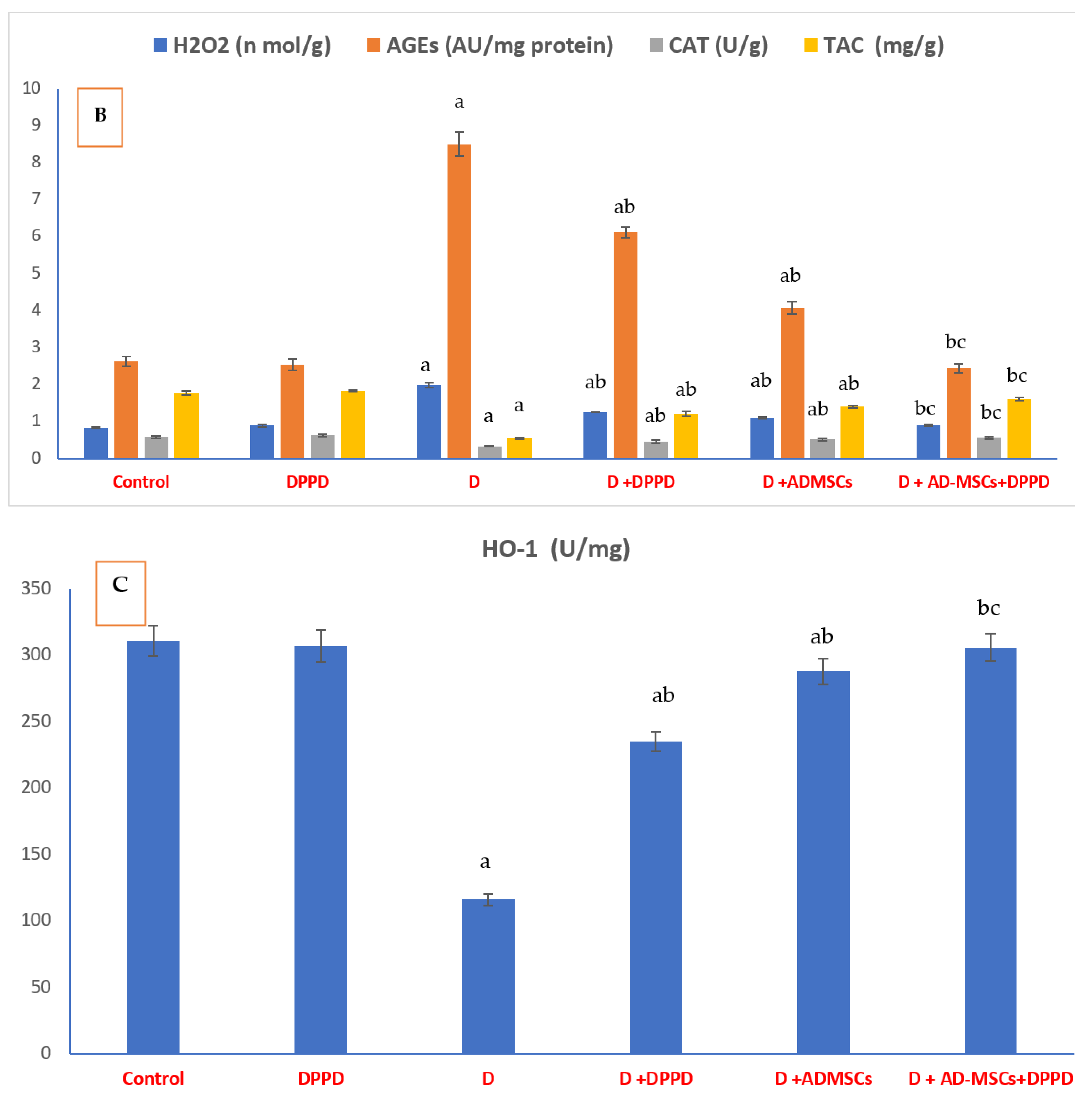
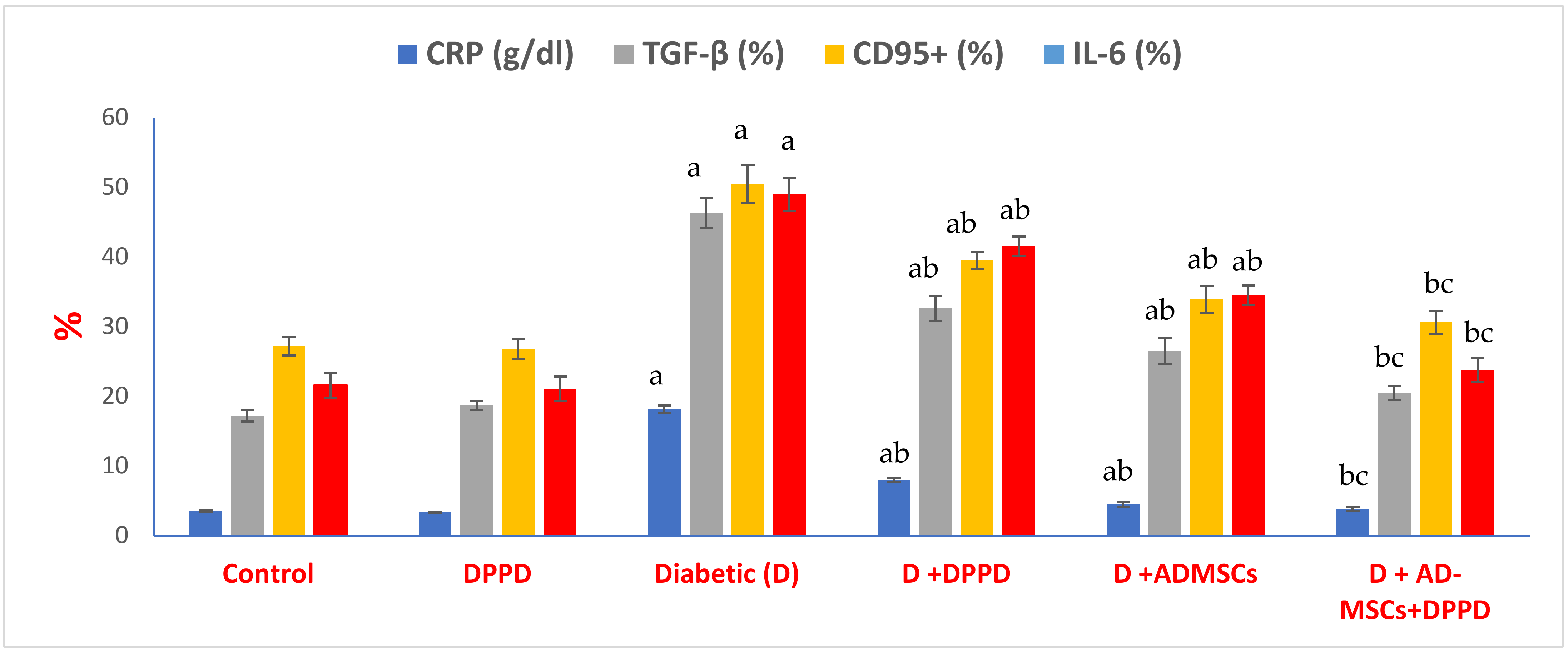
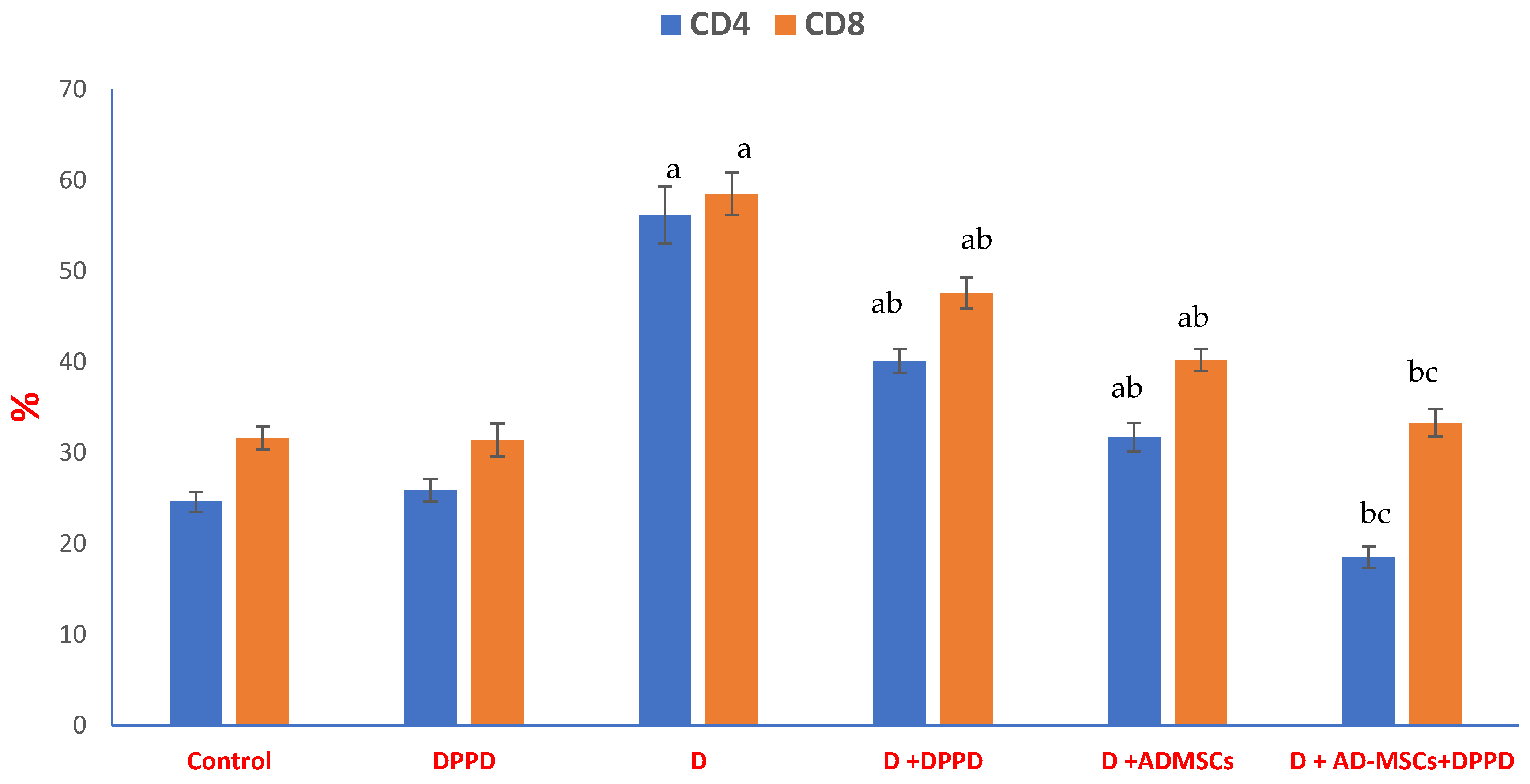
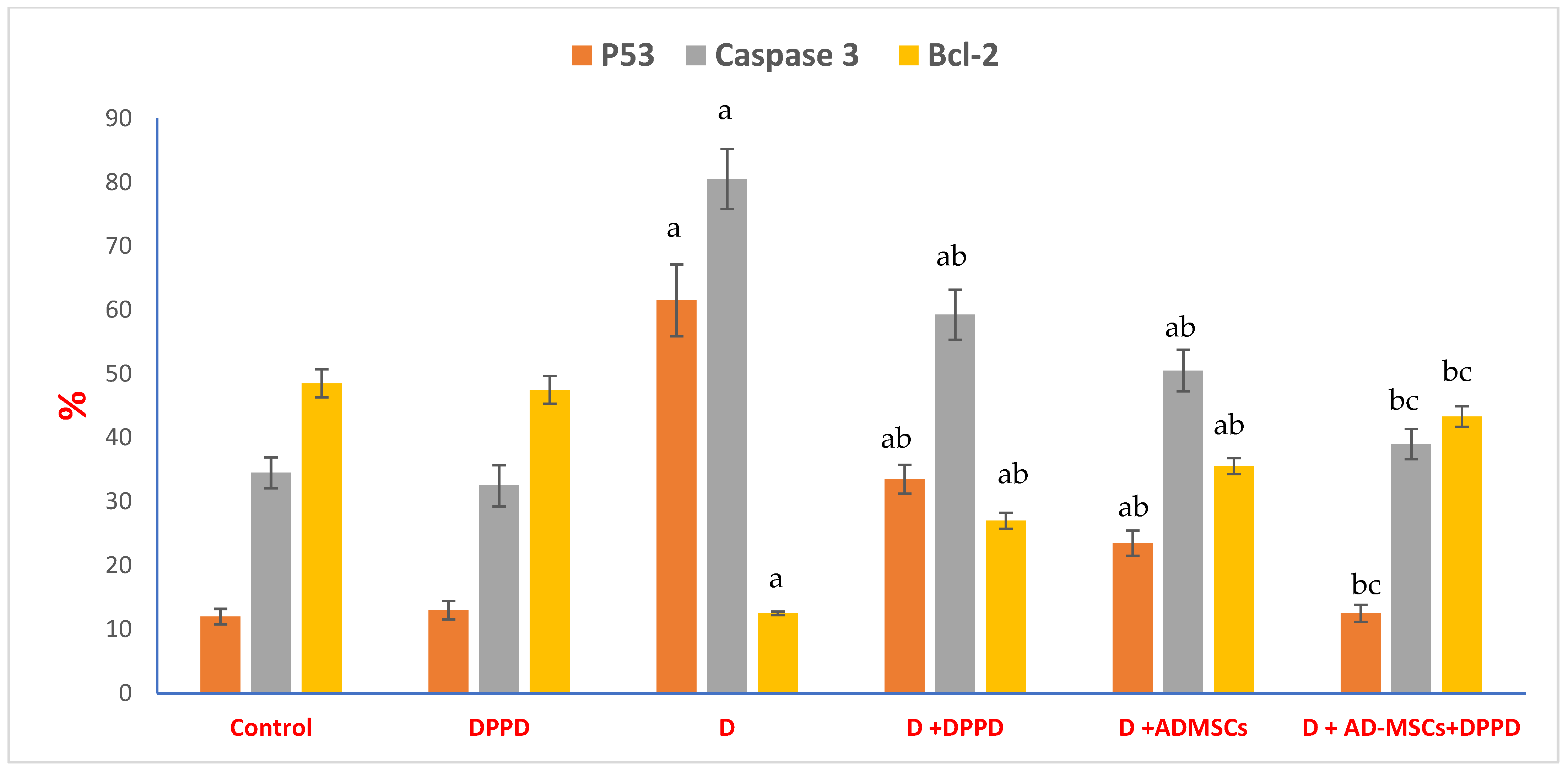
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Salpea, P.; Karuranga, S.; Petersohn, I.; Malanda, B.; Gregg, E.W.; Unwin, N.; Wild, S.H.; Williams, R. Mortality attributable to diabetes in 20–79 years old adults, 2019 estimates: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2020, 162, 108086. [Google Scholar] [CrossRef] [Green Version]
- Miah, R.; Hannan, M.A.; Rahman, A.S.; Khan, S.; Hossain, M.; Rahman, I.T.; Hossain, S.; Shahriar, C.S.; Uddin, M.B.; Talukdar, M.T.H.; et al. Processed Radio Frequency towards Pancreas Enhancing the Deadly Diabetes Worldwide. J. Endocrinol. Res. 2021, 3, 2826. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Tian, Y.; Huang, W.; Tong, N.; Fu, X. Integrative biology of extracellular vesicles in diabetes mellitus and diabetic complications. Theranostics 2022, 12, 1342–1372. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.S.; Lai, P.F.; Kuo, C.H.; Day, C.H.; Chen, R.J.; Ho, T.J.; Yeh, Y.L.; Mahalakshmi, B.; Padmaviswanadha, V.; Kuo, W.W. Resveratrol enhances therapeutic effect on pancreatic regeneration in diabetes mellitus rats receiving autologous transplantation of adipose-derived stem cells. Chin. J. Physiol. 2020, 63, 122–127. [Google Scholar] [PubMed]
- Amer, M.G.; Embaby, A.S.; Karam, R.A.; Amer, M.A. Role of adipose tissue derived stem cells differentiated into insulin producing cells in the treatment of type I diabetes mellitus. Gene 2018, 654, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Liu, D.D.; Thakor, A.S. Mesenchymal Stromal Cell Homing: Mechanisms and Strategies for Improvement. iScience 2019, 15, 421–438. [Google Scholar] [CrossRef] [Green Version]
- Bagno, L.; Hatzistergos, K.E.; Balkan, W.; Hare, J.M. Mesenchymal Stem Cell-Based Therapy for Cardiovascular Disease: Progress and Challenges. Mol. Ther. 2018, 26, 1610–1623. [Google Scholar] [CrossRef] [Green Version]
- El-Sawah, S.G.; Al thobaiti, F.; Aldahahrani, A.; Fayad, E.; Amen, R.M.; Shabana, E.; El-Hallous Rashwan, H.M. Investigation of the antioxidant defensive role of both AD-MSCs and BM-MSCs in modulating the alteration in the oxidative stress status in various STZ-diabetic rats’ tissues. Biocell 2021, 45, 1561–1568. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B., 3rd. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar] [CrossRef]
- Yang, X.; Li, C.J.; Wan, Y.; Smith, P.; Shang, G.; Cui, Q. Antioxidative fullerol promotes osteogenesis of human adipose-derived stem cells. Int. J. Nanomed. 2015, 9, 4023–4031. [Google Scholar] [CrossRef]
- Domingues, C.C.; Kundu, N.; Kropotova, Y.; Ahmadi, N.; Sen, S. Antioxidant-upregulated mesenchymal stem cells reduce inflammation and improve fatty liver disease in diet-induced obesity. Stem Cell Res. Ther. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornienko, J.S.; Smirnova, I.S.; Pugovkina, N.A.; Ivanova, J.S.; Shilina, M.A.; Grinchuk, T.M.; Shatrova, A.N.; Aksenov, N.D.; Zenin, V.V.; Nikolsky, N.N.; et al. High doses of synthetic antioxidants induce premature senescence in cultivated mesenchymal stem cells. Sci. Rep. 2019, 9, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.J.; Shults, N.V. Antioxidant Regulation of Cell Reprogramming. Antioxidants 2019, 8, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skubis-Sikora, A.; Sikora, B.; Witkowska, A.; Mazurek, U.; Gola, J. Osteogenesis of adipose-derived stem cells from patients with glucose metabolism disorders. Mol. Med. 2020, 26, 67. [Google Scholar] [CrossRef] [PubMed]
- Orciani, M.; Davis, S.; Appolloni, G.; Lazzarini, R.; Mattioli-Belmonte, M.; Ricciuti, R.A.; Boscaro, M.; Di Primio, R.; Arnaldi, G. Isolation and characterization of progenitor mesenchymal cells in human pituitary tumors. Cancer Gene Ther. 2015, 22, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Shaban, S.; El-Husseny, M.W.A.; Abushouk, A.I.; Salem, A.M.A.; Mamdouh, M.; Abdel-Daim, M.M. Effects of Antioxidant Supplements on the Survival and Differentiation of Stem Cells. Oxidative Med. Cell. Longev. 2017, 2017, 5032102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Feng, Q.; Ma, J.; Feng, Z.; Zhan, M.; Ouyang, L.; Mu, S.; Liu, B.; Jiang, Z.; Jia, Y.; et al. Melatonin Ameliorates Injury and Specific Responses of Ischemic Striatal Neurons in Rats. J. Histochem. Cytochem. 2013, 61, 591–605. [Google Scholar] [CrossRef]
- Tang, Y.; Cai, B.; Yuan, F.; He, X.; Lin, X.; Wang, J.; Wang, Y.; Yang, G.-Y. Melatonin Pretreatment Improves the Survival and Function of Transplanted Mesenchymal Stem Cells after Focal Cerebral Ischemia. Cell Transplant. 2014, 23, 1279–1291. [Google Scholar] [CrossRef]
- Kadry, S.M.; El-Dakdoky, M.H.; Rashid, L.; Zakaria, N.; Tarek, M. The potential role of melatonin on bone marrow mesenchymal stem cells therapy in pancreatic tissue of streptozotocin-induced diabetic rats. J. Sci. Res. Sci. 2017, 34, 85–107. [Google Scholar] [CrossRef]
- Amin, S.N.; Sharawy, N.; El Tablawy, N.; Elberry, D.A.; Youssef, M.F.; Abdelhady, E.G.; Rashed, L.A.; Hassan, S.S. Melatonin-Pretreated Mesenchymal Stem Cells Improved Cognition in a Diabetic Murine Model. Front. Physiol. 2021, 12, 628107. [Google Scholar] [CrossRef]
- Refat, M.S.; Hamza, R.Z.; Adam, A.M.A.; Saad, H.A.; Gobouri, A.A.; Al-Harbi, F.S.; Al-Salmi, F.A.; Altalhi, T.; El-Megharbel, S.M. Quercetin/Zinc complex and stem cells: A new drug therapy to ameliorate glycometabolic control and pulmonary dysfunction in diabetes mellitus: Structural characterization and genetic studies. PLoS ONE 2021, 16, e0246265. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Satoh, T.; Hibi, D.; Ohno, Y.; Kohda, Y.; Miura, K.; Gemba, M. The Effect of Antioxidant on Development of Fibrosis by Cisplatin in Rats. J. Pharmacol. Sci. 2009, 111, 433–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omar, H.M.; Ahmed, E.; Sarry, A.; Mohammed, S.; Nasser, A.Y. Hepatoprotective effects of vitamin C, DPPD and L- cysteine against cisplatin-induced oxidative stress in male rats. J. Biol. Earth Sci. 2012, 2, 28–36. [Google Scholar]
- Nabil, A.; Elshemy, M.M.; Asem, M.; Gomaa, H.F. Protective Effect of DPPD on Mercury Chloride-Induced Hepatorenal Toxicity in Rats. J. Toxicol. 2020, 2020, 4127284. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shi, S.; Bai, R.; Wang, Y.; Guo, Z.; Li, D. Biological properties of bone marrow stem cells and adipose-derived stem cells derived from T2DM rats: A comparative study. Cell Biosci. 2020, 10, 102. [Google Scholar] [CrossRef]
- Zahran, F.; El Ghareb, M.; Nabil, A. Bone marrow derived mesenchymal stem cells as a therapy for renal injury. Int. J. Adv. Res. 2014, 4, 11–16. [Google Scholar] [CrossRef]
- Kodidela, S.; BegumShaik, F.; Chinta, V.; AliMohammad, S.; Pasala, C.; Mittameedi, S.; Maddu, N.; Wudayagiri, R.; Nallanchakravarthula, V. Possible ameliorative role of green tea on chronic alcohol mediated renal toxicity of STZ -induced diabetic rats. Clin. Nutr. Exp. 2020, 34, 1–25. [Google Scholar] [CrossRef]
- Tribukait, B.; Moberger, G.; Zetterberg, A. Methodological Aspects of Rapid-Flow Cytofluorometry for DNA Analysis of Human Urinary Bladder Cells; European Press: Burlington, MA, USA, 1975; pp. 50–60. [Google Scholar]
- Dang, L.; Phan, N.; Truong, K. Mesenchymal stem cells for diabetes mellitus treatment: New advances. Biomed. Res. Ther. 2017, 4, 1062–1081. [Google Scholar] [CrossRef] [Green Version]
- Abdel Fattah, S.A.; Waly, H.; El-Enein, A.A.; Kamel, A.; Labib, H. Mesenchymal stem cells versus curcumin in enhancing the alterations in the cerebellar cortex of streptozocin-induced diabetic albino rats. The role of GFAP, PLC and α-synuclein. J. Chem. Neuroanat. 2020, 109, 101842. [Google Scholar] [CrossRef]
- Aminzadeh, A.; Maroof, N.T.; Mehrabani, M.; Juybari, K.P.; Sharifi, A.M. Investigating the Alterations of Oxidative Stress Status, Antioxidant Defense Mechanisms, MAP Kinase and Mitochondrial Apoptotic Pathway in Adipose-Derived Mesenchymal Stem Cells from STZ Diabetic Rats. Cell J. 2020, 22, 38–48. [Google Scholar]
- Ezquer, M. Mesenchymal Stem Cell Therapy in Type 1 Diabetes Mellitus and Its Main Complications: From Experimental Findings to Clinical Practice. J. Stem Cell Res. Ther. 2014, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Song, L.; Strange, C.; Dong, X.; Wang, H. Therapeutic effects of adipose stem cells from diabetic mice for the treatment of type 2 diabetes. Mol. Ther. 2018, 26, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Fu, Z.; Chen, Y.; Tang, N.; Wang, L.; Wang, F. Effects of autologous adipose-derived stem cell infusion on type 2 diabetic rats. Endocr. J. 2015, 62, 339–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Atoui, R.; Chiu, R.C. Concise review: Immunomodulatory properties of mesenchymal stem cells in cellular transplantation: Update, controversies, and unknowns. Stem Cells Transl. Med. 2012, 1, 200–205. [Google Scholar] [CrossRef]
- Xie, Z.; Hao, H.; Tong, C.; Cheng, Y.; Liu, J.; Pang, Y.; Si, Y.; Guo, Y.; Zang, L.; Mu, Y. Human umbilical cord-derived mesenchymal stem cells elicit macrophages into an anti-inflammatory phenotype to alleviate insulin resistance in type 2 diabetic rats. Stem Cells 2016, 34, 627–639. [Google Scholar] [CrossRef]
- Takahashi, H.; Sakata, N.; Yoshimatsu, G.; Hasegawa, S.; Kodama, S. Regenerative and Transplantation Medicine: Cellular Therapy Using Adipose Tissue-Derived Mesenchymal Stromal Cells for Type 1 Diabetes Mellitus. J. Clin. Med. 2019, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Van Pham, P.; Vu, N.B.; Phan, N.K. Umbilical cord-derived stem cells (MODULATISTTM) show strong immunomodulation capacity compared to adipose tissue-derived or bone marrow-derived mesenchymal stem cells. Biomed. Res. Ther. 2016, 3, 687–696. [Google Scholar] [CrossRef]
- Omar, H.M.; Ahmed, E.; Sarry, A. The antioxidant activity of vitamin C, DPPD and L-cysteine against cisplatin-induced testicular oxidative stress in rat. Food Cosmet. Toxicol. 2011, 49, 1115–1121. [Google Scholar]
- Zahran, F.; Nabil, A.; Karef, A.E.; Lotfy, A.; Mahmoud, K.; Hozayen, W.G.; Sobh, M. Effect of Antioxidants and Mesenchymal Stem Cells on Cisplatin Induced Renal Fibrosis in Rats. J. Stem Cell Res. Ther. 2016, 1, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Kadry, S.M.; El-Dakdoky, M.H.; Haggag, N.Z.; Rashed, L.A.; Hassen, M.T. Melatonin improves the therapeutic role of mesenchymal stem cells in diabetic rats. Toxicol. Mech. Methods 2018, 28, 529–538. [Google Scholar] [CrossRef] [PubMed]
- ShamsEldeen, A.M.; Ashour, H.; Shoukry, H.S.; Fadel, M.; Kamar, S.S.; Aabdelbaset, M.; Rashed, L.A.; Ammar, H.I. Combined treatment with systemic resveratrol and resveratrol preconditioned mesenchymal stem cells, maximizes antifibrotic action in diabetic cardiomyopathy. J. Cell. Physiol. 2018, 234, 10942–10963. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ju, D.; Day, C.; Yeh, Y.; Chen, R.; Viswanadha, V.P.; Chang, R.; Lin, Y.; Yao, C.; Huang, C. Protective effect of autologous transplantation of resveratrol preconditioned adipose-derived stem cells in the treatment of diabetic liver dysfunction in rat model. J. Tissue Eng. Regen. Ther. 2019, 13, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Kuo, C.; Day, C.H.; Pan, L.; Chen, R.; Chen, B.; Padma, V.V.; Lin, Y.; Huang, C. Resveratrol increases stem cell function in the treatment of damaged pancreas. J. Cell. Physiol. 2019, 234, 20443–20452. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | DPPD | Diabetic (D) | D+DPPD | D+AD-MSCs | D+AD- MSCs+DPPD | ||
|---|---|---|---|---|---|---|---|
| G0/G1 phase (%) | Mean ± SEM | 91.70 ± 3.16 | 92.30 ± 3.37 | 57.50 ± 1.22 a | 75.50 ± 2.43 ab | 85.00 ± 2.63 ab | 93.00 ± 3.25 bc |
| S Phase (%) | Mean ± SEM | 2.50 ± 0.01 | 2.70 ± 0.02 | 11.00 ± 0.03 a | 6.50 ± 0.02 ab | 1.00 ± 0.01 ab | 2.50± 0.03 bc |
| G2/M phase (%) | Mean ± SEM | 2.80 ± 0.02 | 2.00 ± 0.02 | 6.50 ± 0.01 a | 1.70 ± 0.03 ab | 3.00 ± 0.01 ab | 2.70 ± 0.02 bc |
| Control | DPPD | Diabetic (D) | D+DPPD | D+AD-MSCs | D+AD- MSCs+DPPD | ||
|---|---|---|---|---|---|---|---|
| Viable cells (%) | Mean ± SEM | 93.70 ± 3.16 | 92.50 ± 3.37 | 33.20 ± 1.22 a | 63.50 ± 2.43 ab | 72.90 ± 2.63 ab | 93.00 ± 2.25 bc |
| Early apoptosis (%) | Mean ± SEM | 0.50 ± 0.21 | 1.40 ± 0.45 | 3.50 ± 0.63 a | 15.00 ± 0.26 ab | 13.90 ± 0.98 ab | 0.60 ± 0.33 bc |
| Late apoptosis (%) | Mean ± SEM | 4.60 ± 1.42 | 4.80 ± 1.22 | 29.00 ± 2.71 a | 21.80 ± 2.93 ab | 12.50 ± 2.26 ab | 6.50 ± 1.37 bc |
| Necrosis (%) | Mean ± SEM | 1.50 ± 0.12 | 1.40 ± 0.16 | 35.70 ± 2.21 a | 0.20 ± 0.02 ab | 0.10 ± 0.01 ab | 0.30 ± 0.01 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaaban, S.; El-Shamy, H.; Gouda, M.; Darwish, M.K.; Abd El-Lateef, H.M.; Khalaf, M.M.; El-Hallous, E.I.; Radwan, K.H.; Rashwan, H.M.; El-Sawah, S.G. N,N′-Diphenyl-1,4-phenylenediamine Antioxidant’s Potential Role in Enhancing the Pancreatic Antioxidant, Immunomodulatory, and Anti-Apoptotic Therapeutic Capabilities of Adipose-Derived Stem Cells in Type I Diabetic Rats. Antioxidants 2023, 12, 58. https://doi.org/10.3390/antiox12010058
Shaaban S, El-Shamy H, Gouda M, Darwish MK, Abd El-Lateef HM, Khalaf MM, El-Hallous EI, Radwan KH, Rashwan HM, El-Sawah SG. N,N′-Diphenyl-1,4-phenylenediamine Antioxidant’s Potential Role in Enhancing the Pancreatic Antioxidant, Immunomodulatory, and Anti-Apoptotic Therapeutic Capabilities of Adipose-Derived Stem Cells in Type I Diabetic Rats. Antioxidants. 2023; 12(1):58. https://doi.org/10.3390/antiox12010058
Chicago/Turabian StyleShaaban, Saad, Hemdan El-Shamy, Mohamed Gouda, Marwa K. Darwish, Hany M. Abd El-Lateef, Mai M. Khalaf, Ehab I. El-Hallous, Kholoud H. Radwan, Hanan M. Rashwan, and Shady G. El-Sawah. 2023. "N,N′-Diphenyl-1,4-phenylenediamine Antioxidant’s Potential Role in Enhancing the Pancreatic Antioxidant, Immunomodulatory, and Anti-Apoptotic Therapeutic Capabilities of Adipose-Derived Stem Cells in Type I Diabetic Rats" Antioxidants 12, no. 1: 58. https://doi.org/10.3390/antiox12010058
APA StyleShaaban, S., El-Shamy, H., Gouda, M., Darwish, M. K., Abd El-Lateef, H. M., Khalaf, M. M., El-Hallous, E. I., Radwan, K. H., Rashwan, H. M., & El-Sawah, S. G. (2023). N,N′-Diphenyl-1,4-phenylenediamine Antioxidant’s Potential Role in Enhancing the Pancreatic Antioxidant, Immunomodulatory, and Anti-Apoptotic Therapeutic Capabilities of Adipose-Derived Stem Cells in Type I Diabetic Rats. Antioxidants, 12(1), 58. https://doi.org/10.3390/antiox12010058