

Albumin Redox Modifications Promote Cell Calcification Reflecting the Impact of Oxidative Status on Aortic Valve Disease and Atherosclerosis
 , , , ,
, , , ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
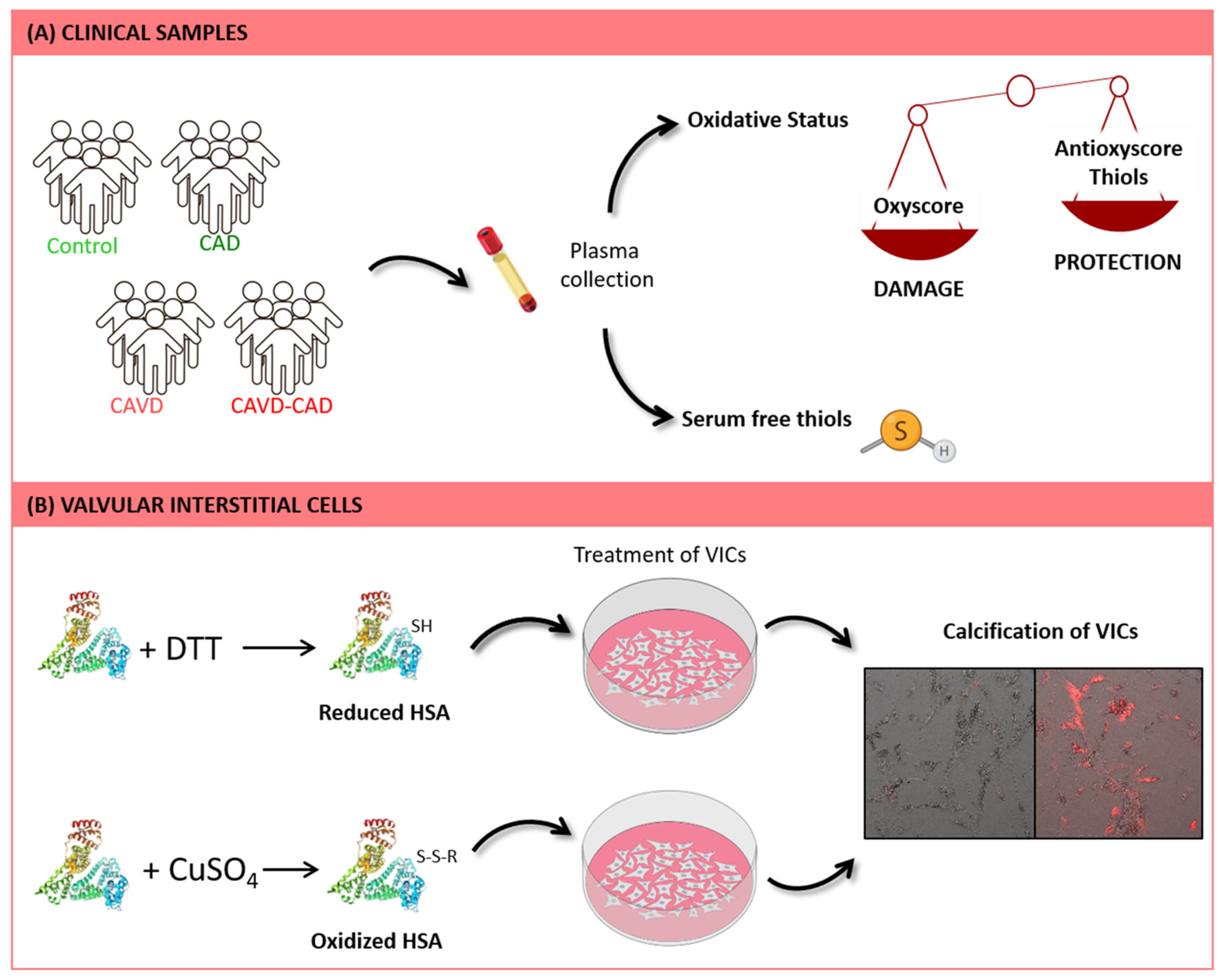
2.1. Patient Selection
2.2. Biomarkers of Oxidative Damage
2.3. Biomarkers of Antioxidant Defense
2.4. Serum Free Thiol Levels
2.5. OxyScore and AntioxyScore
2.6. Valvular Interstitial Cell Culture
2.7. Preparation of Modified HSA
2.8. Alizarin Red Staining
2.9. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
3.2. Markers of Oxidative Damage and of Antioxidant Defenses
3.3. Serum Thiol Levels
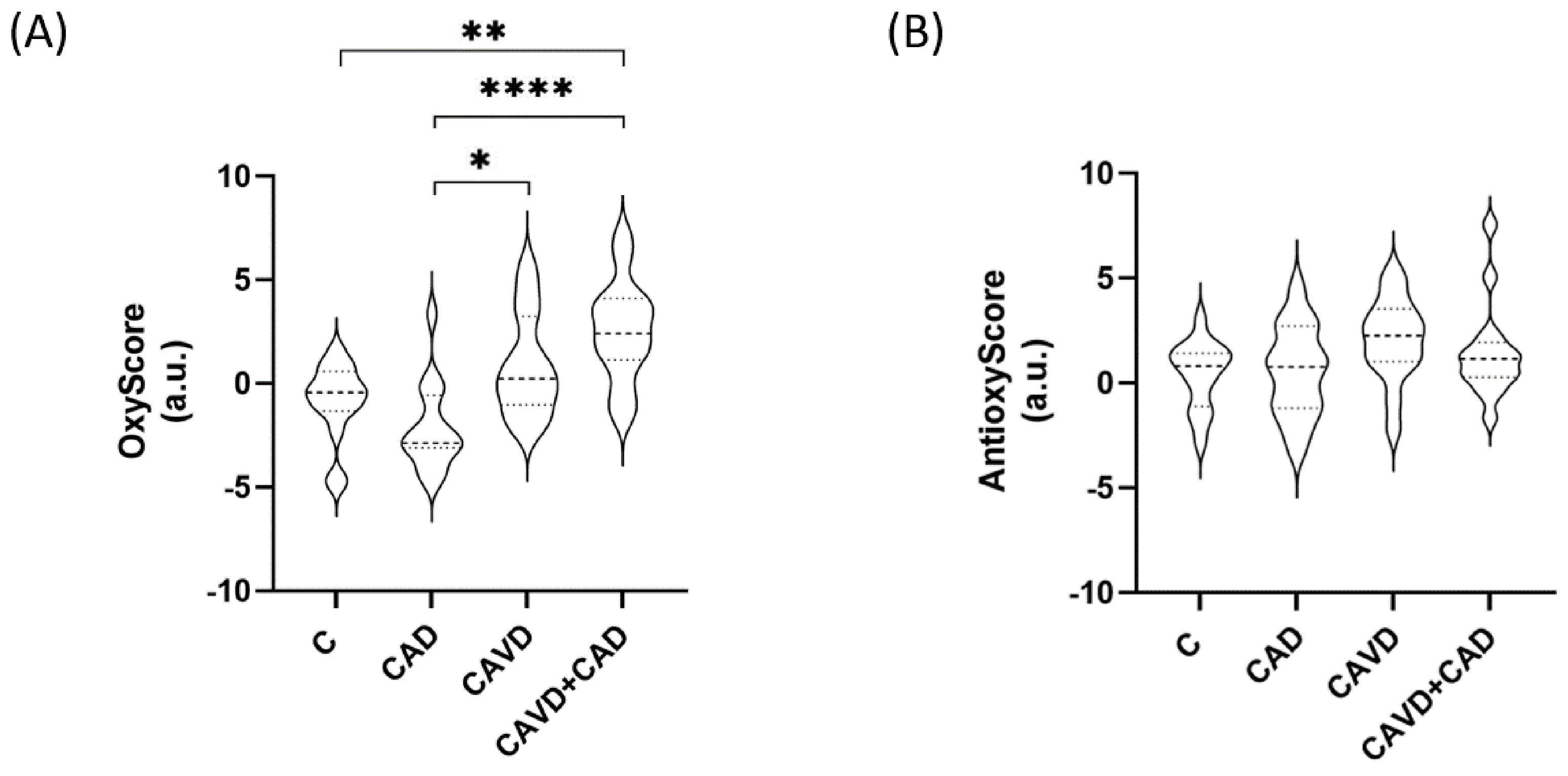
3.4. Global Oxidative Status
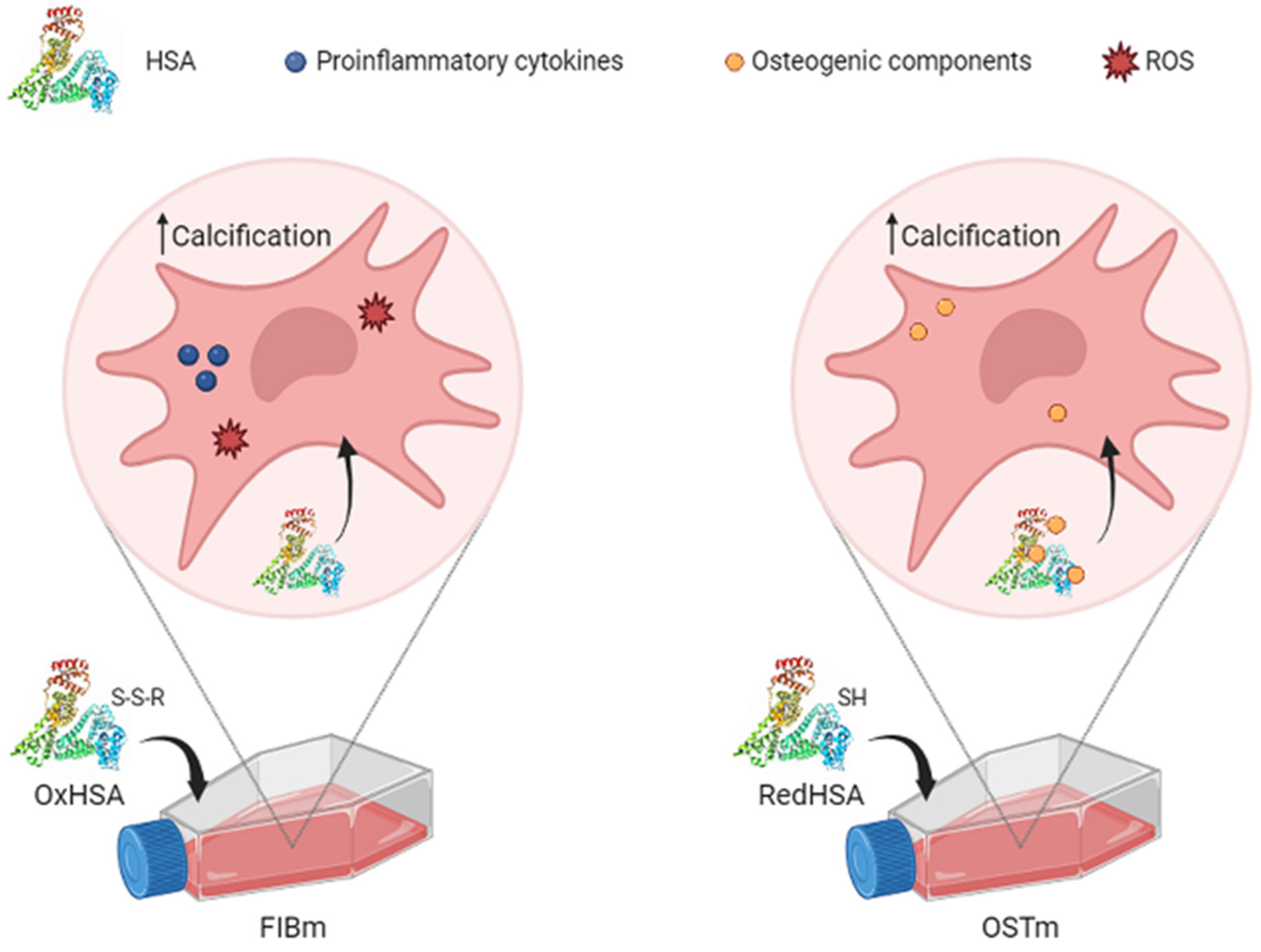
3.5. Cell Calcification on Exposure to HSA
4. Discussion
4.1. Oxidative Status
4.2. How the Human Serum Albumin Redox State Affects Valvular Interstitial Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katz, R.; Wong, N.D.; Kronmal, R.; Takasu, J.; Shavelle, D.M.; Probstfield, J.L.; Bertoni, A.G.; Budoff, M.J.; O’Brien, K.D. Features of the metabolic syndrome and diabetes mellitus as predictors of aortic valve calcification in the Multi-Ethnic Study of Atherosclerosis. Circulation 2006, 113, 2113–2119. [Google Scholar] [CrossRef]
- Ross, R. Atherosclerosis—An inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.M.; Kuusisto, J.; Reichenbach, D.D.; Gown, A.M.; O’Brien, K.D. Characterization of the early lesion of ‘degenerative’ valvular aortic stenosis. Histological and immunohistochemical studies. Circulation 1994, 90, 844–853. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.D.; Reichenbach, D.D.; Marcovina, S.M.; Kuusisto, J.; Alpers, C.E.; Otto, C.M. Apolipoproteins B, (a), and E accumulate in the morphologically early lesion of ‘degenerative’ valvular aortic stenosis. Arter. Thromb. Vasc. Biol. 1996, 16, 523–532. [Google Scholar] [CrossRef] [PubMed]
- van der Wal, A.C.; Becker, A.E.; van der Loos, C.M.; Das, P.K. Site of intimal rupture or erosion of thrombosed coronary atherosclerotic plaques is characterized by an inflammatory process irrespective of the dominant plaque morphology. Circulation 1994, 89, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Naghavi, M.; Libby, P.; Falk, E.; Casscells, S.W.; Litovsky, S.; Rumberger, J.; Badimon, J.J.; Stefanadis, C.; Moreno, P.; Pasterkamp, G.; et al. From vulnerable plaque to vulnerable patient: A call for new definitions and risk assessment strategies: Part I. Circulation 2003, 108, 1664–1672. [Google Scholar] [CrossRef]
- Kimura, Y.; Tsukui, D.; Kono, H. Uric acid in inflammation and the pathogenesis of atherosclerosis. Int. J. Mol. Sci. 2021, 22, 12394. [Google Scholar] [CrossRef] [PubMed]
- Vichova, T.; Motovska, Z. Oxidative stress: Predictive marker for coronary artery disease. Exp. Clin. Cardiol. 2013, 18, e88–e91. [Google Scholar]
- Heistad, D.D.; Wakisaka, Y.; Miller, J.; Chu, Y.; Pena-Silva, R. Novel aspects of oxidative stress in cardiovascular diseases. Circ. J. 2009, 73, 201–207. [Google Scholar] [CrossRef]
- Miller, J.D.; Weiss, R.M.; Heistad, D.D. Calcific aortic valve stenosis: Methods, models, and mechanisms. Circ. Res. 2011, 108, 1392–1412. [Google Scholar] [CrossRef]
- Miller, J.D.; Chu, Y.; Brooks, R.M.; Richenbacher, W.E.; Peña-Silva, R.; Heistad, D.D. Dysregulation of antioxidant mechanisms contributes to increased oxidative stress in calcific aortic valvular stenosis in humans. J. Am. Coll. Cardiol. 2008, 52, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Sung, J.M.; Andreini, D.; Al-Mallah, M.H.; Budoff, M.J.; Cademartiri, F.; Chinnaiyan, K.; Choi, J.H.; Chun, E.J.; Conte, E.; et al. Association between aortic valve calcification progression and coronary atherosclerotic plaque volume progression in the PARADIGM registry. Radiology 2021, 300, 79–86. [Google Scholar] [CrossRef]
- Sastre-Oliva, T.; Corbacho-Alonso, N.; Albo-Escalona, D.; Lopez, J.A.; Lopez-Almodovar, L.F.; Vázquez, J.; Padial, L.R.; Mourino-Alvarez, L.; Barderas, M.G. The influence of coronary artery disease in the development of aortic stenosis and the importance of the albumin redox state. Antioxidants 2022, 11, 317. [Google Scholar] [CrossRef] [PubMed]
- Corbacho-Alonso, N.; Rodriguez-Sanchez, E.; Sastre-Oliva, T.; Mercado-Garcia, E.; Perales-Sanchez, I.; Juarez-Alia, C.; Lopez-Almodovar, L.F.; Padial, L.R.; Tejerina, T.; Mourino-Alvarez, L.; et al. Global oxidative status is linked to calcific aortic stenosis: The differences due to Diabetes Mellitus and the effects of metformin. Antioxidants 2023, 12, 1024. [Google Scholar] [CrossRef] [PubMed]
- Veglia, F.; Cighetti, G.; De Franceschi, M.; Zingaro, L.; Boccotti, L.; Tremoli, E.; Cavalca, V. Age- and gender-related oxidative status determined in healthy subjects by means of OXY-SCORE, a potential new comprehensive index. Biomarkers 2006, 11, 562–573. [Google Scholar] [CrossRef]
- Griendling, K.K.; Touyz, R.M.; Zweier, J.L.; Dikalov, S.; Chilian, W.; Chen, Y.R.; Harrison, D.G.; Bhatnagar, A. Measurement of reactive oxygen species, reactive nitrogen species, and redox-dependent signaling in the cardiovascular system: A scientific statement from the American Heart Association. Circ. Res. 2016, 119, e39–e75. [Google Scholar] [CrossRef] [PubMed]
- Soetan, O.A.; Ajao, F.O.; Ajayi, A.F. Blood glucose lowering and anti-oxidant potential of erythritol: An in vitro and in vivo study. J. Diabetes Metab. Disord. 2023, 22, 1217–1229. [Google Scholar] [CrossRef] [PubMed]
- Jasim, L.M.M.; Homayouni Tabrizi, M.; Darabi, E.; Jaseem, M.M.M. The antioxidant, anti-angiogenic, and anticancer impact of chitosan-coated herniarin-graphene oxide nanoparticles (CHG-NPs). Heliyon 2023, 9, e20042. [Google Scholar] [CrossRef]
- Plácido, A.; do Pais do Amaral, C.; Teixeira, C.; Nogueira, A.; Brango-Vanegas, J.; Alves Barbosa, E.; Moreira, D.C.; Silva-Carvalho, A.; da Silva, M.D.G.; do Nascimento Dias, J.; et al. Neuroprotective effects on microglia and insights into the structure-activity relationship of an antioxidant peptide isolated from Pelophylax perezi. J. Cell. Mol. Med. 2022, 26, 2793–2807. [Google Scholar] [CrossRef]
- Condezo-Hoyos, L.; Rubio, M.; Arribas, S.M.; España-Caparrós, G.; Rodríguez-Rodríguez, P.; Mujica-Pacheco, E.; González, M.C. A plasma oxidative stress global index in early stages of chronic venous insufficiency. J. Vasc. Surg. 2013, 57, 205–213. [Google Scholar] [CrossRef]
- Veglia, F.; Werba, J.P.; Tremoli, E.; Squellerio, I.; Sisillo, E.; Parolari, A.; Minardi, F.; Cavalca, V. Assessment of oxidative stress in coronary artery bypass surgery: Comparison between the global index OXY-SCORE and individual biomarkers. Biomarkers 2009, 14, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Vassalle, C.; Pratali, L.; Boni, C.; Mercuri, A.; Ndreu, R. An oxidative stress score as a combined measure of the pro-oxidant and anti-oxidant counterparts in patients with coronary artery disease. Clin. Biochem. 2008, 41, 1162–1167. [Google Scholar] [CrossRef]
- Ruiz-Hurtado, G.; Condezo-Hoyos, L.; Pulido-Olmo, H.; Aranguez, I.; Del Carmen Gonzalez, M.; Arribas, S.; Cerezo, C.; Segura, J.; Praga, M.; Fernández-Alfonso, M.S.; et al. Development of albuminuria and enhancement of oxidative stress during chronic renin-angiotensin system suppression. J. Hypertens. 2014, 32, 2082–2091; discussion 2091. [Google Scholar] [CrossRef]
- Turell, L.; Radi, R.; Alvarez, B. The thiol pool in human plasma: The central contribution of albumin to redox processes. Free Radic. Biol. Med. 2013, 65, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Adachi, T.; Ashikawa, H.; Hori, M.; Shimozato, T.; Ohtake, H.; Shimizu, S.; Ueyama, J.; Yamada, S. Association between the redox state of human serum albumin and exercise capacity in patients with cardiac disease. Am. J. Cardiol. 2023, 189, 56–60. [Google Scholar] [CrossRef]
- Paramasivan, S.; Adav, S.S.; Ngan, S.C.; Dalan, R.; Leow, M.K.; Ho, H.H.; Sze, S.K. Serum albumin cysteine trioxidation is a potential oxidative stress biomarker of type 2 diabetes mellitus. Sci. Rep. 2020, 10, 6475. [Google Scholar] [CrossRef]
- Sugio, S.; Kashima, A.; Mochizuki, S.; Noda, M.; Kobayashi, K. Crystal structure of human serum albumin at 2.5 A resolution. Protein Eng. 1999, 12, 439–446. [Google Scholar] [CrossRef]
- Nakashima, F.; Shibata, T.; Kamiya, K.; Yoshitake, J.; Kikuchi, R.; Matsushita, T.; Ishii, I.; Giménez-Bastida, J.A.; Schneider, C.; Uchida, K. Structural and functional insights into S-thiolation of human serum albumins. Sci. Rep. 2018, 8, 932. [Google Scholar] [CrossRef]
- Magzal, F.; Sela, S.; Szuchman-Sapir, A.; Tamir, S.; Michelis, R.; Kristal, B. In-vivo oxidized albumin- a pro-inflammatory agent in hypoalbuminemia. PLoS ONE 2017, 12, e0177799. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Morgan, P.E.; Davies, M.J. Quantification of protein modification by oxidants. Free Radic. Biol. Med. 2009, 46, 965–988. [Google Scholar] [CrossRef] [PubMed]
- Saleh, L.; Plieth, C. Total low-molecular-weight antioxidants as a summary parameter, quantified in biological samples by a chemiluminescence inhibition assay. Nat. Protoc. 2010, 5, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Garcia, J.A.; Rodriguez-Sanchez, E.; Aceves-Ripoll, J.; Abarca-Zabalia, J.; Susmozas-Sanchez, A.; Gonzalez Lafuente, L.; Bada-Bosch, T.; Hernandez, E.; Merida-Herrero, E.; Praga, M.; et al. Oxidative status before and after renal replacement therapy: Differences between conventional high flux hemodialysis and on-line hemodiafiltration. Nutrients 2019, 11, 2809. [Google Scholar] [CrossRef]
- Rodriguez-Sanchez, E.; Navarro-Garcia, J.A.; Aceves-Ripoll, J.; Gonzalez-Lafuente, L.; Corbacho-Alonso, N.; Baldan-Martin, M.; Madruga, F.; Alvarez-Llamas, G.; Barderas, M.G.; Ruilope, L.M.; et al. Analysis of global oxidative status using multimarker scores reveals a specific association between renal dysfunction and diuretic therapy in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 1198–1205. [Google Scholar] [CrossRef]
- Rodriguez-Sanchez, E.; Navarro-Garcia, J.A.; Aceves-Ripoll, J.; Gonzalez-Lafuente, L.; Corbacho-Alonso, N.; Martinez, P.; Calvo-Bonacho, E.; Alvarez-Llamas, G.; Barderas, M.G.; Ruilope, L.M.; et al. Lifetime cardiovascular risk is associated with a multimarker score of systemic oxidative status in young adults independently of traditional risk factors. Transl. Res. J. Lab. Clin. Med. 2019, 212, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Latif, N.; Quillon, A.; Sarathchandra, P.; McCormack, A.; Lozanoski, A.; Yacoub, M.H.; Chester, A.H. Modulation of human valve interstitial cell phenotype and function using a Fibroblast Growth Factor 2 formulation. PLoS ONE 2015, 10, e0127844. [Google Scholar] [CrossRef]
- van Engeland, N.C.A.; Bertazzo, S.; Sarathchandra, P.; McCormack, A.; Bouten, C.V.C.; Yacoub, M.H.; Chester, A.H.; Latif, N. Aortic calcified particles modulate valvular endothelial and interstitial cells. Cardiovasc. Pathol. 2017, 28, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Alique, M.; Navalmoral, E.; Noci, M.V.; Bohorquez-Magro, L.; Carracedo, J.; Ramírez, R. Aging-associated oxidized albumin promotes cellular senescence and endothelial damage. Clin. Interv. Aging 2016, 11, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and clinical significance of biomarkers of oxidative stress in humans. Oxidative Med. Cell. Longev. 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Madamanchi, N.R.; Vendrov, A.; Runge, M.S. Oxidative stress and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 29–38. [Google Scholar] [CrossRef]
- Adhikari, R.; Shiwakoti, S.; Ko, J.Y.; Dhakal, B.; Park, S.H.; Choi, I.J.; Kim, H.J.; Oak, M.H. Oxidative stress in calcific aortic valve stenosis: Protective role of natural antioxidants. Antioxidants 2022, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- Romuk, E.; Wojciechowska, C.; Jacheć, W.; Zemła-Woszek, A.; Momot, A.; Buczkowska, M.; Rozentryt, P. Malondialdehyde and Uric Acid as Predictors of Adverse Outcome in Patients with Chronic Heart Failure. Oxidative Med. Cell. Longev. 2019, 2019, 9246138. [Google Scholar] [CrossRef] [PubMed]
- Roest, M.; Voorbij, H.A.; Van der Schouw, Y.T.; Peeters, P.H.; Teerlink, T.; Scheffer, P.G. High levels of urinary F2-isoprostanes predict cardiovascular mortality in postmenopausal women. J. Clin. Lipidol. 2008, 2, 298–303. [Google Scholar] [CrossRef]
- Shishehbor, M.H.; Aviles, R.J.; Brennan, M.L.; Fu, X.; Goormastic, M.; Pearce, G.L.; Gokce, N.; Keaney, J.F., Jr.; Penn, M.S.; Sprecher, D.L.; et al. Association of nitrotyrosine levels with cardiovascular disease and modulation by statin therapy. JAMA 2003, 289, 1675–1680. [Google Scholar] [CrossRef]
- Walter, M.F.; Jacob, R.F.; Jeffers, B.; Ghadanfar, M.M.; Preston, G.M.; Buch, J.; Mason, R.P. Serum levels of thiobarbituric acid reactive substances predict cardiovascular events in patients with stable coronary artery disease: A longitudinal analysis of the PREVENT study. J. Am. Coll. Cardiol. 2004, 44, 1996–2002. [Google Scholar] [CrossRef]
- Leon, M.B.; Piazza, N.; Nikolsky, E.; Blackstone, E.H.; Cutlip, D.E.; Kappetein, A.P.; Krucoff, M.W.; Mack, M.; Mehran, R.; Miller, C.; et al. Standardized endpoint definitions for transcatheter aortic valve implantation clinical trials: A consensus report from the Valve Academic Research Consortium. Eur. Heart J. 2011, 32, 205–217. [Google Scholar] [CrossRef]
- Parenica, J.; Nemec, P.; Tomandl, J.; Ondrasek, J.; Pavkova-Goldbergova, M.; Tretina, M.; Jarkovsky, J.; Littnerova, S.; Poloczek, M.; Pokorny, P.; et al. Prognostic utility of biomarkers in predicting of one-year outcomes in patients with aortic stenosis treated with transcatheter or surgical aortic valve implantation. PLoS ONE 2012, 7, e48851. [Google Scholar] [CrossRef]
- Meijboom, W.B.; Mollet, N.R.; Van Mieghem, C.A.; Kluin, J.; Weustink, A.C.; Pugliese, F.; Vourvouri, E.; Cademartiri, F.; Bogers, A.J.; Krestin, G.P.; et al. Pre-operative computed tomography coronary angiography to detect significant coronary artery disease in patients referred for cardiac valve surgery. J. Am. Coll. Cardiol. 2006, 48, 1658–1665. [Google Scholar] [CrossRef]
- Di Minno, M.N.D.; Poggio, P.; Conte, E.; Myasoedova, V.; Songia, P.; Mushtaq, S.; Cavallotti, L.; Moschetta, D.; Di Minno, A.; Spadarella, G.; et al. Cardiovascular morbidity and mortality in patients with aortic valve calcification: A systematic review and meta-analysis. J. Cardiovasc. Comput. Tomogr. 2019, 13, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Sankaramangalam, K.; Banerjee, K.; Kandregula, K.; Mohananey, D.; Parashar, A.; Jones, B.M.; Jobanputra, Y.; Mick, S.; Krishnaswamy, A.; Svensson, L.G.; et al. Impact of coronary artery disease on 30-day and 1-year mortality in patients undergoing transcatheter aortic valve replacement: A meta-analysis. J. Am. Heart Assoc. 2017, 6, e006092. [Google Scholar] [CrossRef] [PubMed]
- Nagayoshi, Y.; Kawano, H.; Hokamaki, J.; Uemura, T.; Soejima, H.; Kaikita, K.; Sugiyama, S.; Yamabe, H.; Shioji, I.; Sasaki, S.; et al. Differences in oxidative stress markers based on the aetiology of heart failure: Comparison of oxidative stress in patients with and without coronary artery disease. Free Radic. Res. 2009, 43, 1159–1166. [Google Scholar] [CrossRef]
- Xiang, F.; Shuanglun, X.; Jingfeng, W.; Ruqiong, N.; Yuan, Z.; Yongqing, L.; Jun, Z. Association of serum 8-hydroxy-2’-deoxyguanosine levels with the presence and severity of coronary artery disease. Coron. Artery Dis. 2011, 22, 223–227. [Google Scholar] [CrossRef]
- Wang, X.-b.; Cui, N.-h.; Liu, X.n.; Liu, X. Mitochondrial 8-hydroxy-2′-deoxyguanosine and coronary artery disease in patients with type 2 diabetes mellitus. Cardiovasc. Diabetol. 2020, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Di Minno, A.; Turnu, L.; Porro, B.; Squellerio, I.; Cavalca, V.; Tremoli, E.; Di Minno, M.N. 8-Hydroxy-2-Deoxyguanosine levels and cardiovascular disease: A systematic review and meta-analysis of the literature. Antioxid. Redox Signal 2016, 24, 548–555. [Google Scholar] [CrossRef]
- Gorbunova, V.; Seluanov, A.; Mao, Z.; Hine, C. Changes in DNA repair during aging. Nucleic Acids Res. 2007, 35, 7466–7474. [Google Scholar] [CrossRef]
- Redon, J.; Oliva, M.R.; Tormos, C.; Giner, V.; Chaves, J.; Iradi, A.; Sáez, G.T. Antioxidant activities and oxidative stress byproducts in human hypertension. Hypertension 2003, 41, 1096–1101. [Google Scholar] [CrossRef]
- Basile, C.; Fucile, I.; Lembo, M.; Manzi, M.V.; Ilardi, F.; Franzone, A.; Mancusi, C. Arterial hypertension in aortic valve stenosis: A critical update. J. Clin. Med. 2021, 10, 5553. [Google Scholar] [CrossRef]
- Polito, L.; Bortolotti, M.; Battelli, M.G.; Bolognesi, A. Xanthine oxidoreductase: A leading actor in cardiovascular disease drama. Redox Biol. 2021, 48, 102195. [Google Scholar] [CrossRef] [PubMed]
- Battelli, M.G.; Polito, L.; Bolognesi, A. Xanthine oxidoreductase in atherosclerosis pathogenesis: Not only oxidative stress. Atherosclerosis 2014, 237, 562–567. [Google Scholar] [CrossRef]
- Abdulle, A.E.; Bourgonje, A.R.; Kieneker, L.M.; Koning, A.M.; la Bastide-van Gemert, S.; Bulthuis, M.L.C.; Dijkstra, G.; Faber, K.N.; Dullaart, R.P.F.; Bakker, S.J.L.; et al. Serum free thiols predict cardiovascular events and all-cause mortality in the general population: A prospective cohort study. BMC Med. 2020, 18, 130. [Google Scholar] [CrossRef]
- Xuan, Y.; Bobak, M.; Anusruti, A.; Jansen, E.; Pająk, A.; Tamosiunas, A.; Saum, K.U.; Holleczek, B.; Gao, X.; Brenner, H.; et al. Association of serum markers of oxidative stress with myocardial infarction and stroke: Pooled results from four large European cohort studies. Eur. J. Epidemiol. 2019, 34, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Corbacho-Alonso, N.; Baldán-Martín, M.; López, J.A.; Rodríguez-Sánchez, E.; Martínez, P.J.; Mourino-Alvarez, L.; Sastre-Oliva, T.; Cabrera, M.; Calvo, E.; Padial, L.R.; et al. Cardiovascular risk stratification based on oxidative stress for early detection of pathology. Antioxid. Redox Signal 2021, 35, 602–617. [Google Scholar] [CrossRef]
- Ungvari, Z.; Tarantini, S.; Nyúl-Tóth, Á.; Kiss, T.; Yabluchanskiy, A.; Csipo, T.; Balasubramanian, P.; Lipecz, A.; Benyo, Z.; Csiszar, A. Nrf2 dysfunction and impaired cellular resilience to oxidative stressors in the aged vasculature: From increased cellular senescence to the pathogenesis of age-related vascular diseases. GeroScience 2019, 41, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative stress in atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 42. [Google Scholar] [CrossRef]
- Taverna, M.; Marie, A.L.; Mira, J.P.; Guidet, B. Specific antioxidant properties of human serum albumin. Ann. Intensive Care 2013, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, E.; Blache, D. The importance of proteins in defense against oxidation. Antioxid. Redox Signal 2001, 3, 293–311. [Google Scholar] [CrossRef]
- Lim, P.S.; Jeng, Y.; Wu, M.Y.; Pai, M.A.; Wu, T.K.; Liu, C.S.; Chen, C.H.; Kuo, Y.C.; Chien, S.W.; Chen, H.P. Serum oxidized albumin and cardiovascular mortality in normoalbuminemic hemodialysis patients: A cohort study. PLoS ONE 2013, 8, e70822. [Google Scholar] [CrossRef]
- Terawaki, H.; Takada, Y.; Era, S.; Funakoshi, Y.; Nakayama, K.; Nakayama, M.; Ogura, M.; Ito, S.; Hosoya, T. The redox state of albumin and serious cardiovascular incidence in hemodialysis patients. Ther. Apher. Dial. 2010, 14, 465–471. [Google Scholar] [CrossRef]
- Rahmani-Kukia, N.; Abbasi, A.; Pakravan, N.; Hassan, Z.M. Measurement of oxidized albumin: An opportunity for diagnoses or treatment of COVID-19. Bioorg. Chem. 2020, 105, 104429. [Google Scholar] [CrossRef]
- Das, S.; Maras, J.S.; Hussain, M.S.; Sharma, S.; David, P.; Sukriti, S.; Shasthry, S.M.; Maiwall, R.; Trehanpati, N.; Singh, T.P.; et al. Hyperoxidized albumin modulates neutrophils to induce oxidative stress and inflammation in severe alcoholic hepatitis. Hepatology 2017, 65, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, A.; Kubota, K.; Yamada, N.; Tagami, U.; Takehana, K.; Sonaka, I.; Suzuki, E.; Hirayama, K. Identification and characterization of oxidized human serum albumin: A slight structural change impairs its ligand-binding and antioxidant functions. FEBS J. 2006, 273, 3346–3357. [Google Scholar] [CrossRef] [PubMed]
- Oettl, K.; Stauber, R.E. Physiological and pathological changes in the redox state of human serum albumin critically influence its binding properties. Br. J. Pharmacol. 2007, 151, 580–590. [Google Scholar] [CrossRef]
- Wang, W.; Kang, P.M. Oxidative stress and antioxidant treatments in cardiovascular diseases. Antioxidants 2020, 9, 1292. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, L.; Pan, Y.; Wang, X.; Ding, Y.; Zhou, C.; Shah, A.M.; Zhao, G.; Zhang, M. Celastrol alleviates aortic valve calcification via inhibition of NADPH oxidase 2 in valvular interstitial cells. JACC Basic Transl. Sci. 2020, 5, 35–49. [Google Scholar] [CrossRef]
- Tretter, V.; Hochreiter, B.; Zach, M.L.; Krenn, K.; Klein, K.U. Understanding Cellular Redox Homeostasis: A Challenge for Precision Medicine. Int. J. Mol. Sci. 2021, 23, 106. [Google Scholar] [CrossRef]
- Daiber, A.; Hahad, O.; Andreadou, I.; Steven, S.; Daub, S.; Münzel, T. Redox-related biomarkers in human cardiovascular disease—Classical footprints and beyond. Redox Biol. 2021, 42, 101875. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | C (n = 19) | CAD (n = 17) | CAVD (n = 20) | CAVD + CAD (n = 20) | p |
|---|---|---|---|---|---|
| Age | 65.4 ± 8.5 | 80.81 ± 9.2 | 80.70 ± 6.1 | 76.2 ± 7.8 | 0.000 |
| Gender (M/F) | 10/9 | 9/7 | 11/9 | 14/6 | 0.684 |
| %AHT | 47 | 62 | 80 | 55 | 0.183 |
| %Dyslipidemia | 26 | 62 | 45 | 65 | 0.064 |
| %Diabetes | 0 | 0 | 0 | 0 | 1.000 |
| %Smokers | 15 | 0 | 0 | 10 | 0.140 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sastre-Oliva, T.; Corbacho-Alonso, N.; Rodriguez-Sanchez, E.; Mercado-García, E.; Perales-Sanchez, I.; Hernandez-Fernandez, G.; Juarez-Alia, C.; Tejerina, T.; López-Almodóvar, L.F.; Padial, L.R.; et al. Albumin Redox Modifications Promote Cell Calcification Reflecting the Impact of Oxidative Status on Aortic Valve Disease and Atherosclerosis. Antioxidants 2024, 13, 108. https://doi.org/10.3390/antiox13010108
Sastre-Oliva T, Corbacho-Alonso N, Rodriguez-Sanchez E, Mercado-García E, Perales-Sanchez I, Hernandez-Fernandez G, Juarez-Alia C, Tejerina T, López-Almodóvar LF, Padial LR, et al. Albumin Redox Modifications Promote Cell Calcification Reflecting the Impact of Oxidative Status on Aortic Valve Disease and Atherosclerosis. Antioxidants. 2024; 13(1):108. https://doi.org/10.3390/antiox13010108
Chicago/Turabian StyleSastre-Oliva, Tamara, Nerea Corbacho-Alonso, Elena Rodriguez-Sanchez, Elisa Mercado-García, Ines Perales-Sanchez, German Hernandez-Fernandez, Cristina Juarez-Alia, Teresa Tejerina, Luis F. López-Almodóvar, Luis R. Padial, and et al. 2024. "Albumin Redox Modifications Promote Cell Calcification Reflecting the Impact of Oxidative Status on Aortic Valve Disease and Atherosclerosis" Antioxidants 13, no. 1: 108. https://doi.org/10.3390/antiox13010108
APA StyleSastre-Oliva, T., Corbacho-Alonso, N., Rodriguez-Sanchez, E., Mercado-García, E., Perales-Sanchez, I., Hernandez-Fernandez, G., Juarez-Alia, C., Tejerina, T., López-Almodóvar, L. F., Padial, L. R., Sánchez, P. L., Martín-Núñez, E., López-Andrés, N., Ruiz-Hurtado, G., Mourino-Alvarez, L., & Barderas, M. G. (2024). Albumin Redox Modifications Promote Cell Calcification Reflecting the Impact of Oxidative Status on Aortic Valve Disease and Atherosclerosis. Antioxidants, 13(1), 108. https://doi.org/10.3390/antiox13010108