
In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.)

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Extract
2.2. Experimental
Compounds Isolation and Purification
2.3. In Vitro Model System of Protein Glycation
2.3.1. Determination of Fluorescent AGE Formation
2.3.2. Determination of the Formation of Markers Related to Amino Acid Oxidation
2.3.3. Quantification of Carbonylated Proteins
2.3.4. Modification of BSA via Crosslink Formation
2.3.5. Statistical Analysis
3. Results and Discussion
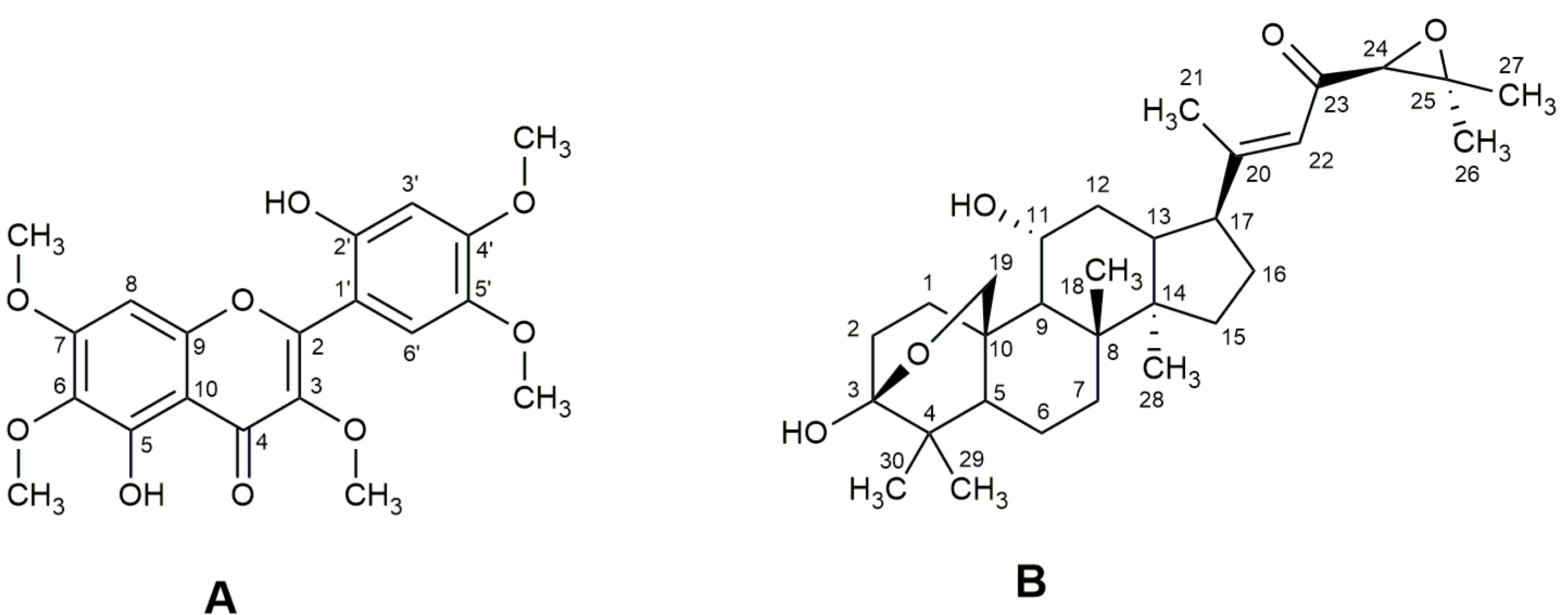
3.1. Identification of Isolated Compounds
3.2. Effects of EEVc, Brickellin and Cordialin A on Glycoxidation Changes in an In Vitro Model System of Protein Glycation
3.2.1. Formation of Fluorescent AGEs and Markers of Oxidation of Amino Acid Residues
3.2.2. Quantification of Carbonyl Groups in Proteins
3.2.3. Modification of BSA via Crosslinking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, H.; Matos, F.J.A. Plantas Medicinais no Brasil: Nativas e Exóticas; Instituto Plantarum: Nova Odessa, Brazil, 2002; Volume 1, p. 100. [Google Scholar]
- de Carvalho, P.M., Jr.; Rodrigues, R.F.O.; Sawaya, A.C.H.F.; Marques, M.O.M.; Shimizu, M.T. Chemical composition and antimicrobial activity of the essential oil of Cordia verbenacea D.C. J. Ethnopharmacol. 2004, 95, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Passos, G.F.; Fernandes, E.S.; da Cunha, F.M.; Ferreira, J.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Anti-inflammatory and anti-allergic properties of the essential oil and active compounds from Cordia verbenacea. J. Ethnopharmacol. 2006, 110, 323–333. [Google Scholar] [CrossRef] [PubMed]
- BRASIL Ministério da Saúde; Secretaria de Ciência, Tecnologia e Insumos Estratégicos, Departamento de Assistência Farmacêutica. Política Nacional de Plantas Medicinais e Fitoterápicos; Ministério da Saúde: Brasília, Brazil, 2006.
- BRASIL Ministério da Saúde; Agência Nacional de Vigilância Sanitária (ANVISA). Formulário de Fitoterápicos da Farmacopeia Brasileira, 2nd ed.; Ministério da Saúde: Brasília, Brazil, 2021; p. 57.
- Sertié, J.A.; Basile, A.C.; Panizza, S.; Matida, A.K.; Zelnik, R. Pharmacological assay of Cordia verbenacea. Part 1. Anti-inflammatory activity and toxicity of the crude extract of the leaves. Planta Med. 1988, 54, 7–10. [Google Scholar] [CrossRef]
- Sertié, J.A.A.; Basile, A.C.; Panizza, S.; Oshiro, T.T.; Azzolini, C.P.; Penna, S.C. Pharmacological assay of Cordia verbenacea III: Oral and topical antiinflammatory activity and gastrotoxicity of a crude leaf extract. J. Ethnopharmacol. 1991, 31, 239–247. [Google Scholar] [CrossRef] [PubMed]
- de Freitas Roldão, E.; Witaicenis, A.; Seito, L.N.; Hiruma-Lima, C.A.; Di Stasi, L.C. Evaluation of the antiulcerogenic and analgesic activities of Cordia verbenacea DC. (Boraginaceae). J. Ethnopharmacol. 2008, 119, 94–98. [Google Scholar] [CrossRef]
- Santi, M.M.; Sanches, F.S.; Silva, J.F.M.; Santos, P.M.L. Determinação do perfil fitoquímico de extrato com atividade antioxidante da espécie medicinal Cordia verbenacea DC. por HPLC-DAD. Rev. Bras. De Plantas Med. 2014, 16, 256–261. [Google Scholar] [CrossRef] [Green Version]
- Sertié, J.A.A.; Woisky, R.G.; Wiezel, G.; Rodrigues, M. Pharmacological assay of Cordia verbenacea V: Oral and topical anti-inflammatory activity, analgesic effect and fetus toxicity of a crude leaf extract. Phytomedicine 2005, 12, 338–344. [Google Scholar] [CrossRef]
- Martim, J.K.P.; Maranho, L.T.; Costa-Casagrande, T.A. Role of the chemical compounds present in the essential oil and in the extract of Cordia verbenacea DC as an anti-inflammatory, antimicrobial and healing product. J. Ethnopharmacol. 2021, 265, 113300. [Google Scholar] [CrossRef]
- Recio, M.C.; Andujar, I.; L Rios, J. Anti-inflammatory agents from plants: Progress and potential. Curr. Med. Chem. 2012, 19, 2088–2103. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bajwa, B.S.; Kuldeep, S.; Kalia, A.N. Anti-inflammatory activity of herbal plants: A review. Int. J. Adv. Pharm., Biol. Chem. 2013, 2, 272–281. [Google Scholar]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Liu, H.; Wang, J.; Sun, B. Inhibitory effect of phenolic compounds and plant extracts on the formation of advance glycation end products: A comprehensive review. Food Res. Int. 2020, 130, 108933. [Google Scholar] [CrossRef] [PubMed]
- Aajith, T.; Vinodkumar, P. Advanced glycation end products: Association with the pathogenesis of diseases and the current therapeutic advances. Curr. Clin. Pharmacol. 2016, 11, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Li, Y.; Ma, J.; Niu, L.; Tay, F.R. Clinical/translational aspects of advanced glycation end-products. Trends Endocrinol. Metab. 2019, 30, 959–973. [Google Scholar] [CrossRef] [PubMed]
- Jahan, H.; Choudhary, M.I. Glycation, carbonyl stress and AGEs inhibitors: A patent review. Expert Opin. Ther. Pat. 2015, 25, 1267–1284. [Google Scholar] [PubMed]
- Van Putte, L.; De Schrijver, S.; Moortgat, P. The effects of advanced glycation end products (AGEs) on dermal wound healing and scar formation: A systematic review. Scars Burn. Heal. 2016, 2, 2059. [Google Scholar] [CrossRef] [PubMed]
- Zieman, S.J.; Kass, D.A. Advanced glycation end products cross-linking: Pathophysiologic role and therapeutic target in cardiovascular disease. Congest. Heart Fail. 2004, 10, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Nemet, I.; Strauch, C.M.; Monnier, V.M. Favored and disfavored pathways of protein crosslinking by glucose: Glucose lysine dimer (GLUCOLD) and crossline versus glucosepane. Amino Acids 2011, 40, 167–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhuri, J.; Bains, Y.; Guha, S.; Kahn, A.; Hall, D.; Bose, N.; Gugliucci, A.; Kapahi, P. The role of advanced glycation end products in aging and metabolic diseases: Bridging association and causality. Cell Metab. 2018, 28, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef]
- Yeh, W.J.; Hsia, S.M.; Lee, W.H.; Wu, C.H. Polyphenols with antiglycation activity and mechanisms of action: A review of recent findings. J. Food Drug Anal. 2017, 25, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.H.C.; Talpo, T.C.; Motta, B.P.; Kaga, A.K.; Baviera, A.M.; Castro, R.N.; da Silva, V.C.; de Sousa-Junior, P.T.; Wessjohann, L.; de Carvalho, M.G. New compounds of Siolmatra brasiliensis and inhibition of in vitro protein glycation damage. Fitoterapia 2019, 133, 109–119. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G. Kinetics of glycoxidation of bovine serum albumin by methylglyoxal and glyoxal and its prevention by various compounds. Molecules 2014, 19, 4880–4896. [Google Scholar] [CrossRef] [Green Version]
- Séro, L.; Sanguinet, L.; Blanchard, P.; Dang, B.T.; Morel, S.; Richomme, P.; Séraphin, D.; Derbré, S. Tuning a 96-well microtiter plate fluorescence-based assay to identify age inhibitors in crude plant extracts. Molecules 2013, 18, 14320–14339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol 1994, 186, 346–357. [Google Scholar]
- Meeprom, A.; Sompong, W.; Chan, C.B.; Adisakwattana, S. Isoferulic acid, a new anti-glycation agent, inhibits fructose-and glucose-mediated protein glycation in vitro. Molecules 2013, 18, 6439–6454. [Google Scholar] [CrossRef] [Green Version]
- Motta, B.P.; Kaga, A.K.; Oliveira, J.O.; Inacio, M.D.; da Silva, C.F.; de Sousa Junior, P.T.; Brunetti, I.L.; Baviera, A.M. In vitro inhibition of protein glycation and advanced glycation end products formation by hydroethanolic extract and two fractions of Simaba trichilioides roots. Nat. Prod. Res 2020, 34, 2389–2393. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.A.S. Avaliação da Variabilidade do Metabolismo Secundário de Cordia verbenacea DC. Ph.D. Thesis, Faculdade de Ciências Farmacêuticas, UNESP, Araraquara, Brazil, 2017. [Google Scholar]
- Roberts, M.F.; Timmermann, B.N.; Mabry, T.J.; Brown, R.; Matlin, S.A. Brickellin, a novel flavone from Brickellia veronicaefolia and B. chlorolepis. Phytochemistry 1984, 23, 163–165. [Google Scholar] [CrossRef]
- IINUMA, M.; MATOBA, Y.; TANAKA, T.; MIZUNO, M. Flavonoids synthesis. I.: Synthesis and spectroscopic properties of flavones with two hydroxy and five methoxy groups at C-2’, 3’, 4’, 5, 6, 6’, 7 and C-2’, 3, 4’, 5, 5’, 6, 7. Chem. Pharm. Bull. 1986, 34, 1656–1662. [Google Scholar] [CrossRef] [Green Version]
- Muschietti, L.; Martino, V.; Ferraro, G.; Coussio, J. 5, 7, 5′-trihydroxy-3, 6, 2′, 4′-tetramethoxyflavone from Eupatorium buniifolium. Phytochemistry 1994, 36, 1085–1086. [Google Scholar] [CrossRef]
- Velde, V.V.; Lavie, D.; Zelnik, R.; Matida, A.K.; Panizza, S. Cordialin A and B, two new triterpenes from Cordia verbenacea DC. J. Chem. Soc., Perkin Trans. 1982, 1, 2697–2700. [Google Scholar] [CrossRef]
- Silverstein, R.M.; Bassler, G.C. Spectrometric Identification of Organic Compounds, 7rd ed; John Wiley & Sons, Inc.: New York, NY, USA, 2005; pp. 82–108. [Google Scholar]
- Chinchansure, A.A.; Korwar, A.M.; Kulkarni, M.J.; Joshi, S.P. Recent development of plant products with anti-glycation activity: A review. RSC Adv. 2015, 5, 31113–31138. [Google Scholar] [CrossRef]
- Arasteh, A.; Farahi, S.; Habibi-Rezaei, M.; Moosavi-Movahedi, A.A. Glycated albumin: An overview of the in vitro models of an in vivo potential disease marker. J. Diabetes Metab. Disord. 2014, 13, 49. [Google Scholar] [CrossRef] [Green Version]
- Atchade, T.; Dabole, B.; Adhikari, A.; Khan, A.; Mbafor, J.; Choudhary, M. Chemical constituents of Cordia platythyrsa and evaluation of their glycation and urease inhibition activities. Nat. Prod. 2012, 8, 346–351. [Google Scholar]
- Al-Musayeib, N.; Perveen, S.; Fatima, I.; Nasir, M.; Hussain, A. Antioxidant, anti-glycation and anti-inflammatory activities of phenolic constituents from Cordia sinensis. Molecules 2011, 16, 10214–10226. [Google Scholar] [CrossRef] [Green Version]
- Lins, A.P.; Alvarenga, M.A.; Gottlieb, O.R.; Oliveira, F. Two flavonois from Cordia verbenacea. Rev. Lat. Am. Química 1990, 21, 82. [Google Scholar]
- Bayeux, M.C.; Fernandes, A.T.; Foglio, M.A.; Carvalho, J.E. Evaluation of the antiedematogenic activity of artemetin isolated from Cordia curassavica DC. Braz. J. Med. Biol. Res. 2002, 35, 1229–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ticli, F.K.; Hage, L.I.; Cambraia, R.S.; Pereira, P.S.; Magro, Â.J.; Fontes, M.R.; Stábeli, R.G.; Giglio, J.R.; França, S.C.; Soares, A.M.; et al. Rosmarinic acid, a new snake venom phospholipase A2 inhibitor from Cordia verbenacea (Boraginaceae): Antiserum action potentiation and molecular interaction. Toxicon 2005, 46, 318–327. [Google Scholar] [CrossRef]
- Lameira, O.A.; Pinto, J.E.B.P.; Cardoso, M.G.; Arrigoni-Blank, M.F. Estabelecimento de cultura de células em suspensão e identificação de flavonóides em Cordia verbenacea DC. Rev. Bras. Plantas Med. 2009, 11, 7–11. [Google Scholar] [CrossRef]
- Matias, E.F.F.; Alves, E.F.; Santos, B.S.; Sobral de Souza, C.E.; Alencar Ferreira, J.V.D.; Santos de Lavor, A.K.L.; Figueredo, F.G.; Ferreira de Lima, L.; Vieira dos Santos, F.A.; Neves Peixoto, F.S.; et al. Biological activities and chemical characterization of Cordia verbenacea DC. as tools to validate the ethnobiological usage. Evid. Based Complement. Altern. Med. 2013, 2013, 164215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, E.F.; Santos, K.K.; Falcão-Silva, V.S.; Siqueira-Júnior, J.P.; Costa, J.G.; Coutinho, H.D. Modulation of the norfloxacin resistance in Staphylococcus aureus by Cordia verbenaceae DC. Indian J. Med. Res. 2013, 137, 178. [Google Scholar] [PubMed]
- Oza, M.J.; Kulkarni, Y.A. Traditional uses, phytochemistry and pharmacology of the medicinal species of the genus Cordia (Boraginaceae). J. Pharm. Pharmacol. 2017, 69, 755–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohn, S. Possibilities and limitations in the analysis of covalent interactions between phenolic compounds and proteins. Food Res. Int. 2014, 65, 13–19. [Google Scholar] [CrossRef]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bügel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef]
- Matsuda, H.; Wang, T.; Managi, H.; Yoshikawa, M. Structural requirements of flavonoids for inhibition of protein glycation and radical scavenging activities. Bioorg. Med. Chem. 2003, 11, 5317–5323. [Google Scholar] [CrossRef]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Res. Int. 2020, 129, 108843. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, X. Structures required of polyphenols for inhibiting advanced glycation end products formation. Curr. Drug Metab. 2013, 14, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Yin, M. Anti-glycative potential of triterpenes: A mini-review. BioMedicine 2012, 2, 2–9. [Google Scholar] [CrossRef]
- Motomura, K.; Fujiwara, Y.; Kiyota, N.; Tsurushima, K.; Takeya, M.; Nohara, T.; Nagai, R.; Ikeda, T. Astragalosides isolated from the root of astragalus radix inhibit the formation of advanced glycation end products. J. Agric. Food Chem. 2009, 57, 7666–7672. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hartley, D.P.; Liu, J. Protection against carbon tetrachloride hepatotoxicity by oleanolic acid is not mediated through metallothionein. Toxicol. Lett. 1998, 95, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Yamada, K.; Yoshikawa, N.; Nakamura, K.; Haginaka, J.; Kunitomo, M. Corosolic acid prevents oxidative stress, inflammation and hypertension in SHR/NDmcr-cp rats, a model of metabolic syndrome. Life Sci. 2006, 79, 2474–2479. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Narayanasamy, P. Flavonoid enhances the glyoxalase pathway in cerebellar neurons to retain cellular functions. Sci. Rep. 2017, 7, 5126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Hsu, C.C.; Huang, C.N.; Yin, M.C. Anti-glycative effects of oleanolic acid and ursolic acid in kidney of diabetic mice. Eur. J. Pharmacol. 2010, 628, 255–260. [Google Scholar] [CrossRef]
- Jang, S.M.; Kim, M.J.; Choi, M.S.; Kwon, E.Y.; Lee, M.K. Inhibitory effects of ursolic acid on hepatic polyol pathway and glucose production in streptozotocin-induced diabetic mice. Metabolism 2010, 59, 512–519. [Google Scholar] [CrossRef]
- Sudhahar, V.; Kumar, S.A.; Varalakshmi, P. Role of lupeol and lupeol linoleate on lipemic–oxidative stress in experimental hypercholesterolemia. Life Sci. 2006, 78, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Traaseth, N.J.; Davies, K.J.A. Tyrosine oxidation products: Analysis and biological relevance. Amino Acids 2003, 25, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Basran, J.; Efimov, I.; Chauhan, N.; Thackray, S.J.; Krupa, J.L.; Eaton, G.; Griffith, G.A.; Mowat, C.G.; Handa, S.; Raven, E.L. The mechanism of formation of N-formylkynurenine by heme dioxygenases. J. Am. Chem. Soc. 2011, 133, 16251–16257. [Google Scholar] [CrossRef]
- Manzanares, D.; Rodriguez-Capote, K.; Liu, S.; Haines, T.; Ramos, Y.; Zhao, L.; Doherty-Kirby, A.; Lajoie, G.; Possmayer, F. Modification of tryptophan and methionine residues is implicated in the oxidative inactivation of surfactant protein B. Biochemistry 2007, 46, 5604–5615. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.M.; Gibbs, B.F.; Kabashi, E.; Minotti, S.; Durham, H.D.; Agar, J.N. Tryptophan 32 potentiates aggregation and cytotoxicity of a copper/zinc superoxide dismutase mutant associated with familial amyotrophic lateral sclerosis. J. Biol. Chem. 2007, 282, 16329–16335. [Google Scholar] [CrossRef] [Green Version]
- Irvine, G.B.; El-Agnaf, O.M.; Shankar, G.M.; Walsh, D.M. Protein aggregation in the brain: The molecular basis for Alzheimer’s and Parkinson’s diseases. Mol. Med. 2008, 14, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Gieβauf, A.; van Wickern, B.; Simat, T.; Steinhart, H.; Esterbauer, H. Formation of N-formylkynurenine suggests the involvement of apolipoprotein B-100 centered tryptophan radicals in the initiation of LDL lipid peroxidation. FEBS Lett. 1996, 389, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Röhrig, U.F.; Awad, L.; Grosdidier, A.; Larrieu, P.; Stroobant, V.; Colau, D.; Cerundolo, V.; Simpson, A.J.; Vogel, P.; Van den Eynde, B.J.; et al. Rational design of indoleamine 2, 3-dioxygenase inhibitors. J. Med. Chem. 2010, 53, 1172–1189. [Google Scholar]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Porozov, Y.B.; Terentiev, A.A. Oxidative stress and advanced lipoxidation and glycation end products (ALEs and AGEs) in aging and age-related diseases. Oxidative Med. Cell. Longev. 2019, 2019, 3085756. [Google Scholar] [CrossRef] [Green Version]
- Schalkwijk, C.G.; Stehouwer, C.D.A. Methylglyoxal, a highly reactive dicarbonyl compound, in diabetes, its vascular complications, and other AGE-related diseases. Physiol. Rev. 2019, 100, 407–461. [Google Scholar] [CrossRef]
- Colzani, M.; De Maddis, D.; Casali, G.; Carini, M.; Vistoli, G.; Aldini, G. Reactivity, selectivity, and reaction mechanisms of aminoguanidine, hydralazine, pyridoxamine, and carnosine as sequestering agents of reactive carbonyl species: A comparative study. ChemMedChem 2016, 11, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- König, J.; Jung, T.; Grune, T. Protein carbonylation in aging and senescence. In Protein Carbonylation: Principles, Analysis, and Biological Implications; Ros, J., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2017; Volume 1, pp. 272–290. [Google Scholar]
- Dalle-Donne, I.; Aldini, G.; Carini, M.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation, cellular dysfunction, and disease progression. J Cell. Mol. Med. 2006, 10, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Vistoli, G.; De Maddis, D.; Cipak, A.; Zarkovic, N.; Carini, M.; Aldini, G. Advanced glycoxidation and lipoxidation end products (AGEs and ALEs): An overview of their mechanisms of formation. Free Radic. Res. 2013, 47 (Suppl. 1), 3–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emel’yanov, V.V. Glycation, antiglycation, and deglycation: Their role in aging mechanisms and geroprotective effects (literature review). Adv. Gerontol. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaklai, N.; Garlick, R.L.; Bunn, H.F. Nonenzymatic glycosylation of human serum albumin alters its conformation and function. J. Biol. Chem. 1984, 259, 3812–3817. [Google Scholar] [CrossRef]
- Szkudlarek, A.; Maciążek-Jurczyk, M.; Chudzik, M.; Równicka-Zubik, J.; Sułkowska, A. Alteration of human serum albumin tertiary structure induced by glycation: Spectroscopic study. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 153, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Okabe, N.; Hashizume, N. Drug binding properties of glycosylated human serum albumin as measured by fluorescence and circular dichroism. Biol. Pharm. Bull. 1994, 17, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Ghuman, J.; Zunszain, P.A.; Petitpas, I.; Bhattacharya, A.A.; Otagiri, M.; Curry, S. Structural basis of the drug-binding specificity of human serum albumin. J. Mol. Biol. 2005, 353, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.A.; Pea, F.; Lipman, J. The clinical relevance of plasma protein binding changes. Clin. Pharmacokinet. 2013, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Trynda-Lemiesz, L.; Wiglusz, K. Effects of glycation on meloxicam binding to human serum albumin. J. Mol. Struct. 2011, 995, 35–40. [Google Scholar] [CrossRef]
- Morais, M.P.P.; Mackay, J.D.; Bhamra, S.K.; Buchanan, J.G.; James, T.D.; Fossey, J.S.; van den Elsen, J.M.H. Analysis of protein glycation using phenylboronate acrylamide gel electrophoresis. In Protein Electrophoresis; Humana Press: Totowa, NJ, USA, 2010; pp. 93–109. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | δ ¹³C | δ 1H J | δ ¹³C a | δ 1H J a | δ 1H J b | δ ¹³C c | δ 1H J c |
|---|---|---|---|---|---|---|---|
| 2 | 155.9 s | ---- | 155.5 s | ---- | ---- | 155.4 s | ---- |
| 3 | 136.6 s | ---- | 136.3 s | ---- | ---- | 140.4 s | ---- |
| 4 | 177.4 s | ---- | 176.9 s | ---- | ---- | 179.0 s | ---- |
| 5 | 152.8 s | ---- | 152.3 s | ---- | ---- | 153.0 s | ---- |
| 6 | 132.6 s | ---- | 132.0 s | ---- | ---- | 132.6 s | ---- |
| 7 | 159.1 s | ---- | 158.5 s | ---- | ---- | 158.7 s | ---- |
| 8 | 90.6 d | 6.52 s | 90.4 d | 6.35 s | 6.62 s | 90.6 d | 6.67 s |
| 9 | 152.8 s | ---- | 153.4 s | ---- | ---- | 155.4 s | ---- |
| 10 | 106.4 s | ---- | 105.9 s | ---- | ---- | 113.1 s | ---- |
| 1′ | 108.7 s | ---- | 108.2 s | ---- | ---- | 107.1 s | ---- |
| 2′ | 154.0 s | ---- | 152.3 s | ---- | ---- | 152.9 s | ---- |
| 3′ | 103.0 d | 6.64 s | 102.6 d | 6.45 s | 6.59 s | 99.9 d | 6.43 s |
| 4′ | 151.3 s | ---- | 150.8 s | ---- | ---- | 152.9 s | ---- |
| 5′ | 143.6 s | ---- | 143.1 s | ---- | ---- | 140.4 s | ---- |
| 6′ | 111.2 d | 7.11 s | 110.9 d | 6.88 s | 7.10 s | 110.2 d | 6.94 s |
| OMe | 62.1 q | 3.89 s | 61.7 q | 3.70–4.00 s | 3.80 s | 60.9 q | 3.79 s |
| OMe | 60.8 q | 3.93 s | 60.5 q | 3.70–4.00 s | 3.90 s | 60.6 q | 3.80 s |
| OMe | 56.0 q | 3.97 s | 56.5 q | 3.70–4.00 s | 3.93 s | 56.3 q | 3.91 s |
| OMe | 56.4 q | 3.94 s | 56.1 q | 3.70–4.00 s | 3.93 s | 56.4 q | 3.91 s |
| OMe | 56.8 q | 3.93 s | 55.7 q | 3.70–4.00 s | 3.96 s | 56.8 q | 3.91 s |
| 2′-OH | ---- | 7.90 s | ---- | ---- | 7.87 s | ---- | ---- |
| 5-OH | ---- | 12.38 s | ---- | ---- | 12.36 s | ---- | ---- |
| C | δ ¹³C | δ 1H J (Hz) | δ ¹³C a | δ 1H J (Hz) a | δ ¹³C b | δ 1H J (Hz) b |
|---|---|---|---|---|---|---|
| 1 | 30.0 t | 3.12 td (12.4; 5.9) | 29.8 t | 3.12 td (12.5; 6.0) | 30.0 t | 3.11 td (12.7; 5.9) |
| 2 | 37.8 t | nd * | 37.8 t | nd | 37.9 t | nd |
| 3 | 98.4 s | ----- | 98.7 s | ---- | 98.6 s | ---- |
| 4 | 36.1 s | ----- | 36.1 s | ---- | 36.1 s | ---- |
| 5 | 51.0 d | 1.31 dl (2.8) | 50.9 d | nd | 50.9 d | 1.31 dl (3.3) |
| 6 | 19.4 t | 1.66 m 1.48 m | 19.4 t | nd | 19.4 t | 1.67 dd (12.3; 3.3) 1.48 m |
| 7 | 34.3 t | nd | 34.3 t | nd | 34.4 t | nd |
| 8 | 39.9 s | ---- | 39.9 s | ---- | 39.9 s | ---- |
| 9 | 50.1 d | 1.57 d (10.8) | 50.0 d | nd | 50.1 d | 1.56 d (11.0) |
| 10 | 40.9 s | nd | 41.0 s | nd | 41.0 s | nd |
| 11 | 70.9 d | 3.67 (10.8; 4.0) | 70.5 d | 3.65 td (10.5; 4.2) | 70.8 d | 3.65 td (11.0; 3.9) |
| 12 | 37.2 t | 1.89 m | 36.9 t | nd | 37.2 t | 1.85 td (11.2; 3.9) |
| 13 | 44.6 d | nd | 44.8 d | nd | 44.7 d | nd |
| 14 | 49.0 s | nd | 49.0 s | ---- | 49.0 s | ---- |
| 15 | 31.6 t | ---- | 31.7 t | nd | 31.7 t | nd |
| 16 | 27.8 t | nd | 27.9 t | nd | 27.8 t | nd |
| 17 | 51.3 d | 2.38 td (10.4; 6.0) | 51.5 d | 2.40 td (11.0; 6.0) | 51.4 d | 2.38 td (10.5; 5.7) |
| 18 | 15.4 q | 0.95 s | 15.4 q | 0.95 s | 15.4 q | 0.95 s |
| 19 | 67.8 t | 4.16 dd (8.6; 1.4) 4.33 dd (8.6; 2.5) | 67.8 t | α-H 4.15 dd (8.5; 0.7) β-H 4.32 dd (8.5; 2.0) | 67.9 t | 4.15 dd (8.5;1.0) 4.31 dd (8.5; 2.5) |
| 20 | 164.3 s | ---- | 164.8 s | ----- | 164.6 s | ---- |
| 21 | 17.2 q | 2.12 d (0.8) | 17.3 q | 2.12 sl | 17.3 q | 2.11 sl |
| 22 | 120.3 d | 6.28 sl | 120.4 d | 6.28 sl | 120.5 d | 6.27 sl |
| 23 | 195.7 s | ---- | 195.7 s | ---- | 195.7 s | ---- |
| 24 | 66.3 d | 3.34 s | 66.4 d | 3.34 s | 66.4 d | 3.33 s |
| 25 | 61.0 s | ---- | 61.2 s | ---- | 61.2 d | ---- |
| 26 | 24.9 q | 1.43 s | 24.9 q | 1.42 s | 25.0 d | 1.42 s |
| 27 | 18.7 q | 1.27 s | 18.6 q | 1.27 s | 18.7 q | 1.26 s |
| 28 | 26.3 q | 1.04 s | 26.4 q | 1.03 s | 26.4 q | 1.03 s |
| 29 | 18.9 q | 1.01 s | 18.8 q | 1.01 s | 18.9 q | 1.00 s |
| 30 | 16.8 q | 0.91 s | 16.8 q | 0.90 s | 16.9 q | 0.90 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, W.D.; Cardoso, F.N.; Baviera, A.M.; dos Santos, A.G. In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.). Antioxidants 2023, 12, 522. https://doi.org/10.3390/antiox12020522
Rodrigues WD, Cardoso FN, Baviera AM, dos Santos AG. In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.). Antioxidants. 2023; 12(2):522. https://doi.org/10.3390/antiox12020522
Chicago/Turabian StyleRodrigues, Winner Duque, Felipe Nunes Cardoso, Amanda Martins Baviera, and André Gonzaga dos Santos. 2023. "In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.)" Antioxidants 12, no. 2: 522. https://doi.org/10.3390/antiox12020522
APA StyleRodrigues, W. D., Cardoso, F. N., Baviera, A. M., & dos Santos, A. G. (2023). In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.). Antioxidants, 12(2), 522. https://doi.org/10.3390/antiox12020522