

Ramifications of Heat Stress on Rabbit Production and Role of Nutraceuticals in Alleviating Its Negative Impacts: An Updated Review



Abstract
:1. Introduction
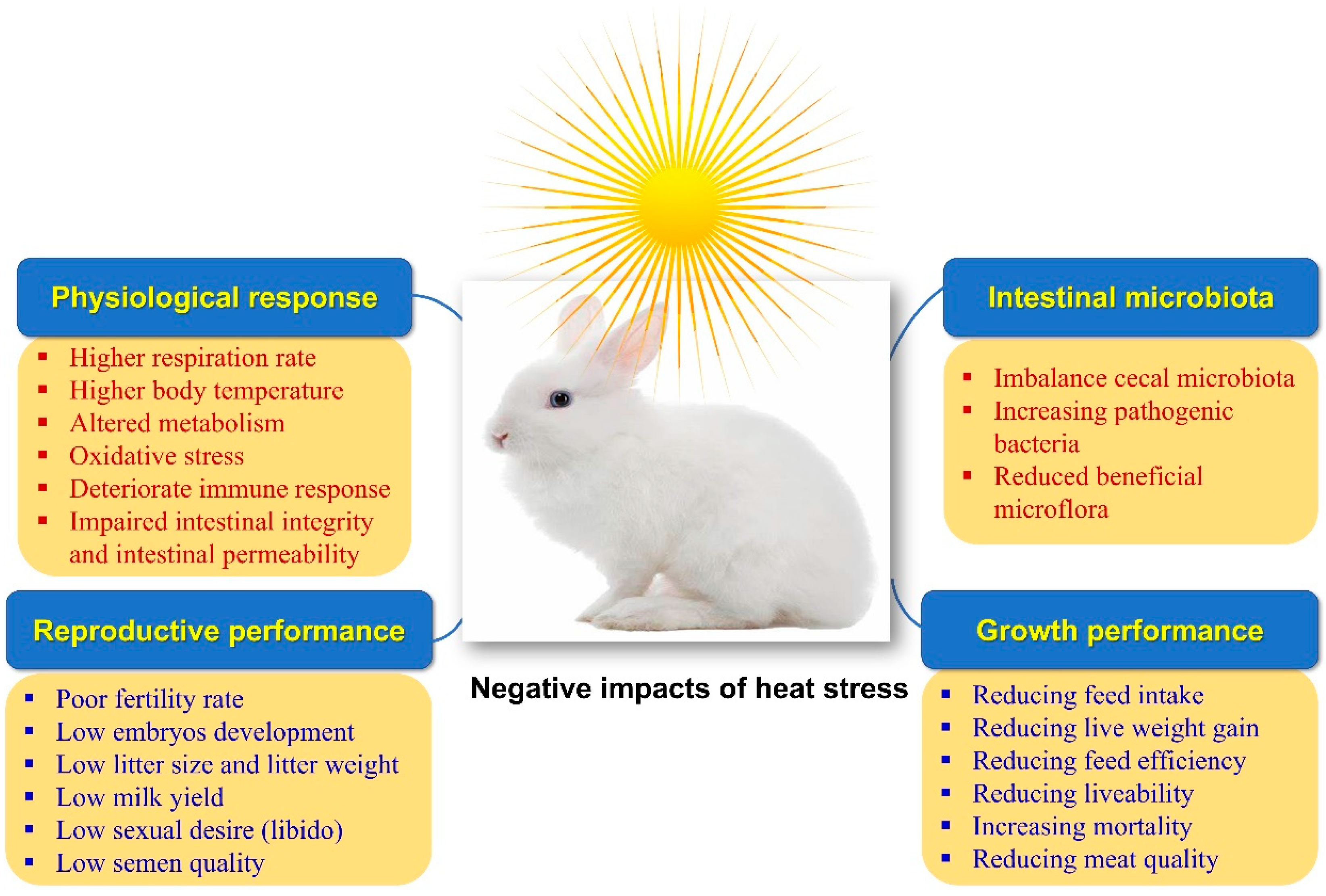
2. Effect of HS on Growth Performance
3. Effect of HS on Reproductive Performance

{kind=link}
{kind=link}
| Additives | Level | Heat Stress Conditions | Animal | Main Impacts | References |
|---|---|---|---|---|---|
| Vitamin C | 0.5 g/kg dietfrom 5 to 14 wk of age | 32.44 °C and 84.67% relative humidity | Giant Flander male growing rabbits |
| [38] |
| Vitamin C | 200 mg/kg diet from 6 to 12 wk of age | 28–39 °C and 60% relative humidity | New Zealand White growing rabbits |
| [39] |
| Vitamin E | 0.25 g/kg diet from 7 to 14 wk of age | 36.4 °C and 97% relative humidity | Californian unsexed growing rabbits |
| [40] |
| Vitamin E | 100 mg/kg diet for 3 months | 32.9 °C and 80.38% relative humidity | New Zealand White rabbits does |
| [41] |
| Vitamin A | 12,000 IU/kg diet | 30–34 °C | Rex rabbits |
| [42] |
| Selenium | 25 and 50 mg of nano-Se/kg diet from 7 to 13 wk of age | 33 °C and 90% relative humidity | Domestic rowing rabbits |
| [43] |
| Selenium | 0.3 mg organic Se/kg diet for 12 wk | 31 °C and 75% relative humidity | Adult V-line male rabbits |
| [44] |
| Zinc | 20, 40, 60, and 80 mg nano-Zn/kg dietfor 60 d | 38.20–40.10 °C and 45–50% relative humidity | New Zealand White male growing rabbits |
| [16] |
| Zinc | 75 mg ZnSO4/kg diet or 75 mg Zn picolinate/kg diet from 32 to 42 wk of age | 30.7–37.6 °C and 70–80% relative humidity | New Zealand White rabbit bucks |
| [45] |
| Copper | 200 mg Cu-methionine/kg diet or 200 mg copper-glycine/kg diet for 5 wk | 30.12 °C and 82.40% relative humidity | V line unsexed growing rabbits |
| [46] |
| Copper | 100 mg Cu-acetate/kg diet or 50 mg nano-Cu/kg diet from 5 to 14 wk of age | 17 and 22 °C | New Zealand White unsexed growing rabbits |
| [47] |
| Chromium | 0.4–1.6 mg organic Cr/kg diet | 30 °C and 80% relative humidity | Growing rabbits |
| [48] |
| Chromium | 2.5 mg Cr-yeast/kg diet for 198 d | 29.3 °C and 71% relative humidity | New Zealand White male rabbits |
| [49] |
| Probiotics | 5 × 106 CFU Clostridium butyricum, 2 × 108 CFU Enterococcus faecium, or 2.5 × 106 CFU C. butyricum + 1 × 108 CFU E. faecium/kg diet from 35 to 91 d of age | 31.78 °C and 60.19% relative humidity | New Zealand White male growing rabbits |
| [50] |
| Probiotics | 3 × 109 CFU Saccharomyces cerevisiae/kg or 3 × 109 CFU Lactobacillus acidophilus/kg from 5 to 13 wk of age | 33 °C and 81% relative humidity | New Zealand White growing rabbits |
| [51] |
| Prebiotics | 0.3% mannan-oligosaccharides or 0.05% isomalto-oligosaccharide from 6 to 16 wk of age | 19 °C | New Zealand White male growing rabbits |
| [19] |
| Prebiotics | 3 g Bio-Mos®/kg diet from 4 to 12 wk of age | 31.50 °C and 79.07% relative humidity | New Zealand White male growing rabbits |
| [7] |
| Moringa oleifera | 200 mg Moringa oleifera leaves powder/kg BW daily for 4 wk (from 32 to 32 wk of age) | 35 °C and 80% relative humidity | New Zealand White male rabbits |
| [52,53] |
| Moringa oleifera | 50 mg Moringa oleifera leaves ethanolic extract/kg BW for 12 consecutive weeks | 31.11 °C and 87% relative humidity | V-line rabbit bucks |
| [54] |
| Ginger | 7.5 g ginger powder/kg diet from 5 to 13 wk of age | 33 °C and 74.5% relative humidity | APRI growing rabbits |
| [55] |
| Ginger | 250 mg ginger/doe/d for 8 wk | 35 °C and 80% relative humidity | New Zealand White virgin female rabbits |
| [56] |
| Thyme | 16 g thyme/kg diet for 90 d | 39 °C and 30–35% relative humidity | New Zealand White male rabbits |
| [57] |
| Thyme | 100 thyme essential oil mg/kg diet from 6 to 9 months of age | 33 °C and 80% relative humidity | APRI rabbit does |
| [58] |
| Turmeric | 2.5 g turmeric nanoparticles/kg diet from 5 to 13 wk of age | 32.77 °C and 43.23% relative humidity | APRI growing rabbits |
| [59] |
| Maca (Lepidium meyenii) | 400 or 600 mg maca extract/head twice weekly | 33 °C and 80% relative humidity | V-line rabbit bucks |
| [24] |
| Pumpkin | 2 mL pumpkin seed essential oil/kg diet from 5 to 13 wk of age | 38 °C and 60% relative humidity | New Zealand White male growing rabbits |
| [60] |
4. Effect of HS on Carcass Traits and Meat Quality
5. Effect of HS on the Intestinal Microbiome
6. Effect of HS on Antioxidative Properties
7. Effect of HS on Immune Responsiveness
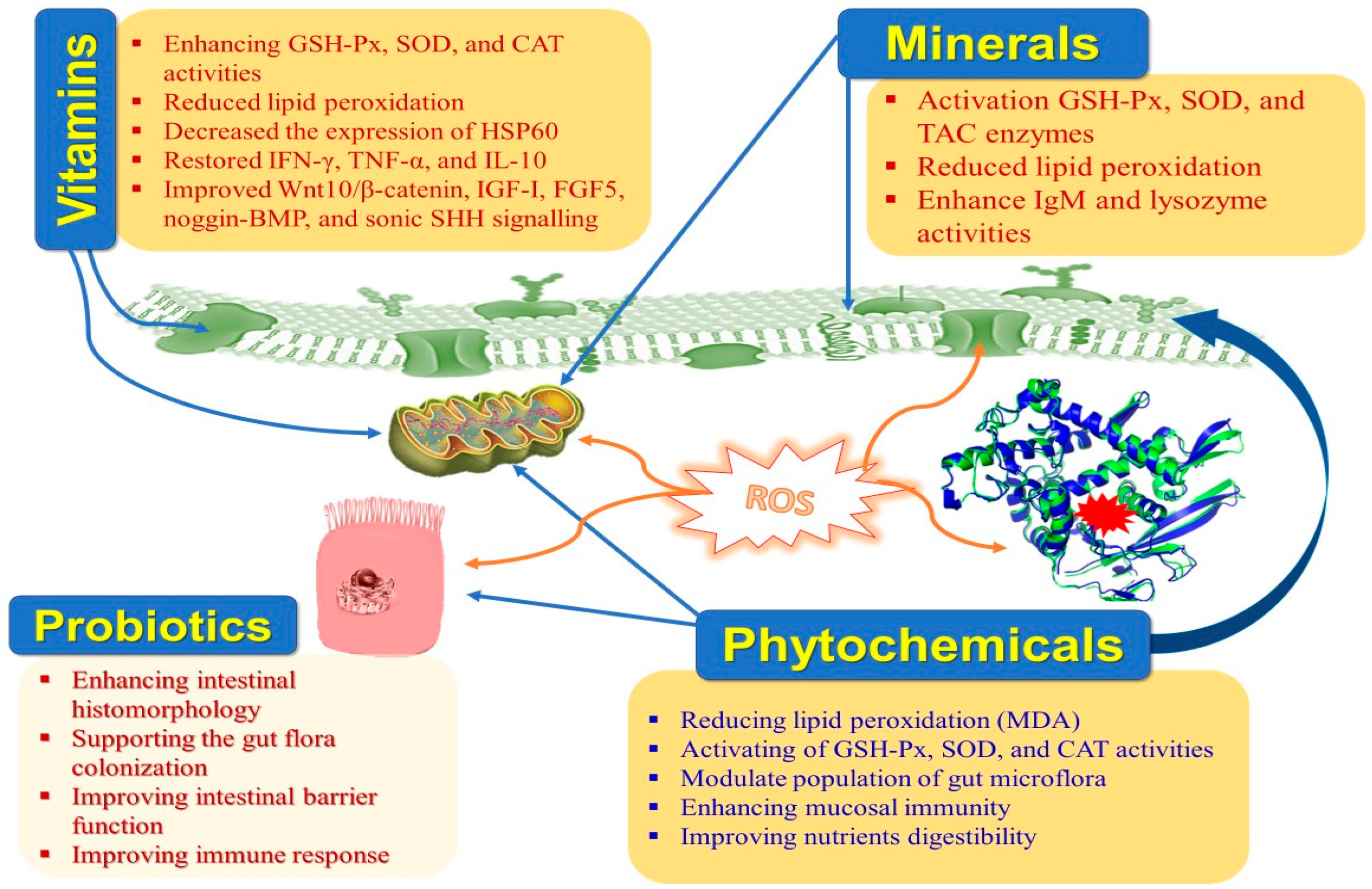
8. Dietary Manipulation to Mitigate Heat-Stressed Rabbits
8.1. Vitamins
8.1.1. Vitamin C
8.1.2. Vitamin E
8.1.3. Vitamin A
8.2. Minerals
8.2.1. Selenium
8.2.2. Zinc
8.2.3. Copper
8.2.4. Chromium
8.3. Electrolytes
8.4. Probiotics
8.5. Prebiotics
8.6. Phytobiotics
8.6.1. Moringa oleifera
8.6.2. Ginger
8.6.3. Thyme
8.6.4. Turmeric
8.6.5. Other Phytobiotics
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dalle Zotte, A.; Szendrő, Z. The Role of Rabbit Meat as Functional Food. Meat Sci. 2011, 88, 319–931. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.E.; Shehata, A.M.; Khidr, R.E.; Paswan, V.K.; Ibrahim, N.S.; El-Ghoul, A.A.; Aldhumri, S.A.; Gabr, S.A.; Mesalam, N.M.; Elbaz, A.M.; et al. Nutritional Manipulation to Combat Heat Stress in Poultry—A Comprehensive Review. J. Therm. Biol. 2021, 98, 102915. [Google Scholar] [CrossRef]
- Liang, Z.L.; Chen, F.; Park, S.; Balasubramanian, B.; Liu, W.C. Impacts of Heat Stress on Rabbit Immune Function, Endocrine, Blood Biochemical Changes, Antioxidant Capacity and Production Performance, and the Potential Mitigation Strategies of Nutritional Intervention. Front. Vet. Sci. 2022, 9, 906084. [Google Scholar] [CrossRef]
- Matics, Z.; Gerencsér, Z.; Kasza, R.; Terhes, K.; Nagy, I.; Radnai, I.; Dalle Zotte, A.; Cullere, M.; Szendrő, Z. Effect of Ambient Temperature on the Productive and Carcass Traits of Growing Rabbits Divergently Selected for Body Fat Content. Animal 2021, 15, 100096. [Google Scholar] [CrossRef]
- Oladimeji, A.M.; Johnson, T.G.; Metwally, K.; Farghly, M.; Mahrose, K.M. Environmental Heat Stress in Rabbits: Implications and Ameliorations. Int. J. Biometeorol. 2022, 66, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.; Haeeb, A.A.M.; Gad, A.E. Biological Functions in Young Pregnant Rabbit Does as Affected by Heat Stress and Lighting Regime under Subtropical Conditions of Egypt. Trop. Subtrop. Agroecosyst. 2007, 7, 165–176. [Google Scholar]
- Ayyat, M.S.; Al-Sagheer, A.A.; Abd El-Latif, K.M.; Khalil, B.A. Organic Selenium, Probiotics, and Prebiotics Effects on Growth, Blood Biochemistry, and Carcass Traits of Growing Rabbits During Summer and Winter Seasons. Biol. Trace. Elem. Res. 2018, 186, 162–173. [Google Scholar] [CrossRef]
- Farghly, M.F.A.; Mahrose, K.M.; Mahmoud, G.B.; Ali, R.M.; Daghash, W.; Metwally, K.A.; Abougaba, M.S. Lighting Programs as an Appliance to Improve Growing New Zealand White Rabbit’s Performance. Inter. J. Biometeorol. 2020, 64, 1295–1303. [Google Scholar] [CrossRef]
- El-Badawi, A.Y.; El-Wardany, I.; Abd El-Moez, S.I.; Helal, F.I.S.; Ali, N.G.M.; Shourrap, M.I.; Aboelazab, O.M. Impact of Dietary Moringa oleifera Leaves on Intestinal Pathogenic Load and Histological Structure of Growing Rabbits Raised under Heat-Stress Conditions. Anim. Prod. Sci. 2018, 58, 1901–1907. [Google Scholar] [CrossRef]
- Abdelsalam, M.; Al-Homidan, I.; Ebeid, T.; Abou-Emera, O.; Mostafa, M.; Abd El-Razik, M.; Shehab-El-Deen, M.; Abdel Ghani, S.; Fathi, M. Effect of Silver Nanoparticles Administration on Productive Performance, Blood Parameters, Antioxidative Status, and Silver Residues in Growing Rabbits under Hot Climate. Animals 2019, 9, 845. [Google Scholar] [CrossRef] [Green Version]
- Ebeid, T.A.; Ketta, M.; Al-Homidan, I.H.; Barakat, H.; Abdel-Moneim, E.A.M. In Ovo Feeding of Nutraceuticals and its Role in Adjusting the Gastrointestinal Tract, Antioxidative Properties, Immunological Response, and Performance in Poultry: An Updated Review. Czech J. Anim. Sci. 2023, 68, 1–16. [Google Scholar] [CrossRef]
- Tůmová, E.; Chodová, D.; Volek, Z.; Ebeid, T.A.; Ketta, M.; Skřivanová, V. A Comparative Study on the Effect of Quantitative Feed Restriction in Males and Females of Broiler Chickens, Rabbits and Nutrias. I. Performance and Carcass Composition. Czech J. Anim. Sci. 2022, 67, 47–54. [Google Scholar] [CrossRef]
- Tůmová, E.; Chodová, D.; Volek, Z.; Ebeid, T.A.; Ketta, M.; Skřivanová, V. A Comparative Study on the Effect of Quantitative Feed Restriction in Males and Females of Broiler Chickens, Rabbits and Nutrias. II. Meat Quality. Czech J. Anim. Sci. 2022, 67, 55–64. [Google Scholar] [CrossRef]
- Dalle Zotte, A.; Cullere, M.; Gleeson, E.Y.; Cossu, M.E. Animal Fat and Vitamin E in Rabbit Diets: Total Tract Apparent Digestibility, Growth Performance, Carcass and Meat Quality Traits. Czech J. Anim. Sci. 2020, 65, 380–388. [Google Scholar] [CrossRef]
- Al-Homidan, I.; Fathi, M.; Abdelsalam, M.; Ebied, T.; Abou-Emera, O.; Mostafa, M.; Abd El-Razik, M.; Shehab-El-Deen, M. Effect of Propolis Supplementation and Breed on Growth Performance, Immunity, Blood Parameters and Cecal Microbiota in Growing Rabbits. Anim. Biosci. 2022, 35, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wareth, A.A.A.; Amer, S.A.; Mobashar, M.; El-Sayed, H.G.M. Use of Zinc Oxide Nanoparticles in the Growing Rabbit Diets to Mitigate Hot Environmental Conditions for Sustainable Production and Improved Meat Quality. BMC Vet. Res. 2022, 18, 354. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wareth, A.A.A.; Mohamed, E.M.H.; Hassan, H.A.; Eldeek, A.A.; Lohakare, J. Effect of Substituting Hydroponic Barley Forage with or without Enzymes on Performance of Growing Rabbits. Sci. Rep. 2023, 13, 943. [Google Scholar] [CrossRef] [PubMed]
- El-Deep, M.H.; Dawood, M.A.O.; Assar, M.H.; Ahamad Paray, B. Aspergillus awamori Positively Impacts the Growth Performance, Nutrient Digestibility, Antioxidative Activity and Immune Responses of Growing Rabbits. Vet. Med. Sci. 2021, 7, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, A.H.; El-Kasrawy, N.I.; Abd El-Hack, M.E.; Kamel, S.Z.; Mahrous, U.E.; El-Deeb, E.M.; Atta, M.S.; Amer, M.S.; Naiel, M.A.E.; Khafaga, A.F.; et al. Growth, Immunity, Relative Gene Expression, Carcass Traits and Economic Efficiency of Two Rabbit Breeds Fed Prebiotic Supplemented Diets. Anim. Biotechnol. 2022, 33, 417–428. [Google Scholar] [CrossRef]
- Abd El-Aziz, A.H.; Abo Ghanima, M.M.; Alsanie, W.F.; Gaber, A.; Alsenosy, A.E.; Easa, A.A.; Moawed, S.A.; Raza, S.H.A.; Elfadadny, A.; Yossef, H.A.; et al. Fructooligosaccharide Supplementation Boosts Growth Performance, Antioxidant Status, and Cecal Microbiota Differently in Two Rabbit Breeds. Animals 2022, 12, 1528. [Google Scholar] [CrossRef]
- Nwachukwu, C.U.; Aliyu, K.I.; Ewuola, E.O. Growth Indices, Intestinal Histomorphology, and Blood Profile of Rabbits Fed Probiotics- and Prebiotics-Supplemented Diets. Transl. Anim. Sci. 2021, 5, txab096. [Google Scholar] [CrossRef]
- Fathi, M.; Abdelsalam, M.; Al-Homidan, I.; Ebeid, T.; Shehab-El-Deen, M.; Abd El-Razik, M.; Abou-Emera, O.; Mostafa, M. Supplemental Effects of Eucalyptus (Eucalyptus camaldulensis) Leaves on Growth Performance, Carcass Characteristics, Blood Biochemistry and Immune Response of Growing Rabbits. Ann. Anim. Sci. 2019, 19, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Mutwedu, V.B.; Nyongesa, A.W.; Kitaa, J.M.; Ayagirwe, R.B.B.; Baharanyi, C.; Mbaria, J.M. Effects of Moringa oleifera Aqueous Seed Extracts on Reproductive Traits of Heat-Stressed New Zealand White Female Rabbits. Front. Vet. Sci. 2022, 9, 883976. [Google Scholar] [CrossRef]
- Ragab, M.A.; Hassan, M.A.E.; Shazly, S.A.; El-Kholany, M.E.; Ahmed, M.E.; El-Raghi, A.A. The Benefits of Maca (Lepidium meyenii) Extract Administration for Male Rabbits Affected by Environmental Heat Stress. J. Anim. Physiol. Anim. Nutr. 2023, 107, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Szendrő, Z.; Papp, Z.; Kustos, K. Effect of Ambient Temperature and Restricted Feeding on the Production of Rabbit Does and Their Kits. Acta Agraria. Kaposváriensis 2018, 22, 1–17. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Parkanyi, V.; Pivko, J. High Temperature Impairs Rabbit Viability, Feed Consumption, Growth and Fecundity: Examination of Endocrine Mechanisms. Domest. Anim. Endocrinol. 2021, 74, 106478. [Google Scholar] [CrossRef]
- Dahmani, Y.; Benali, N.; Saidj, D.; Chirane, M.; Ainbaziz, H.; Temim, S. Effects of Heat Stress on Growth Performance, Carcass Traits, Physiological Components, and Biochemical Parameters in Local Algerian Growing Rabbits. World Vet. J. 2022, 12, 405–417. [Google Scholar] [CrossRef]
- Ragab, M.A.; Shazly, S.A.; Ibrahem, M.A.; El-Kholany, M.E.; Khalil, W.A. Black Maca (Lepidium meyenii Walp.) Hydroalcoholic Extract as an Ameliorating Agent Against Heat Stress Conditions of V-Line Rabbit Does. Sustainability 2022, 14, 15154. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, B.; Li, F.; Liu, L.; Yang, T.; Zhang, H.; Li, F. Effects of Heat Stress on Growth Performance, Carcass Traits, Serum Metabolism, and Intestinal Microflora of Meat Rabbits. Front. Microbiol. 2022, 13, 998095. [Google Scholar] [CrossRef]
- Slimen, B.I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Bakr, M.H.; Tusell, L.; Rafel, O.; Terré, M.; Sánchez, J.P.; Piles, M. Lactating Performance, Water and Feed Consumption of Rabbit Does Reared under a Mediterranean Summer Circadian Cycle of Temperature v. Comfort Temperature Conditions. Animal 2015, 9, 1203–1209. [Google Scholar] [CrossRef] [Green Version]
- Al-Sagheer, A.A.; Daader, A.H.; Gabr, H.A.; Abd El-Moniem, E.A. Palliative Effects of Extra Virgin Olive Oil, Gallic Acid, and Lemongrass Oil Dietary Supplementation on Growth Performance, Digestibility, Carcass Traits, and Antioxidant Status of Heat-Stressed Growing New Zealand White Rabbits. Environ. Sci. Pollut. Res. 2017, 24, 6807–6818. [Google Scholar] [CrossRef]
- Ayyat, M.S.; El-Latif, A.; Khaled, M.; Helal, A.A.; Al-Sagheer, A.A. Interaction of Supplementary L-Carnitine and Dietary Energy Levels on Feed Utilization and Blood Constituents in New Zealand White Rabbits Reared under Summer Conditions. Trop. Anim. Health. Prod. 2021, 53, 279. [Google Scholar] [CrossRef]
- Saghir, S.A.M.; Hroob, A.M.; Majrashi, K.A.; Jaber, F.A.; Abduh, M.S.; Al-Gabri, N.; Albaqami, N.M.; Abdelnour, S.A.; Alqhtani, A.H.; Abd El-Hack, M.E.; et al. Effects of Alginates on the Growth, Haematological, Immunity, Antioxidant and Pro-inflammatory Responses of Rabbits under High Temperature. Res. Vet. Sci. 2023, 155, 36–43. [Google Scholar] [CrossRef]
- Hassan, R.A.; Ebeid, T.A.; Abd El-Lateif, A.I.; Ismail, N.B. Effect of Dietary Betaine Supplementation on Growth, Carcass and Immunity of New Zealand White Rabbits under High Ambient Temperature. Livest. Sci. 2011, 135, 103–109. [Google Scholar] [CrossRef]
- Marai, I.F.M.; Habee, A.A.M.; Gad, A.E. Rabbits’ Productive, Reproductive and Physiological Performance Traits as Affected by Heat Stress: A Review. Livest. Prod. Sci. 2002, 78, 71–90. [Google Scholar] [CrossRef]
- Marco-Jiménez, F.; García-Diego, F.J.; Vicente, J.S. Effect of Gestational and Lactational Exposure to Heat Stress on PERFORMANCE in Rabbits. World Rabbit Sci. 2017, 25, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Hassan, F.A.; Shalaby, A.G.; Elkassas, N.E.M.; El-Medany, S.A.; Hamdi Rabie, A.; Mahrose, K.; Abd El-Aziz, A.; Bassiony, S. Efficacy of Ascorbic Acid and Different Sources of Orange Peel on Growth Performance, Gene Expression, Anti-Oxidant Status and Microbial Activity of Growing Rabbits under Hot Conditions. Anim. Biotechnol. 2022, 23, 1–12. [Google Scholar]
- Abdel-Latif, M.; Sakran, T.; Badawi, Y.K.; Abdel-Hady, D.S. Influence of Moringa oleifera Extract, Vitamin C, and Sodium Bicarbonate on Heat Stress-Induced Hsp70 Expression and Cellular Immune Response in Rabbits. Cell Stress Chaperone. 2018, 23, 975–984. [Google Scholar] [CrossRef]
- Sherif, S.K. Response of Growing Rabbits to Stoking Density and Dietary Supplementation with Ascorbic Acid and Vitamin E under Summer Conditions. Egypt. Poult. Sci. J. 2018, 38, 831–846. [Google Scholar] [CrossRef] [Green Version]
- El-Ratel, I.; Gabr, A.A. Effect of Spirulina and Vitamin E on Reproduction and in vitro Embryo Production in Heat-Stressed Rabbits. Pak. J. Biol. Sci. 2019, 22, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Z.; Li, C.; Liu, Y.; Liu, M.; Zhao, M.; Li, F.; Liu, L. Vitamin A alleviates Heat Stress-Induced Damage to Hair Follicle Development in Rex Rabbits. J. Sci. Food Agric. 2022, 102, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Sheiha, A.M.; Abdelnour, S.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Metwally, K.A.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; El-Saadony, M.T. Effects of Dietary Biological or Chemical-Synthesized Nano-Selenium Supplementation on Growing Rabbits Exposed to Thermal Stress. Animals 2020, 10, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosny, N.S.; Hashem, N.M.; Morsy, A.S.; Abo-Elezz, Z.R. Effects of Organic Selenium on the Physiological Response, Blood Metabolites, Redox Status, Semen Quality, and Fertility of Rabbit Bucks Kept under Natural Heat Stress Conditions. Front. Vet. Sci. 2020, 7, 290. [Google Scholar] [CrossRef]
- El-Kholy, M.S.; El-Mekkawy, M.M.; Madkour, M.; Abd El-Azeem, N.; Di Cerbo, A.; Mohamed, L.A.; Alagawany, M.; Selim, D.A. The Role of Different Dietary Zn Sources in Modulating Heat Stress-related Effects on SomeThermoregulatory Parameters of New Zealand White Rabbit Bucks. Anim. Biotechnol. 2021, 23, 1–10. [Google Scholar]
- Goodb, F.; Soliman, F.; Elghalid, O.; Abd El-Hady, A.M. Can copper amino acid chelates reduce the physiological strain of growing rabbits under summer conditions? World Rabbit Sci. 2022, 30, 277–286. [Google Scholar] [CrossRef]
- Al-Sagheer, A.A.; Abdel-Rahman, G.; Elsisi, G.F.; Ayyat, M.S. Comparative Effects of Supplementary Different Copper Forms on Performance, Protein Efficiency, Digestibility of Nutrients, Immune Function and Architecture of Liver and Kidney in Growing Rabbits. Anim. Biotechnol. 2022, 22, 1–11. [Google Scholar] [CrossRef]
- Huang, C.B.; Tang, L.; Guo, Z.Q.; Yan, J.Y.; Xie, X.H.; Lei, M. Effects of Organic Chromium on the Production Performance and Immune Function of Heat-Stressed Rabbits. Chin. J. Anim. Husb. 2017, 53, 93–95. [Google Scholar]
- El-Kholy, K.H.; Shabaan, H.M.A.; Gad-Alla, S.Z.; Abdel-Kafy, E.M.; Ghazal, M.N. Productive and Physiological Responses of New Zealand White Rabbit Males to Dietary Organic Chromium Addition. Egypt. J. Rabbit Sci. 2014, 24, 1–18. [Google Scholar]
- Bassiony, S.S.; Al-Sagheer, A.A.; El-Kholy, M.S.; Elwakeel, E.A.; Helal, A.A.; Alagawany, M. Evaluation of Enterococcus faecium NCIMB 11181 and Clostridium butyricum Probiotic Supplements in Post-Weaning Rabbits Reared under Thermal Stress Conditions. Ital. J. Anim. Sci. 2021, 20, 1232–1243. [Google Scholar] [CrossRef]
- Hegab, I.M.; Attia, E.A.; Hassan, R.A.; El-Azzazi, F.E. Effect of Probiotics on Productive, Physiological and Microbiological Parameters of New Zealand White Rabbits Reared under Hot Summer Conditions. Egypt. Poult. Sci. 2019, 39, 599–614. [Google Scholar] [CrossRef]
- Khalid, A.R.; Yasoob, T.B.; Zhang, Z.; Yu, D.; Feng, J.; Zhu, X.; Hang, S. Supplementation of Moringa oleifera Leaf Powder Orally Improved Productive Performance by Enhancing the Intestinal Health in Rabbits under Chronic Heat Stress. J. Therm. Biol. 2020, 93, 102680. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.R.; Yasoob, T.B.; Zhang, Z.; Zhu, X.; Hang, S. Dietary Moringa oleifera Leaf Powder Improves Jejunal Permeability and Digestive Function by Modulating The Microbiota Composition and Mucosal Immunity in Heat Stressed Rabbits. Environ. Sci. Pollut. Res. Int. 2022, 29, 80952–80967. [Google Scholar] [CrossRef] [PubMed]
- El-Desoky, N.I.; Hashem, N.M.; Elkomy, A.; Abo-Elezz, Z.R. Physiological Response and Semen Quality of Rabbit Bucks Supplemented with Moringa Leaves Ethanolic Extract During Summer Season. Animal 2017, 11, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Amber, K.; Badawy, N.A.; El-Sayd, A.E.A.; Morsy, W.A.; Hassan, A.M.; Dawood, M.A.O. Ginger Root Powder Enhanced the Growth Productivity, Digestibility, and Antioxidative Capacity to Cope with the Impacts of Heat Stress in Rabbits. J. Therm. Biol. 2021, 100, 103075. [Google Scholar] [CrossRef]
- Habeeb, A.A.; El-Darawany, A.A.; Nasr, A.S.; Sharaf, A.K. Impact of Some Medicinal Plants Supplement on Pregnant Rabbits Diet During Hot Summer Season. Res. J. Med. Plants 2019, 13, 145–154. [Google Scholar]
- Ahmed, A.E.; Alkahtani, M.A.; Abdel-Wareth, A.A.A. Thyme Leaves as an Eco-Friendly Feed Additive Improves Both the Productive and Reproductive Performance of Rabbits under Hot Climatic Conditions. Vet. Med. Czech. 2020, 65, 65553–65563. [Google Scholar]
- Abdelnour, S.A.; El-Ratel, I.T.; Peris, S.I.; El-Raghi, A.A.; Fouda, S. Effects of Dietary Thyme essential oil, on Blood Haematobiochemical, Redox Status, Immunological and Reproductive Variables of Rabbit Does Exposed to High Environmental Temperature. Ital. J. Anim. Sci. 2022, 21, 51–61. [Google Scholar] [CrossRef]
- El-Ratel, I.T.; Tag El-Din, T.E.H.; Bedier, M.M. Beneficial Effects of Curcumin as A Native or Nanoparticles Form on Productive Efficiency, Liver and Kidney Functions, Antioxidative Status and Immunity of Heat-Stressed Growing Rabbits. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1778–1787. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Metwally, M.G.E.; Bahgat, L.B.; Naiel, M.A.E. Pumpkin Seed Oil–Supplemented Diets Promoted the Growth Productivity, Antioxidative Capacity, and Immune Response in Heat-Stressed Growing Rabbits. Trop. Anim. Health. Prod. 2023, 55, 55. [Google Scholar] [CrossRef]
- García, M.L.; Argente, M.J. Exposure to High Ambient Temperatures Alters Embryology in Rabbits. Int. J. Biometeorol. 2017, 61, 1555–1560. [Google Scholar] [CrossRef]
- Mutwedu, V.; Nyongesa, A.; Oduma, J.; Kitaa, J.; Mbaria, J. Thermal Stress Causes Oxidative Stress and Physiological Changes in Female Rabbits. J. Therm Biol. 2021, 95, 102780. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Bai, X.; Xie, X.; Chen, G.; Jia, X.; Lei, M.; Li, C.; Lai, S. Negative Effects of Heat Stress on Ovarian Tissue in Female Rabbit. Front. Vet. Sci. 2022, 9, 1009182. [Google Scholar] [CrossRef]
- Somuncu, S.; Cakmak, M.; Dikmen, G.; Akman, H.; Kaya, M. Ischemia-Reperfusion Injury of Rabbit Ovary and Protective Effect of Trapidil: An Experimental Study. Pediatr. Surg. Int. 2008, 24, 315–318. [Google Scholar] [CrossRef]
- Marco-Jiménez, F.; Naturil-Alfonso, C.; Peñaranda, D.S.; Jiménez-Trigos, E.; García-Diego, F.J.; Vicente, J.S. Maternal Exposure to High Temperatures Disrupts OCT4 mRNA Expression of Rabbit Pre-Implantation Embryos and Endometrial Tissue. Reprod. Domest. Anim. 2013, 48, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Zeferino, C.P.; Komiyama, C.M.; Fernandes, S.; Sartori, J.R.; Teixeira, P.S.; Moura, A.S. Carcass and Meat Quality Traits of Rabbits under Heat Stress. Animal 2013, 7, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Maya-Soriano, M.J.; Taberner, E.; Sabes-Alsina, M.; Ramon, J.; Rafel, O.; Tusell, L.; Piles, M.; Lopez-Bejar, M. Daily Exposure to Summer Temperatures Affects the Motile Subpopulation Structure of Epididymal Sperm Cells but Not Male Fertility in an In Vivo Rabbit Model. Theriogenology 2015, 84, 384–389. [Google Scholar] [CrossRef]
- Sabés-Alsina, M.; Tallo-Parra, O.; Mogas, M.T.; Morrell, J.M.; Lopez-Bejar, M. Heat Stress Has an Effect on Motility and Metabolic Activity of Rabbit Spermatozoa. Anim. Reprod. Sci. 2016, 173, 18–23. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Al-Gabri, N.A.; Hashem, N.M.; Gonzalez-Bulnes, A. Supplementation With Proline Improves Haemato-Biochemical and Reproductive Indicators in Male Rabbits Affected by Environmental Heat-Stress. Animals 2021, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Sabés-Alsina, M.; Planell, N.; Torres-Mejia, E.; Taberner, E.; Maya-Soriano, M.J.; Tusell, L.; Ramon, J.; Dalmau, A.; Piles, M.; López-Béjar, M. Daily Exposure to Summer Circadian Cycles Affects Spermatogenesis, But Not Fertility in an in vivo Rabbit Model. Theriogenology 2015, 83, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Wu, Y.; Cao, J.; Qin, Y. Effects of Chronic Heat Stress on The Reproductive Capacity of Male Rex Rabbits. Livest. Sci. 2012, 146, 13–21. [Google Scholar] [CrossRef]
- Jie, Z.; Chao, Y.; Min, L.; Li, T.; Zhang, X.Y.; Xie, X.H. The Effect of Heat Stress on the Reproductive Performance of Rabbits and the Research Progress of Related Heat Shock Proteins. Rabbit Rais. China 2020, 235, 19–22. [Google Scholar]
- Durairajanayagam, D.; Agarwal, A.; Ong, C. Causes, Effects and Molecular Mechanisms of Testicular Heat Stress. Reprod. Biomed. Online 2015, 30, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xie, X.H.; Lei, M.; Tang, L.; Zhang, X.Y.; Yang, C. Research Progress on the Effect of Heat Stress on the Semen Quality of Male Rabbits and its Mechanism. Chin. Rabbit Rais. 2018, 6, 24–28. [Google Scholar]
- Fathi, M.; Abdelsalam, M.; Al-Homidan, I.; Ebeid, T.; El-Zarei, M.; Abou-Emera, O. Effect of Probiotic Supplementation and Genotype on Growth Performance, Carcass Traits, Hematological Parameters and Immunity of Growing Rabbits under Hot Environmental Conditions. Anim. Sci. J. 2017, 88, 1644–1650. [Google Scholar] [CrossRef]
- Ebeid, T.A.; Al-Homidan, I.H.; Fathi, M.M. Physiological and Immunological Benefits of Probiotics and Their Impacts in Poultry Productivity. World’s Poul. Sci. J. 2021, 77, 883–899. [Google Scholar] [CrossRef]
- Ebeid, T.A.; Tůmová, E.; Al-Homidan, I.H.; Ketta, M.; Chodová, D. The Potential Role of Feed Restriction on Productivity, Carcass Composition, Meat Quality, and Muscle Fibre Properties of Growing Rabbits: A Review. Meat Sci. 2022, 191, 108845. [Google Scholar] [CrossRef]
- Blas, J.C.D. Nutritional Impact On Health and Performance in Intensively Reared Rabbits. Animal 2013, 7, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Shi, Y.; Tang, L.; Chen, L.; Fan, H.; Wang, H.; Wang, J.; Jia, X.; Chen, S.; Lai, S. Heat Stress Affects Faecal Microbial and Metabolic Alterations of Rabbits. Front. Microbiol. 2022, 12, 817615. [Google Scholar] [CrossRef]
- Li, F.; Liu, H.; Wu, X.; Liu, M.; Yue, Z.; Liu, L.; Li, F. Copper Modulates Mitochondrial Oxidative Phosphorylation to Enhance Dermal Papilla Cells Proliferation in Rex Rabbits. Int. J. Mol. Sci. 2022, 23, 6209. [Google Scholar] [CrossRef]
- Yasoob, T.B.; Yu, D.; Khalid, A.R.; Zhang, Z.; Zhu, X.; Saad, H.M.; Hang, S. Oral Administration of Moringa oleifera Leaf Powder Relieves Oxidative Stress, Modulates Mucosal Immune Response and Cecal Microbiota After Exposure to Heat Stress in New Zealand White Rabbits. J. Anim. Sci. Biotechnol. 2021, 12, 66. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Kar, I. Heat Stress on Microbiota Composition, Barrier Integrity, and Nutrient Transport in Gut, Production Performance, and its Amelioration in Farm Animals. J. Anim. Sci. Technol. 2021, 63, 211–247. [Google Scholar] [CrossRef] [PubMed]
- Ebeid, T.A.; Zeweil, H.S.; Basyony, M.M.; Dosoky, W.M.; Badry, H. Fortification of Rabbits Diets with Vitamin E or Selenium Affects Growth Performance, Lipid Peroxidation, Oxidative Status and Immune Response in Growing Rabbits. Livest. Sci. 2013, 155, 323–331. [Google Scholar] [CrossRef]
- Jimoh, O.A.; Ayedun, E.S.; Oyelade, W.A.; Oloruntola, O.D.; Daramola, O.T.; Ayodele, S.O.; Omoniyi, I.S. Protective Effect of Soursop (Annona Muricata Linn.) Juice on Oxidative Stress in Heat Stressed Rabbits. J. Anim. Sci. Technol. 2018, 60, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, L.D.; Li, C.Y.; Guo, Z.Q.; Ren, Y.J.; Zheng, J.; Mei, X.L. Effects of Heat Stress on Reproductive Performance, Serum Biochemical Indexes and Reproductive Hormones in Female Rabbit of Qixing. Southwest China J. Agric. Sci. 2021, 34, 1323–1329. [Google Scholar]
- Madkour, M.; Shakweer, W.M.E.; Hashem, N.M.; Aboelazab, O.; Younis, E.; El-Azeem, N.A.; Shourrap, M. Antioxidants Status and Physiological Responses to Early and Late Heat Stress in Two Rabbit Breeds. J. Anim. Physiol. Anim. Nutr. 2023, 107, 298–307. [Google Scholar] [CrossRef]
- Madkour, M.; Aboelenin, M.M.; Younis, E.; Mohamed, M.A.; Hassan, H.; Alagawany, M.; Shourrap, M. Hepatic Acute-Phase Response, Antioxidant Biomarkers and DNA Fragmentation of Two Rabbit Breeds Subjected to Acute Heat Stress. Ital. J. Anim. Sci. 2020, 19, 1558–1566. [Google Scholar] [CrossRef]
- El-Desoky, N.I.; Hashem, N.M.; Gonzalez-Bulnes, A.; Elkomy, A.G.; Abo-Elezz, Z.R. Effects of A Nanoencapsulated Moringa Leaf Ethanolic Extract on The Physiology, Metabolism, and Reproductive Performance of Rabbit Does During Summer. Antioxidants 2021, 10, 1326. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Hassan, M.A.E.; Mohammed, A.K.; Alhimaidi, A.R.; Al-Gabri, N.; Al-Khaldi, K.O.; Swelum, A.A. The Effect of Adding Different Levels of Curcumin and its Nanoparticles to Extender on Post-Thaw Quality of Cryopreserved Rabbit Sperm. Animals 2020, 10, 1508. [Google Scholar] [CrossRef]
- Hassan, F.A.; Elkassas, N.; Salim, I.; El-Medany, S.; Aboelenin, S.M.; Shukry, M.; Taha, A.E.; Peris, S.; Soliman, M.; Mahrose, K. Impacts of Dietary Supplementations of Orange Peel and Tomato Pomace Extracts as Natural Sources for Ascorbic Acid on Growth Performance, Carcass Characteristics, Plasma Biochemicals And Antioxidant Status of Growing Rabbits. Animals 2021, 11, 1688. [Google Scholar] [CrossRef]
- Wang, W.L.; Xu, Y.Q.; Jiang, B.Y. Research Progress of Rabbit Heat Stress. Feed Res. 2021, 44, 140–143. [Google Scholar]
- Anoh, K.U.; Barje, P.P.; Iyeghe-Erakpotobor, G.T.; Akpa, G.N. Growth Performance of Heat Stressed Rabbits Fed Diets Supplemented with Synthetic and Organic Antioxidants. Niger. J. Anim. Prod. 2017, 44, 177–180. [Google Scholar] [CrossRef]
- Daader, A.H.; Al-Sagheer, A.A.; Gabr, H.A.; Abd El-Moniem, E.A. Alleviation of Heat-Stress-Related Physiological Perturbations in Growing Rabbits Using Natural Antioxidants. Spanish J. Agric. Res. 2018, 16, e0610. [Google Scholar] [CrossRef] [Green Version]
- Ewuola, E.O.; Sanni, R.O.; Aina, T.; Oni, O.; Jimoh, O.A. Thermoregulatory Response and Oxidative Stress Indices of Rabbit Bucks Administered Ascorbic Acid and Sodium Bicarbonate in A Humid Tropical Environment. J. Basic Appl. Zool. 2022, 83, 55. [Google Scholar] [CrossRef]
- Petrescu, D.C.; Petrescu-Mag, R.M. Consumer Behaviour Related to Rabbit Meat as Functional Food. World Rabbit Sci. 2018, 26, 321–333. [Google Scholar] [CrossRef]
- Carabaño, R.; Piquer, J.; Menoyo, D.; Badiola, I. The Digestive System of the Rabbit. In Nutrition of the Rabbit, 2nd ed.; De Blas, C., Wiseman, J., Eds.; CAB International: Wallington, UK, 2010; pp. 1–18. [Google Scholar]
- Hashem, N.M.; Abd El-Hady, A.M.; Hassan, O.A. Inclusion of Phytogenic Feed Additives Comparable to Vitamin E in Diet of Growing Rabbits: Effects on Metabolism and Growth. Ann. Agric. Sci. 2017, 62, 161–167. [Google Scholar] [CrossRef]
- Sharaf, A.K.; El-Darawany, A.A.; Nasr, A.S.; Habeeb, A.A.M. Alleviation the Negative Effects of Summer Heat Stress by Adding Selenium With Vitamin E Or AD3E Vitamins Mixture in Drinking Water of Female Rabbits. Biol. Rhythm. Res. 2021, 52, 535–548. [Google Scholar] [CrossRef]
- Sharaf, A.K.; El-Darawany, A.A.; Nasr, A.S.; Habeeb, A.A.M. Recent Techniques for Amelioration the Effect of Heat Stress Conditions on Male Rabbits. Zag. J. Agric. Res. 2019, 46, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of Vitamin A in the Immune System. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Zhao, G.; Zhang, Y. The Effect of Heat Stress on Rabbits and its Nutrition Regulation. Feed Res. 2006, 7, 19–22. [Google Scholar]
- Lebas, F. Vitamins in Rabbit Nutrition: Literature Review and Recommendations. World Rabbit Sci. 2000, 8, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Strychalski, J.; Gugołek, A.; Antoszkiewicz, Z.; Fopp-Bayat, D.; Kaczorek-Łukowska, E.; Snarska, A.; Zwierzchowski, G.; Król-Grzymała, A.; Matusevičius, P. The Effect of the BCO2 Genotype on the Expression of Genes Related to Carotenoid, Retinol, and A-Tocopherol Metabolism in Rabbits Fed A Diet With Aztec Marigold Flower Extract. Int. J. Mol. Sci. 2022, 23, 10552. [Google Scholar] [CrossRef]
- Strychalski, J.; Gugołek, A.; Brym, P.; Antoszkiewicz, Z.; Chwastowska-Siwiecka, I. Polymorphism of the BCO2 Gene and the Content of Carotenoids, Retinol and A-Tocopherol in the Liver and Fat of Rabbits. Braz. J. Anim. Sci. 2019, 48, e20180243. [Google Scholar]
- Strychalski, J.; Gugołek, A.; Brym, P.; Antoszkiewicz, Z. Effect of The Β-Carotene Oxygenase 2 Genotype on the Content of Carotenoids, Retinol and A-Tocopherol in the Liver, Fat and Milk of Rabbit Does, Reproduction Parameters and Kitten Growth. J. Anim. Phys. Anim. Nutr. 2019, 103, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Ewuola, E.O.; Akinyemi, D.E. Semen Characteristics of Rabbit Bucks Orally Administered Exogenous L-Selenomethionine. Annu. Res. Rev. Biol. 2017, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Abdulrashid, M.; Juniper, D.T. Effect of Dietary Protein, Selenium and Temperature Humidity Index on Reproductive Traits of Male Rabbits in a Tropical Environment. J. Anim. Prod. Res. 2016, 28, 61–75. [Google Scholar]
- Swain, P.S.; Rao, S.B.N.; Rajendran, D.; Dominic, G.; Selvaraju, S. Nano Zinc, an Alternative to Conventional Zinc as Animal Feed Supplement: A Review. Anim. Nutr. 2016, 2, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.A.A.; Ragab, M.M.; Ahmed, E.A.M.; Abudabos, A.M.; Ebeid, T.A. Effect of Dietary Zinc-Methionine Supplementation on Growth Performance, Nutrient Utilization, Antioxidative Properties and Immune Response in Broiler Chickens under High Ambient Temperature. J. Appl. Anim. Res. 2018, 46, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Chrastinová, Ľ.; Čobanová, K.; Chrenková, M.; Poláčiková, M.; Formelová, Z.; Lauková, A.; Ondruška, Ľ.; Simonová, M.P.; Strompfová, V.; Bučko, O. High Dietary Levels of Zinc for Young Rabbits. Slovak. J. Anim. Sci. 2015, 48, 57–63. [Google Scholar]
- Kamel, D.A.; Abdel-Khalek, A.E.; Gab, S.A. Effect of Dietary Zinc-Oxide or Nano-Zinc Oxide on Growth Performance, Oxidative Stress, and Immunity of Growing Rabbits under Hot Climate Conditions. J. Anim. Poult. Prod. 2020, 11, 565–571. [Google Scholar] [CrossRef]
- Hassan, F.; Mobarez, S.; Mohamed, M.; Attia, Y.; Mekawy, A.; Mahrose, K. Zinc and/or Selenium Enriched Spirulina as Antioxidants in Growing Rabbit Diets to Alleviate the Deleterious Impacts of Heat Stress During Summer Season. Animals 2021, 11, 756. [Google Scholar] [CrossRef]
- Mateos, G.; Rebollar, P.; De Blas, C. Minerals, Vitamins and Additives. In Nutrition of the Rabbit, 3rd ed.; de Blas, C., Wiseman, J., Eds.; CABI: Oxfordshire, UK, 2020; pp. 126–158. [Google Scholar]
- Itoh, S.; Ozumi, K.; Kim, H.W.; Nakagawa, O.; McKinney, R.D.; Folz, R.J.; Zelko, I.N.; Ushio-Fukai, M.; Fukai, T. Novel Mechanism for Regulation of Extracellular SOD Transcription and Activity by Copper: Role of Antioxidant-1. Free Radic. Biol. Med. 2009, 46, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Peng, G.; Lu, Y.; Wang, K.; Ju, Q.; Ju, Y.; Ouyang, M. Relationship Between Copper and Immunity: The Potential Role of Copper in Tumor Immunity. Front Oncol. 2022, 12, 1019153. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wu, X.; Liu, H.; Zhang, B.; Liu, L.; Li, F. Dietary Copper Supplementation Enhances Lipolysis in Rex Rabbits. J. Trace. Elem. Med. Biol. 2021, 68, 126851. [Google Scholar] [CrossRef]
- Cheng, Y.F.; Zhao, M.L.; Li, X.H.; Liu, L.W.; Wang, H.W.; Geng, G.R. Effects of Dietary Chromium Levels on Testicular Tissue and Serum Reproductive Hormones in Heat-Stressed Male Rabbits. Chin. J. Vet. Med. 2015, 35, 946–948. [Google Scholar]
- Hashem, S.H.; Al-Saadi, M.J. Chromium Piclonat Effects in Diet of Male Rabbits Exposed to Heat Stress on Some Productive Traits and Blood Serum Parameters. Iraq. J. Agric. Sci. 2019, 50, 1100–1108. [Google Scholar]
- Anoh, K.U.; Ayuba, D.; Ozung, P.O.; Udayi, M.A. Physiological Performance of Heat Stressed Growing Rabbits Fed Diets Supplemented With Vitamin Anti-Oxidants and Bicarbonate Buffers. Afr. J. Biotechnol. 2022, 21, 139–145. [Google Scholar] [CrossRef]
- Anoh, K.U.; Archibong, E.E.; Nsa, E.E.; Udoekong, E.C.; Jimmy, N.P. Antioxidant Activities of Vitamin C and Bi-Carbonate Buffers on Hormones Secretion and Serum Metabolites of Heat-Stressed Rabbit Buck. Ann. Res. Rev. Biol. 2022, 37, 59–68. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.S.; Hassan, A.A.; Basyony, M.M.; Abu Hafsa, S.H. Rabbit Growth, Carcass Characteristic, Digestion, Caecalfermentation, Microflora, and Some Blood Biochemical components Affected by Oral Administration of Anaerobicprobiotic (ZAD). Egypt. J. Nutri. Feeds 2018, 21, 693–710. [Google Scholar] [CrossRef] [Green Version]
- Mancini, S.; Paci, G. Probiotics in Rabbit Farming: Growth Performance, Health Status, and Meat Quality. Animals 2021, 11, 3388. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, D.; Yang, M.; Wen, B.; Lai, J.; Zhou, Y.; Sun, H.; Xiong, L.; Wang, J.; Lin, Y.; et al. Probiotic Clostridium butyricum Improves the Growth Performance, Immune Function, and Gut Microbiota of Weaning Rex Rabbits. Probiotics Antimicro. Proteins 2019, 11, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Metzler-Zebeli, B.U.; Zebeli, Q. Gut Function-Enhancing Properties and Metabolic Effects of Dietary Indigestible Sugars in Rodents and Rabbits. Nutrients 2015, 7, 8348–8365. [Google Scholar] [CrossRef] [Green Version]
- Aboelhadid, S.M.; Hashem, S.; Abdel-Kafy, E.S.; Mahrous, L.N.; Farghly, E.M.; Abdel-Baki, A.S.; Al-Quraishy, S.; Kamel, A.A. Prebiotic Supplementation Effect on Escherichia coli and Salmonella Species Associated with Experimentally Induced Intestinal Coccidiosis in Rabbits. PeerJ 2021, 9, e10714. [Google Scholar] [CrossRef] [PubMed]
- Abdelhady, D.H.; El-Abasy, M.A. Effect of Prebiotic and Probiotic on Growth, Immuno-Hematological Responses and Biochemical Parameters of Infected Rabbits With Pasteurella Multocida. Benha Vet. Med. J. 2015, 28, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Dalle Zotte, A.; Celia, C.; Szendrő, Z. Herbs and spices inclusion as feed stuff or additive in growing rabbit diets and as additive in rabbit meat: A review. Livest. Sci. 2016, 189, 82–90. [Google Scholar] [CrossRef]
- Yasoob, T.B.; Khalid, A.R.; Zhang, Z.; Zhu, X.; Hang, S. Liver Transcriptome of Rabbits Supplemented With Oral Moringa oleifera Leaf Powder under Heat Stress Is Associated With Modulation of Lipid Metabolism and Up-Regulation of Genes for Thermo-Tolerance, Antioxidation, and Immunity. Nutr. Res. 2022, 99, 25–39. [Google Scholar] [CrossRef]
- Asghar, M.U.; Rahman, A.; Hayat, Z.; Rafique, M.K.; Badar, I.H.; Yar, M.K.; Ijaz, M. Exploration of Zingiber Officinale Effects on Growth Performance, Immunity and Gut Morphology in Broilers. Braz. J. Biol. 2021, 83, e250296. [Google Scholar] [CrossRef] [PubMed]
- El-Ratel, I.T.; Attia, K.A.H.; El-Raghi, A.A.; Fouda, S.F. Relief of the Negative Effects of Heat Stress on Semen Quality, Reproductive Efficiency and Oxidative Capacity of Rabbit Bucks Using Different Natural Antioxidants. Anim. Biosci. 2021, 34, 844. [Google Scholar] [CrossRef]
- Ajao, B.H.; Ola, S.I. Effects of Dietary Supplementation of Ginger, Garlic and Onion on Semen Profile, Haematological and Serum Antioxidant Status of Rabbit Bucks Raised in The Dry Season of The Humid Tropic. J. Agric. Sci.–Sri Lanka. 2022, 17, 228–240. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Taha, E.M.M.; Südekum, K.H.; Lohakare, J. Thyme Oil Inclusion Levels in A Rabbit Ration: Evaluation of Productive Performance, Carcass Criteria and Meat Quality under Hot Environmental Conditions. Anim. Nutr. 2018, 4, 410–416. [Google Scholar] [CrossRef]
- Abdel-Wareth, A.A.A.; Al-Kahtani, M.A.; Alsyaad, K.M.; Shalaby, F.M.; Saadeldin, I.M.; Alshammari, F.A.; Mobashar, M.; Suleiman, M.H.A.; Ali, A.H.H.; Taqi, M.O.; et al. Combined Supplementation of Nano-Zinc Oxide and Thyme Oil Improves the Nutrient Digestibility and Reproductive Fertility in the Male Californian Rabbits. Animals 2020, 10, 2234. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.M.; Chao, T.Y.; Chang, W.C.; Chang, M.J.; Lee, M.F. Thymol Reduces Oxidative Stress, Aortic Intimal Thickening, and Inflammation-Related Gene Expression in Hyperlipidemic Rabbits. J. Food. Drug Anal. 2016, 24, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Placha, I.; Chrastinova, L.; Laukova, A.; Cobanova, K.; Takacova, J.; Strompfova, V.; Chrenkova, M.; Formelova, Z.; Faix, S. Effect of Thyme Oil on Small Intestine Integrity and Antioxidant Status, Phagocytic Activity and Gastrointestinal Microbiota in Rabbits. Acta Vet. Hung. 2013, 61, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Samy, A.; Hassan, H.M.A.; Abd El-Ghany, F.T.F.; Morsy, S.H. Comparative Study of Curcumin and Garlic Extracts as Antioxidants in Growing Rabbit Diets on Productive Performance and Antioxidant Status. Adv. Anim. Vet. Sci. 2022, 10, 1039–1047. [Google Scholar] [CrossRef]
- El-Kholy, K.H.; Wafa, W.M.; El-Nagar, H.A.; Aboelmagd, A.M.; El-Ratel, I.T. Physiological Response, Testicular Function, and Health Indices of Rabbit Males Fed Diets Containing Phytochemicals Extract under Heat Stress Conditions. J. Adv. Vet. Anim. Res. 2021, 8, 256–265. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Kadasi, A.; Stochmalova, A.; Balazi, A.; Földesiová, M.; Makovicky, P.; Chrenek, P.; Harrath, A.H. Effect of Turmeric on the Viability, Ovarian Folliculogenesis, Fecundity, Ovarian Hormones and Response to Luteinizing Hormone of Rabbits. Animal 2017, 12, 1242–1249. [Google Scholar] [CrossRef]
- Jimoh, O.A.; Daramola, O.T.; Okin-Aminu, H.O.; Ojo, O.A.; Oyeyemi, W.A. Effect of Phytogenic Supplements on the Reproductive Physiology and Metabolic Hormones of Rabbits Exposed to Heat Stress Conditions. J. Therm. Biol. 2023, 112, 103438. [Google Scholar] [CrossRef]
- El-Bolkiny, Y.E.; Monsour, M.A.; Tabl, G.A.; Rabie, H.M.; Kamel, K.I. Effect of Sage and Neem Aqueous Leaf Extracts on Growth, Carcass and Hematological Parameters of Growing APRI Rabbits under Summer and Winter Conditions. J. Rabbit Sci. 2022, 32, 41–58. [Google Scholar] [CrossRef]
- El-Gindy, Y.M.; Zahran, S.M.; Hassan, M.A.; Sabir, S.A. Effect on Physiological Parameters and Semen Quality Upon Oral Administration of Fresh Onion Juice to V-Line Rabbit Buck During Severe Heat Stress. Anim. Biotechnol. 2022, 11, 1–9. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebeid, T.A.; Aljabeili, H.S.; Al-Homidan, I.H.; Volek, Z.; Barakat, H. Ramifications of Heat Stress on Rabbit Production and Role of Nutraceuticals in Alleviating Its Negative Impacts: An Updated Review. Antioxidants 2023, 12, 1407. https://doi.org/10.3390/antiox12071407
Ebeid TA, Aljabeili HS, Al-Homidan IH, Volek Z, Barakat H. Ramifications of Heat Stress on Rabbit Production and Role of Nutraceuticals in Alleviating Its Negative Impacts: An Updated Review. Antioxidants. 2023; 12(7):1407. https://doi.org/10.3390/antiox12071407
Chicago/Turabian StyleEbeid, Tarek A., Hamad S. Aljabeili, Ibrahim H. Al-Homidan, Zdeněk Volek, and Hassan Barakat. 2023. "Ramifications of Heat Stress on Rabbit Production and Role of Nutraceuticals in Alleviating Its Negative Impacts: An Updated Review" Antioxidants 12, no. 7: 1407. https://doi.org/10.3390/antiox12071407
APA StyleEbeid, T. A., Aljabeili, H. S., Al-Homidan, I. H., Volek, Z., & Barakat, H. (2023). Ramifications of Heat Stress on Rabbit Production and Role of Nutraceuticals in Alleviating Its Negative Impacts: An Updated Review. Antioxidants, 12(7), 1407. https://doi.org/10.3390/antiox12071407