
Anthocyanin Accumulation in Muscadine Berry Skins Is Influenced by the Expression of the MYB Transcription Factors, MybA1, and MYBCS1

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Cell Culture Maintenance
2.2. RNA Extraction, Gel Electrophoresis and cDNA Synthesis
2.3. Isolation of the VrMybA1, and VrMYBCS1 (VrMYB5a) Genes
2.4. Cloning and Sequencing of VrMybA1, and VrMYBCS1 (VrMYB5a) Genes
2.5. Bioinformatics Analysis
2.6. Expression Pattern in Different Berry Stages and in Cell Cultures
2.7. Analysis of Total Anthocyanin in the Berry Skins
3. Results
3.1. Isolation and Cloning of VrMybA1 and VrMYBCS1 Genes from Muscadine Berry Skin Tissues
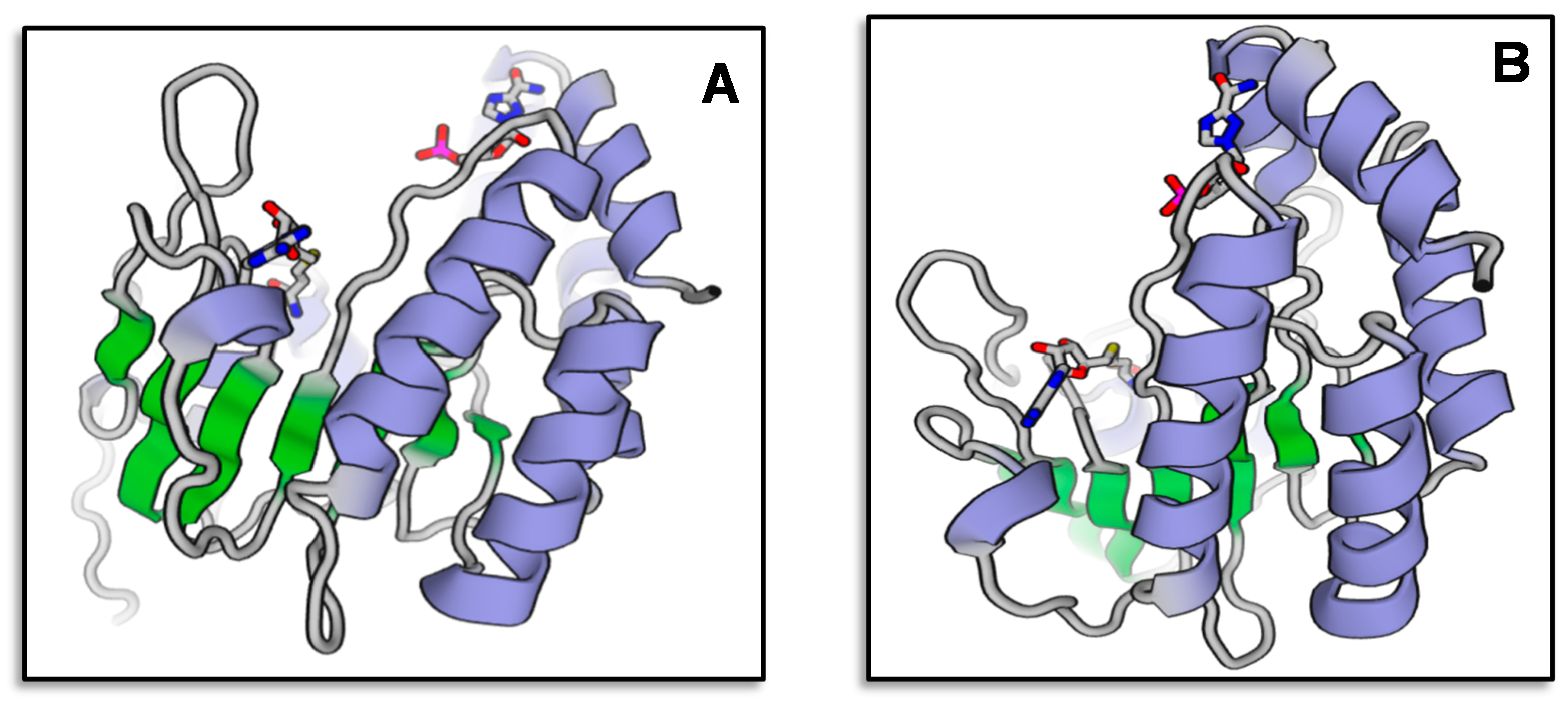
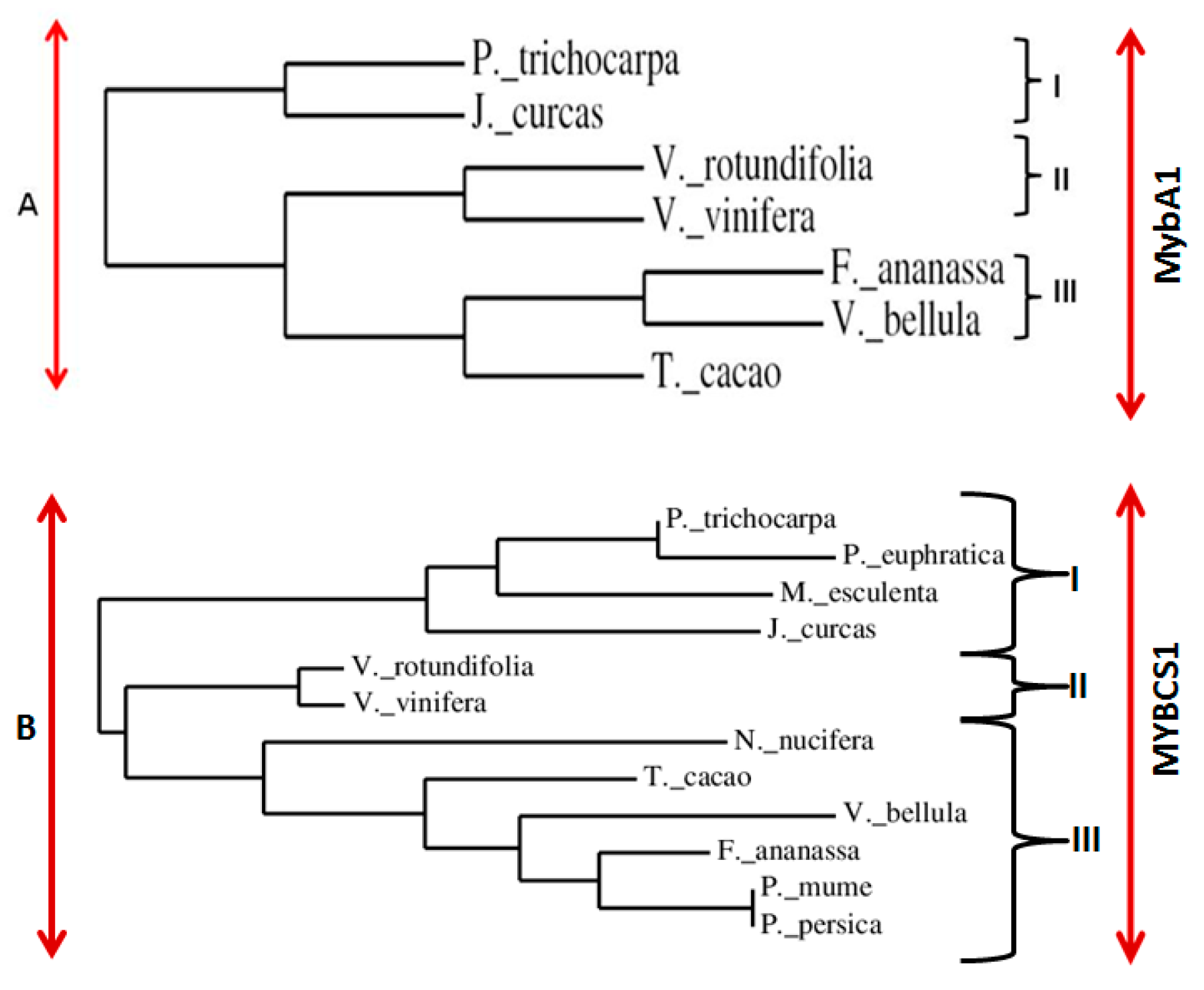
3.2. Sequence Analysis of VrMybA1 and VrMYBCS1
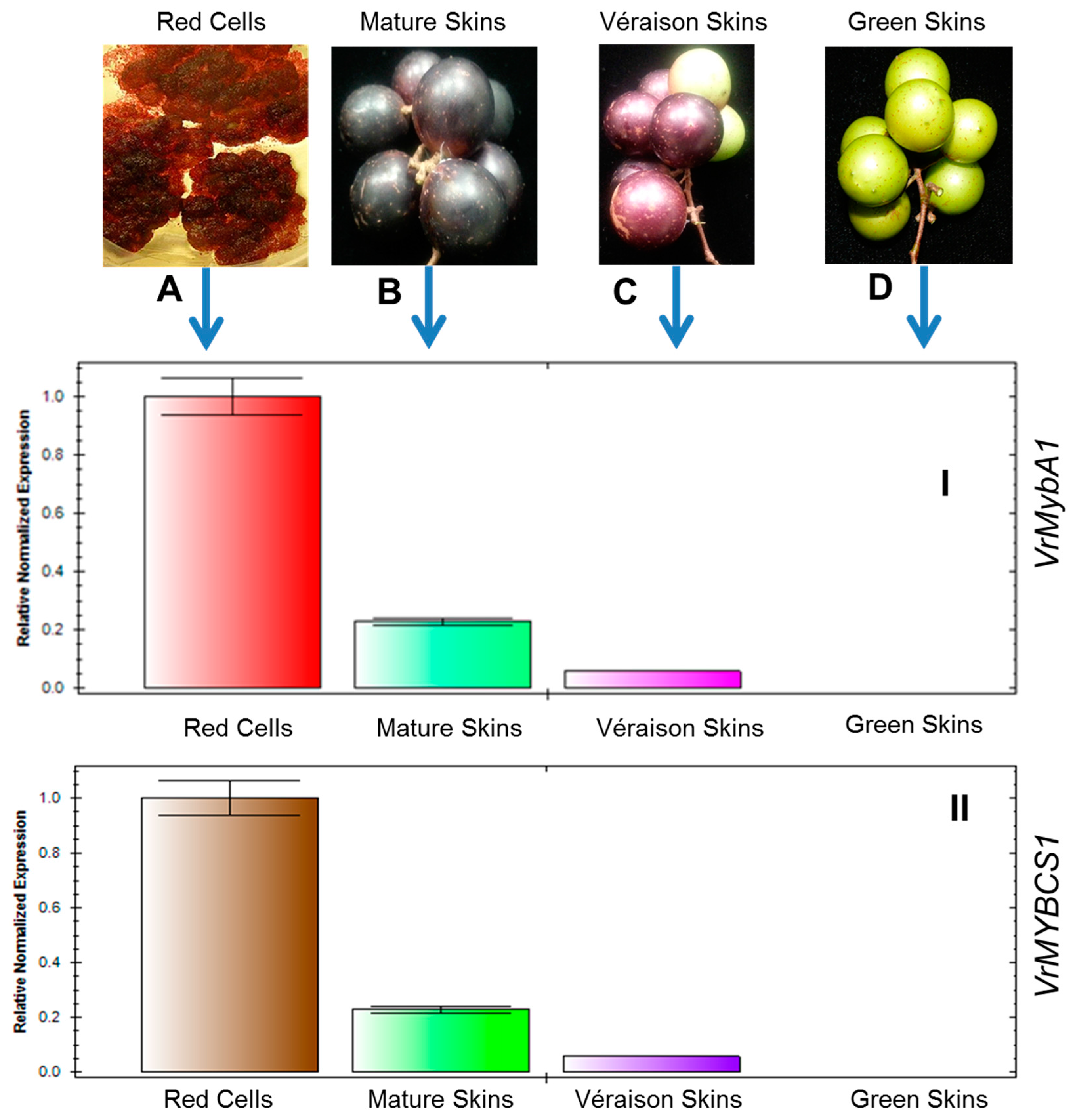
3.3. Expression of VrMybA1 and VrMYBCS1 Genes in Berry Skins Are Influenced by Physiological Changes
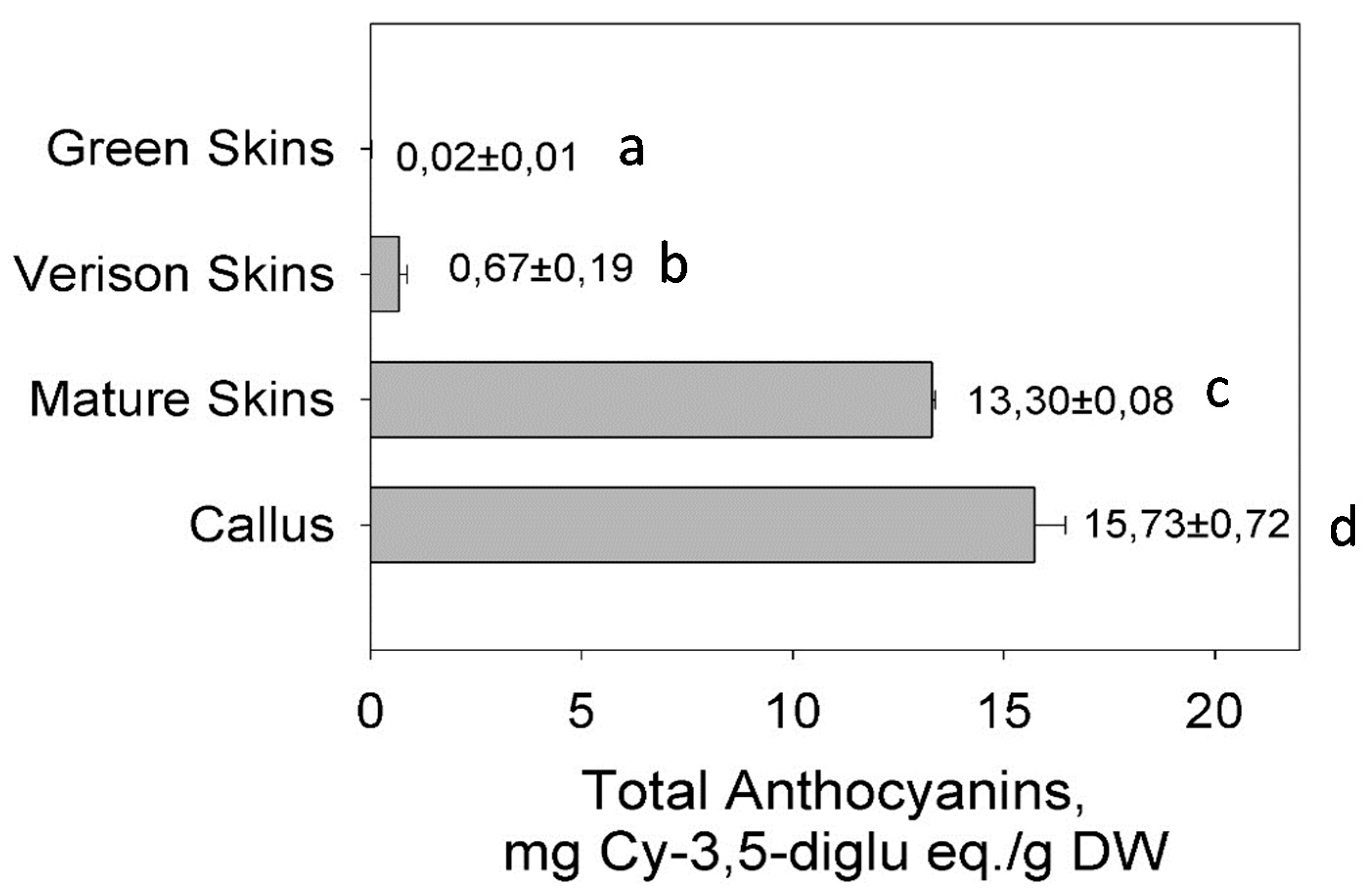
3.4. Anthocyanin Accumulation in the Berry Skin and Cell Cultures
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Georgiev, V.; Ananga, A.; Tsolova, V. Recent advances and uses of grape flavonoids as nutraceuticals. Nutrients 2014, 6, 391–415. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, V.; Ananga, A.; Tsolova, V. Dietary supplements/nutraceuticals made from grapes and wines. In Wine Safety, Consumer Preference, and Human Health; Springer: Cham, Switzerland, 2016; pp. 201–227. [Google Scholar]
- Ananga, A.; Georgiev, V.; Tsolova, V. Manipulation and engineering of metabolic and biosynthetic pathway of plant polyphenols. Curr. Pharm. Des. 2013, 19, 6186–6206. [Google Scholar] [CrossRef] [PubMed]
- Ananga, A.; Phills, B.; Ochieng, J.; Georgiev, V.; Tsolova, V. Production of Anthocyanins in Grape Cell Cultures: A Potential Source of Raw Material for Pharmaceutical, Food, and Cosmetic Industries; InTech Open Access Publisher: Rijeka, Croatia, 2013. [Google Scholar]
- Samuelian, S.K.; Camps, C.; Kappel, C.; Simova, E.P.; Delrot, S.; Colova, V.M. Differential screening of overexpressed genes involved in flavonoid biosynthesis in North American native grapes: “Noble” muscadinia var. and “Cynthiana” aestivalis var. Plant Sci. 2009, 177, 211–221. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Gu, L. Antioxidant capacity, phenolic content, and profiling of phenolic compounds in the seeds, skin, and pulp of Vitis rotundifolia (muscadine grapes) as determined by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2010, 58, 4681–4692. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, B.; Williams, P.; Pace, R.D. Identification of anthocyanins in muscadine grapes with HPLC-ESI-MS. LWT-Food Sci. Technol. 2009, 42, 819–824. [Google Scholar] [CrossRef]
- Hopkins, D.; Purcell, A. Xylella fastidiosa: Cause of Pierce’s disease of grapevine and other emergent diseases. Plant Dis. 2002, 86, 1056–1066. [Google Scholar] [CrossRef]
- Fry, S.; Milholland, R. Response of resistant, tolerant, and susceptible grapevine tissues to invasion by the Pierce’s disease bacterium, Xylella fastidiosa. Phytopathology 1990, 80, 66–69. [Google Scholar] [CrossRef]
- Brownleader, M.D.; Jackson, P.; Mobasheri, A.; Pantelides, A.T.; Sumar, S.; Trevan, M.; Dey, P.M. Molecular aspects of cell wall modifications during fruit ripening. Crit. Rev. Food Sci. Nutr. 1999, 39, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.P.; Davies, C. Molecular biology of grape berry ripening. Aust. J. Grape Wine Res. 2000, 6, 175–188. [Google Scholar] [CrossRef]
- Brummell, D.A.; Dal Cin, V.; Crisosto, C.H.; Labavitch, J.M. Cell wall metabolism during maturation, ripening and senescence of peach fruit. J. Exp. Bot. 2004, 55, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Deytieux, C.; Geny, L.; Lapaillerie, D.; Claverol, S.; Bonneu, M.; Donèche, B. Proteome analysis of grape skins during ripening. J. Exp. Bot. 2007, 58, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Deytieux-Belleau, C.; Geny, L.; Roudet, J.; Mayet, V.; Donèche, B.; Fermaud, M. Grape berry skin features related to ontogenic resistance to Botrytis cinerea. Eur. J. Plant Pathol. 2009, 125, 551–563. [Google Scholar] [CrossRef]
- Antolín, M.C.; Baigorri, H.; Luis, I.D.; Aguirrezábal, F.; Geny, L.; Broquedis, M. ABA during reproductive development in non-irrigated grapevines (Vitis vinifera L. cv. Tempranillo). Aust. J. Grape Wine Res. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Geny, L.; Deytieux, C.; Donèche, B. Importance of Hormonal Profile on the Onset of Ripening in Grape Berries of Vitis vinifera L. In Proceedings of the V International Postharvest Symposium 682, Verona, Italy, 6 June 2004; pp. 99–106.
- Okamoto, G.; Kuwamura, T.; Hirano, K. Effects of water deficit stress on leaf and berry ABA and berry ripening in Chardonnay grapevines (Vitis vinifera). VITIS J. Grapevine Res. 2015, 43, 15–17. [Google Scholar]
- Kanellis, A.; Roubelakis-Angelakis, K. Grape. In Biochemistry of Fruit Ripening; Springer: Dordrecht, The Netherlands, 1993; pp. 189–234. [Google Scholar]
- Keller, M.; Hrazdina, G. Interaction of nitrogen availability during bloom and light intensity during veràison. II. Effects on anthocyanin and phenolic development during grape ripening. Am. J. Enol. Vitic. 1998, 49, 341–349. [Google Scholar]
- Berli, F.J.; Moreno, D.; Piccoli, P.; Hespanhol-Viana, L.E.; Silva, M.F.; Bressan-Smith, R.; Cavagnaro, J.B.; Bottini, R. Abscisic acid is involved in the response of grape (Vitis vinifera L.) cv. Malbec leaf tissues to ultraviolet-B radiation by enhancing ultraviolet-absorbing compounds, antioxidant enzymes and membrane sterols. Plant Cell Environ. 2010, 33, 1–10. [Google Scholar] [PubMed]
- Takos, A.M.; Jaffé, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, S.D.; Pfeiffer, A.; Sivilotti, P.; Degan, M.; Peterlunger, E.; Di Gaspero, G. Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 2007, 30, 1381–1399. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.; Ananga, A.; Georgiev, V.; Ochieng, J.; Cebert, E.; Tsolova, V. Molecular Cloning, Characterization, and Expression Analysis of Flavanone 3-Hydroxylase (F3H) Gene during Muscadine Grape Berry Development. J. Biotechnol. Biomater. 2015, 5, 1–7. [Google Scholar]
- Corbiere, P.; Ananga, A.; Ochieng, J.; Cebert, E.; Tsolova, V. Gene Expression and Molecular Architecture Reveals UDP-Glucose: Flavonoid-3-O-Glucosyltransferase UFGT as a Controller of Anthocyanin Production in Grapes. Jacobs J. Biotechnol. Bioeng. 2015, 2. [Google Scholar] [CrossRef]
- Espley, R.V.; Hellens, R.P.; Putterill, J.; Stevenson, D.E.; Kutty-Amma, S.; Allan, A.C. Red colouration in apple fruit is due to the activity of the MYB transcription factor, MdMYB10. Plant J. 2007, 49, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Holton, T.A.; Cornish, E.C. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Paz-Ares, J. MYB transcription factors in plants. Trends Genet. 1997, 13, 67–73. [Google Scholar] [CrossRef]
- Jin, H.; Martin, C. Multifunctionality and diversity within the plant MYB-gene family. Plant Mol. Biol. 1999, 41, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Schwinn, K.E. Transcriptional regulation of secondary metabolism. Funct. Plant Biol. 2003, 30, 913–925. [Google Scholar] [CrossRef]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Mol, J.; Jenkins, G.; Schäfer, E.; Weiss, D.; Walbot, V. Signal perception, transduction, and gene expression involved in anthocyanin biosynthesis. Crit. Rev. Plant Sci. 1996, 15, 525–557. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis: A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.P.; Mérillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Deluc, L.; Bogs, J.; Walker, A.R.; Ferrier, T.; Decendit, A.; Merillon, J.M.; Robinson, S.P.; Barrieu, F. The transcription factor VvMYB5b contributes to the regulation of anthocyanin and proanthocyanidin biosynthesis in developing grape berries. Plant Physiol. 2008, 147, 2041–2053. [Google Scholar] [CrossRef] [PubMed]
- Lijavetzky, D.; Ruiz-García, L.; Cabezas, J.A.; De Andrés, M.T.; Bravo, G.; Ibáñez, A.; Carreño, J.; Cabello, F.; Ibáñez, J.; Martínez-Zapater, J.M. Molecular genetics of berry colour variation in table grape. Mol. Genet. Genom. 2006, 276, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Lee, E.; Robinson, S.P. Two new grape cultivars, bud sports of Cabernet Sauvignon bearing pale-coloured berries, are the result of deletion of two regulatory genes of the berry colour locus. Plant Mol. Biol. 2006, 62, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Howard, S.; Chen, S.; Wang, Y.; Yu, O.; Kovacs, L.G.; Qiu, W. Berry skin development in Norton grape: Distinct patterns of transcriptional regulation and flavonoid biosynthesis. BMC Plant Biol. 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Colova, V. Synchronized Strains of Subepidermal Cells of Muscadine (muscadine sp.) Grapevine Pericarp for Use as a Sourse of Flavonoids (nutraceuticals). U.S. Patent No. US 2011/0054195 A1, 3 March 2011. [Google Scholar]
- Gollop, R.; Even, S.; Colova-Tsolova, V.; Perl, A. Expression of the grape dihydroflavonol reductase gene and analysis of its promoter region. J. Exp. Bot. 2002, 53, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Oxford University Press: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny. fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001. [Google Scholar] [CrossRef]
- Grotewold, E.; Sainz, M.B.; Tagliani, L.; Hernandez, J.M.; Bowen, B.; Chandler, V.L. Identification of the residues in the Myb domain of maize C1 that specify the interaction with the bHLH cofactor R. Proc. Natl. Acad. Sci. USA 2000, 97, 13579–13584. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.Z.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.A.; Ricke, D.; Lan, T.H.; Presting, G.; Wang, R.; Dunn, M.; Glazebrook, J.; Sessions, A.; Oeller, P.; Varma, H.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. japonica). Science 2002, 296, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.P.; Braun, E.L.; McMullen, M.D.; Grotewold, E. Recently duplicated maize R2R3 MyB genes provide evidence for distinct mechanisms of evolutionary divergence after duplication. Plant Physiol. 2003, 131, 610–620. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, O.S.; Sentenac, A.; Fromageot, P. Specific DNA binding by c-Myb: Evidence for a double helix-turn-helix-related motif. Science 1991, 253, 1140–1143. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Morikawa, S.; Nakamura, H.; Sekikawa, A.; Inoue, T.; Kanai, H.; Sarai, A.; Ishii, S.; Nishimura, Y. Solution structure of a specific DNA complex of the Myb DNA-binding domain with cooperative recognition helices. Cell 1994, 79, 639–648. [Google Scholar] [CrossRef]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oglesby, L.; Ananga, A.; Obuya, J.; Ochieng, J.; Cebert, E.; Tsolova, V. Anthocyanin Accumulation in Muscadine Berry Skins Is Influenced by the Expression of the MYB Transcription Factors, MybA1, and MYBCS1. Antioxidants 2016, 5, 35. https://doi.org/10.3390/antiox5040035
Oglesby L, Ananga A, Obuya J, Ochieng J, Cebert E, Tsolova V. Anthocyanin Accumulation in Muscadine Berry Skins Is Influenced by the Expression of the MYB Transcription Factors, MybA1, and MYBCS1. Antioxidants. 2016; 5(4):35. https://doi.org/10.3390/antiox5040035
Chicago/Turabian StyleOglesby, Lillian, Anthony Ananga, James Obuya, Joel Ochieng, Ernst Cebert, and Violeta Tsolova. 2016. "Anthocyanin Accumulation in Muscadine Berry Skins Is Influenced by the Expression of the MYB Transcription Factors, MybA1, and MYBCS1" Antioxidants 5, no. 4: 35. https://doi.org/10.3390/antiox5040035
APA StyleOglesby, L., Ananga, A., Obuya, J., Ochieng, J., Cebert, E., & Tsolova, V. (2016). Anthocyanin Accumulation in Muscadine Berry Skins Is Influenced by the Expression of the MYB Transcription Factors, MybA1, and MYBCS1. Antioxidants, 5(4), 35. https://doi.org/10.3390/antiox5040035