
Schizochytrium spp. Dietary Supplementation Modulates Immune-Oxidative Transcriptional Signatures in Monocytes and Neutrophils of Dairy Goats




Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Trial
2.2. Blood Collection, Cells Isolation, RNA Extraction, and cDNA Synthesis
2.3. Primer Design and Real-Time Quantitative PCR
2.4. Statistics
3. Results
3.1. Feed Consumption and Animal Performance
3.2. Effect of Schizochytrium spp. on Anti- and Pro-Oxidant-Related Gene Expressions of Monocytes and Neutrophils

3.3. Effect of Schizochytrium spp. on Immune-Related Gene Expressions of Monocytes and Neutrophils

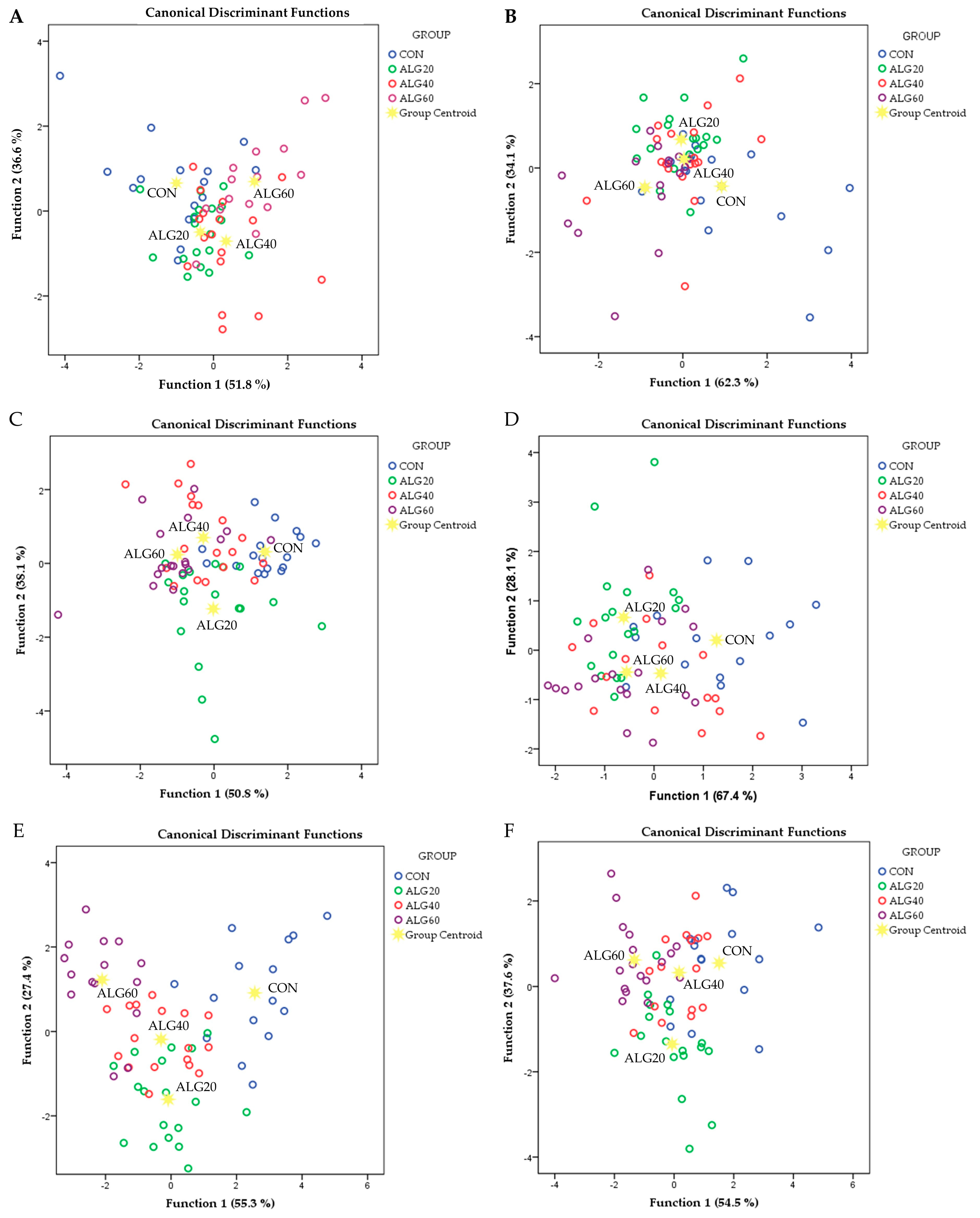
3.4. Holistic Statistical Analyses

4. Discussion
4.1. The Effect of Dietary Supplementation with Schizochytrium spp. on Pro- and Anti-Oxidant Gene Expressions
4.2. The Effect of Dietary Supplementation with Schizochytrium spp. on Immune-Related Gene Expressions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health Benefits throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Polyunsaturated Fatty Acids, Inflammation, and Immunity. Lipids 2001, 36, 1007–1024. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Wall, R.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. Health Implications of High Dietary Omega-6 Polyunsaturated Fatty Acids. J. Nutr. Metab. 2012, 2012, 539426. [Google Scholar] [CrossRef] [PubMed]
- Zisis, F.; Kyriakaki, P.; Satolias, F.F.; Mavrommatis, A.; Simitzis, P.E.; Pappas, A.C.; Surai, P.F.; Tsiplakou, E. The Effect of Dietary Inclusion of Microalgae Schizochytrium spp. on Ewes’ Milk Quality and Oxidative Status. Foods 2022, 11, 2950. [Google Scholar] [CrossRef] [PubMed]
- Mavrommatis, A.; Tsiplakou, E. The Impact of the Dietary Supplementation Level with Schizochytrium sp. on Milk Chemical Composition and Fatty Acid Profile, of Both Blood Plasma and Milk of Goats. Small Rumin. Res. 2020, 193, 106252. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Sotirakoglou, K.; Kamilaris, C.; Tsiplakou, E. Effects of Inclusion of Schizochytrium spp. and Forage-to-Concentrate Ratios on Goats’ Milk Quality and Oxidative Status. Foods 2021, 10, 1322. [Google Scholar] [CrossRef]
- di Nunzio, M.; Valli, V.; Bordoni, A. PUFA and Oxidative Stress. Differential Modulation of the Cell Response by DHA. Int. J. Food Sci. Nutr. 2016, 67, 834–843. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Chronopoulou, E.G.; Sotirakoglou, K.; Labrou, N.E.; Zervas, G.; Tsiplakou, E. The Impact of the Dietary Supplementation Level with Schizochytrium sp., on the Oxidative Capacity of Both Goats’ Organism and Milk. Livest. Sci. 2018, 218, 37–43. [Google Scholar] [CrossRef]
- Lingappan, K. NF-ΚB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Andrés Juan, C.; Plou, F.J.; Pérez-Lebeña, E. Superoxide Anion Chemistry—Its Role at the Core of the Innate Immunity. Int. J. Mol. Sci. 2023, 24, 1841. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Mavrommatis, A.; Theodorou, G.; Politis, I.; Tsiplakou, E. Schizochytrium sp. Dietary Supplementation Modify Toll-like Receptor 4 (TLR4) Transcriptional Regulation in Monocytes and Neutrophils of Dairy Goats. Cytokine 2021, 148, 155588. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, X.; Liu, S.; Shen, D.; Zhu, J.; Liu, K. Role of Resolvins in the Inflammatory Resolution of Neurological Diseases. Front. Pharmacol. 2020, 11, 612. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Dong, J.; Wu, W.; Yang, T.; Wang, T.; Guo, L.; Chen, L.; Xu, D.; Wen, F. Resolvin D1 Attenuates Inflammation in Lipopolysaccharide-Induced Acute Lung Injury through a Process Involving the PPARγ/NF-ΚB Pathway. Respir. Res. 2012, 13, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorachek, W.R.; Hugejiletu; Bobe, G.; Hall, J.A. Reference Gene Selection for Quantitative PCR Studies in Sheep Neutrophils. Int. J. Mol. Sci. 2013, 14, 11484–11495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoni, G.; Minuti, A.; Trevisi, E. Immune System, Inflammation and Nutrition in Dairy Cattle. Anim. Prod. Sci. 2015, 55, 943–948. [Google Scholar] [CrossRef]
- Samartín, S.; Chandra, R.K. Obesity, Overnutrition and the Immune System. Nutr. Res. 2001, 21, 243–262. [Google Scholar] [CrossRef]
- Lambertucci, R.H.; Hirabara, S.M.; Silveira, L.D.R.; Levada-Pires, A.C.; Curi, R.; Pithon-Curi, T.C. Palmitate Increases Superoxide Production through Mitochondrial Electron Transport Chain and NADPH Oxidase Activity in Skeletal Muscle Cells. J. Cell. Physiol. 2008, 216, 796–804. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell. Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Grotto, D.; Santa Maria, L.; Valentini, J.; Paniz, C.; Schmitt, G.; Garcia, S.C.; Pomblum, V.J.; Rocha, J.B.T.; Farina, M. Importance of the Lipid Peroxidation Biomarkers and Methodological Aspects for Malondialdehyde Quantification. Quim. Nova 2009, 32, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Manea, A.; Manea, S.A.; Todirita, A.; Albulescu, I.C.; Raicu, M.; Sasson, S.; Simionescu, M. High-Glucose-Increased Expression and Activation of NADPH Oxidase in Human Vascular Smooth Muscle Cells Is Mediated by 4-Hydroxynonenal-Activated PPARα and PPARβ/δ. Cell Tissue Res. 2015, 361, 593–604. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, S.H.; Lee, J.H.; Lee, B.H.; Yoon, H.J.; Shin, D.H.; Park, S.S.; Jang, S.B.; Park, J.S.; Jee, Y.K. Superoxide Dismutase Gene (SOD1, SOD2, and SOD3) Polymorphisms and Antituberculosis Drug-Induced Hepatitis. Allergy Asthma Immunol. Res. 2014, 7, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koussounadis, A.; Langdon, S.P.; Um, I.H.; Harrison, D.J.; Smith, V.A. Relationship between Differentially Expressed MRNA and MRNA-Protein Correlations in a Xenograft Model System. Sci. Rep. 2015, 5, 10775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelinová, V.; Mistrík, I.; Pavlovkin, J.; Tamás, L. Glutathione Peroxidase Expression and Activity in Barley Root Tip after Short-Term Treatment with Cadmium, Hydrogen Peroxide and t-Butyl Hydroperoxide. Protoplasma 2013, 250, 1057–1065. [Google Scholar] [CrossRef]
- Rudkowska, I.; Paradis, A.M.; Thifault, E.; Julien, P.; Tchernof, A.; Couture, P.; Lemieux, S.; Barbier, O.; Vohl, M.C. Transcriptomic and Metabolomic Signatures of an N-3 Polyunsaturated Fatty Acids Supplementation in a Normolipidemic/Normocholesterolemic Caucasian Population. J. Nutr. Biochem. 2013, 24, 54–61. [Google Scholar] [CrossRef]
- Gaffney, M.; O’Rourke, R.; Murphy, R. Manipulation of Fatty Acid and Antioxidant Profiles of the Microalgae Schizochytrium sp. through Flaxseed Oil Supplementation. Algal. Res. 2014, 6, 195–200. [Google Scholar] [CrossRef]
- Son, Y.; Cheong, Y.-K.; Kim, N.-H.; Chung, H.-T.; Kang, D.G.; Pae, H.-O. Mitogen-Activated Protein Kinases and Reactive Oxygen Species: How Can ROS Activate MAPK Pathways? J. Signal Transduct. 2011, 2011, 792639. [Google Scholar] [CrossRef] [Green Version]
- Smesny, S.; Milleit, B.; Hipler, U.C.; Milleit, C.; Schäfer, M.R.; Klier, C.M.; Holub, M.; Holzer, I.; Berger, G.E.; Otto, M.; et al. Omega-3 Fatty Acid Supplementation Changes Intracellular Phospholipase A 2 Activity and Membrane Fatty Acid Profiles in Individuals at Ultra-High Risk for Psychosis. Mol. Psychiatry 2014, 19, 317–324. [Google Scholar] [CrossRef]
- Orr, S.K.; Butler, K.L.; Hayden, D.; Tompkins, R.G.; Serhan, C.N.; Irimia, D. Gene Expression of Proresolving Lipid Mediator Pathways Is Associated with Clinical Outcomes in Trauma Patients. Crit. Care Med. 2015, 43, 2642–2650. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.Q.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-Inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radzikowska, U.; Rinaldi, A.O.; Sözener, Z.Ç.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The Influence of Dietary Fatty Acids on Immune Responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeb, L.; Hellen, P.; Boehnke, C.; Hoffmann, J.; Schuh-Hofer, S.; Dirnagl, U.; Reuter, U. IL-1β Stimulates COX-2 Dependent PGE2 Synthesis and CGRP Release in Rat Trigeminal Ganglia Cells. PLoS ONE 2011, 6, e17360. [Google Scholar] [CrossRef] [PubMed]
- Nauroth, J.M.; Liu, Y.C.; van Elswyk, M.; Bell, R.; Hall, E.B.; Chung, G.; Arterburn, L.M. Docosahexaenoic Acid (DHA) and Docosapentaenoic Acid (DPAn-6) Algal Oils Reduce Inflammatory Mediators in Human Peripheral Mononuclear Cells in Vitro and Paw Edema in Vivo. Lipids 2010, 45, 375–384. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Amplicon Size | Tm °C | GenBank RefSeq |
|---|---|---|---|---|
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | F: 5’-AAAGGCCATCACCATCTTCCA-3’ | 75 | 62 | XM_005680968.3 |
| R: 5’-ACCACGTACTCAGCACCAGCAT-3’ | ||||
| Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta (YWHAZ) | F: 5’-TGTTCTATTGTGCCTAGTACACTGT-3′ | 70 | 62 | XM_018058314.1 |
| R: 5’-CATCAAGACTCACTGCCTCCC-3′ | ||||
| Catalase (CAT) | F: 5′-GAGGAAACGCCTGTGTGAGA-3′ | 116 | 60 | XM_005690077.3 |
| R: 5′-GGATGCGGGAGCCATATTCA-3′ | ||||
| Superoxide dismutase 1 (SOD1) | F: 5′-ATCCACTTCGAGGCAAAGGG-3′ | 124 | 60 | NM_001285550.1 |
| R: 5′-CTGCACTGGTACAGCCTTGT-3′ | ||||
| Superoxide dismutase 2 (SOD2) | F: 5′-GCCCGATTATCTGAAGGCCA-3′ | 99 | 60 | XM_018053428.1 |
| R: 5′-CTCAGTGTAAGGCTGACGGT-3′ | ||||
| Superoxide dismutase 3 (SOD3) | F: 5′-CGAGTGTAAGGCCGTCTGAG-3′ | 76 | 60 | XM_018049136.1 |
| R: 5′-GGACATAGAAGGGGTCTGCG-3′ | ||||
| Glutathione peroxidase 1 (GPX1) | F: 5′-CATCGACATCGAGCCTGACA-3′ | 109 | 60 | XM_005695962.3 |
| R: 5′-AAAATCCCCGGAGAGCAGTG-3′ | ||||
| Glutathione peroxidase 2 (GPX2) | F: 5′-CCTCCCCACCCCTTTAATCG-3′ | 115 | 62 | XM_005685982.3 |
| R: 5′-GGCTGATAGCACTGAGGTCG-3′ | ||||
| Glutathione peroxidase 3 (GPX3) | F: 5′-GGAGGCCAAGGGGAAGTAAC-3′ | 114 | 60 | XM_005683183.3 |
| R: 5′-GCATGGGAGTGTGGCATAGT-3′ | ||||
| Glutathione transferase 1 (MGST1) | F: 5′-CTCCTGCTCGGATTCACACC-3′ | 88 | 60 | XM_018048426.1 |
| R: 5′-TTAGGTGCGAAAGGTTGACCA-3′ | ||||
| Glutathione transferase 2 (MGST2) | F: 5′-AAAGTTATGCCCCCATCCGT -3′ | 85 | 60 | XM_013970519.2 |
| R:5′-CACCAGACCCAGACAAGTAGC -3′ | ||||
| Glutathione transferase 3 (MGST3) | F: 5′-CCCCACTCTGATAGAGGCCA -3′ | 121 | 60 | XM_013975063.2 |
| R:5′-GTAGTCGTCCAGCCTCGTTT -3′ | ||||
| Glutathione reductase (GSR) | F: 5′-CTGCCCTGGGTTCTAAGACA-3′ | 104 | 60 | XM_018041989.1 |
| R:5′-AGCATTCTCCAGCTCTTCGG-3′ | ||||
| Nicotinamide adenine dinucleotide phosphate oxidase 1; (NOX1) | F: 5′-TCTTTCAAGCCTCGAGTCCC-3′ | 74 | 60 | XM_018044365.1 |
| R: 5′-AGGTCCATGAAGCTCAGTGATG-3′ | ||||
| Nicotinamide adenine dinucleotide phosphate oxidase 2 (NOX2) | F: 5′-ACGACCCAACTGGGATAACG-3′ | 127 | 60 | XM_005700924.3 |
| R: 5′-GGAGTTGGAGATGCACTGCT-3′ | ||||
| Cyclooxygenase-2 (COX2) | F: 5′-TCCCATCCATGCCAGAATCG-3′ | 77 | 60 | XM_018060731.1 |
| R: 5′-CCTGTTCGGGTACAGTCACA-3′ | ||||
| Prostaglandin E receptor 2 (PTGER2) | F: 5′-GGACACAAGCAGACCACGTA-3′ | 108 | 60 | NM_001314255.1 |
| R: 5′-CATGCGGATGAGGTTGACGA-3′ | ||||
| Arachidonate 12-Lipoxygenase (ALOX12) | F: 5′-AGGACTGCGCTCAAATCAGG-3′ | 83 | 60 | XM_018064507.1 |
| R: 5′-TCCTGGAGAGTGGGCTTCTC-3′ | ||||
| Arachidonate 5-Lipoxygenase Activating Protein (ALOX5AP) | F: 5′-ACTTTGTTGGCTACCTGGGG-3′ | 107 | 60 | XM_005687536.3 |
| R: 5′-GTTGAGTATCCCAGCGAGGG-3′ | ||||
| Leukotriene A4 Hydrolase (LTA4H) | F: 5′-TCCCTTTCTCTCGCGCTCAG-3′ | 78 | 61 | XM_005680471.3 |
| R:5′-GTGAGGAGTCCCGATGCAC-3′ | ||||
| Leukotriene C4 Synthase (LTC4S) | F: 5′-TGTCTAGGGCTGGAGGAAAG-3′ | 102 | 60 | XM_018051605.1 |
| R: 5′-CAGAAGTACCAGGGAGCAGATG-3′ | ||||
| Cytosolic phospholipase A2 (PLA2G4A) | F: 5′-TTGTGCTACAGAGAGGAGAGGA-3′ | 119 | 61 | XM_018060732.1 |
| R: 5′-GTGCCACGTAGCACCACTAC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriakaki, P.; Mavrommatis, A.; Tsiplakou, E. Schizochytrium spp. Dietary Supplementation Modulates Immune-Oxidative Transcriptional Signatures in Monocytes and Neutrophils of Dairy Goats. Antioxidants 2023, 12, 497. https://doi.org/10.3390/antiox12020497
Kyriakaki P, Mavrommatis A, Tsiplakou E. Schizochytrium spp. Dietary Supplementation Modulates Immune-Oxidative Transcriptional Signatures in Monocytes and Neutrophils of Dairy Goats. Antioxidants. 2023; 12(2):497. https://doi.org/10.3390/antiox12020497
Chicago/Turabian StyleKyriakaki, Panagiota, Alexandros Mavrommatis, and Eleni Tsiplakou. 2023. "Schizochytrium spp. Dietary Supplementation Modulates Immune-Oxidative Transcriptional Signatures in Monocytes and Neutrophils of Dairy Goats" Antioxidants 12, no. 2: 497. https://doi.org/10.3390/antiox12020497
APA StyleKyriakaki, P., Mavrommatis, A., & Tsiplakou, E. (2023). Schizochytrium spp. Dietary Supplementation Modulates Immune-Oxidative Transcriptional Signatures in Monocytes and Neutrophils of Dairy Goats. Antioxidants, 12(2), 497. https://doi.org/10.3390/antiox12020497