The Subcellular Distribution of Alpha-Tocopherol in the Adult Primate Brain and Its Relationship with Membrane Arachidonic Acid and Its Oxidation Products

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Serum and Brain Collection
2.3. Preparation of Brain Membranes
2.4. α-Tocopherol Extraction from Brain Regions, Membranes, and Serum
2.5. Fatty Acid Extraction and Protein Determination in Brain Regions and Membranes
2.6. PUFA Oxidation Determination in Brain Tissue
2.7. Statistical Analysis
3. Results
3.1. Tocopherol Concentrations in Rhesus Monkey Stock Diet and Serum
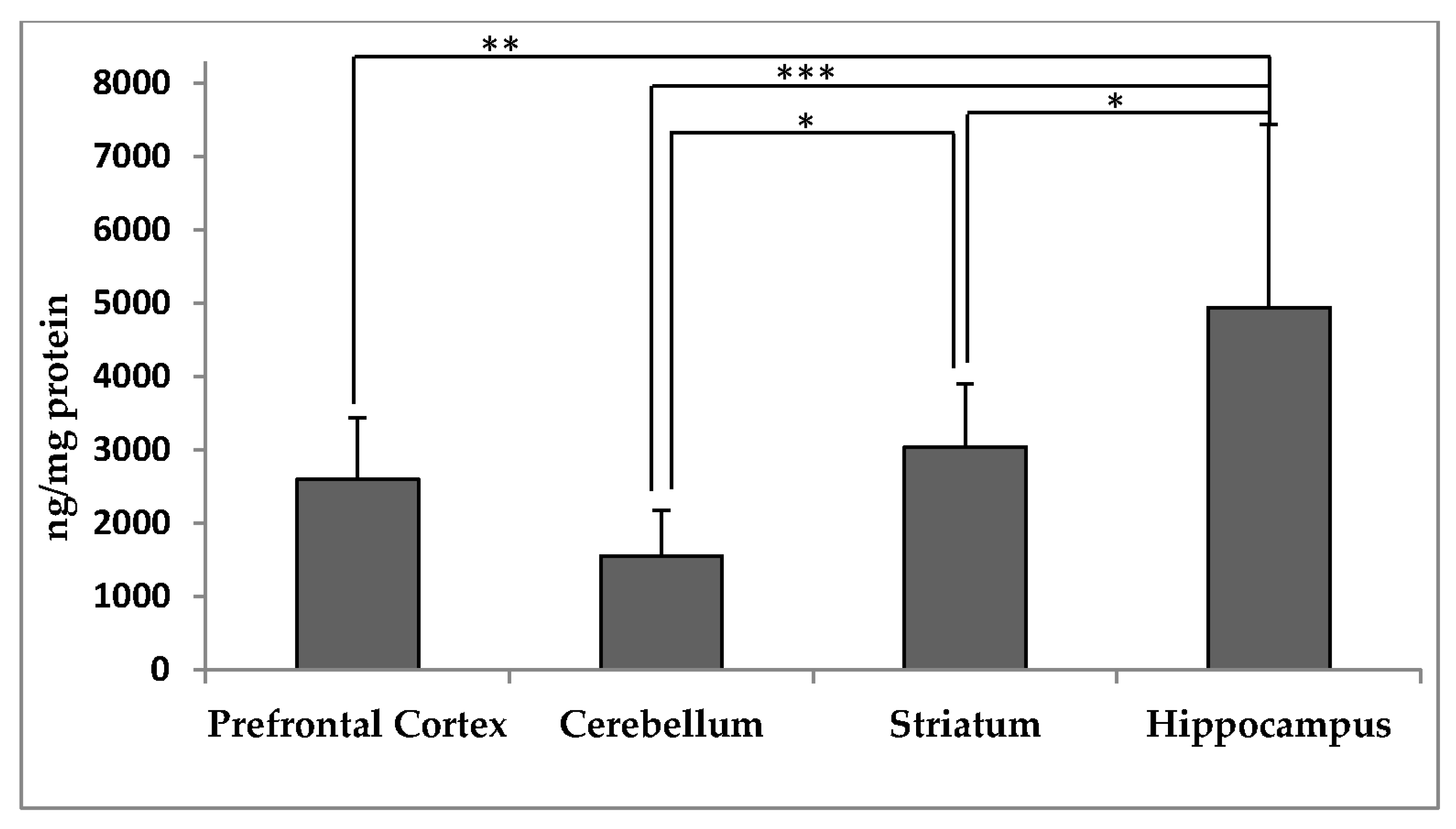
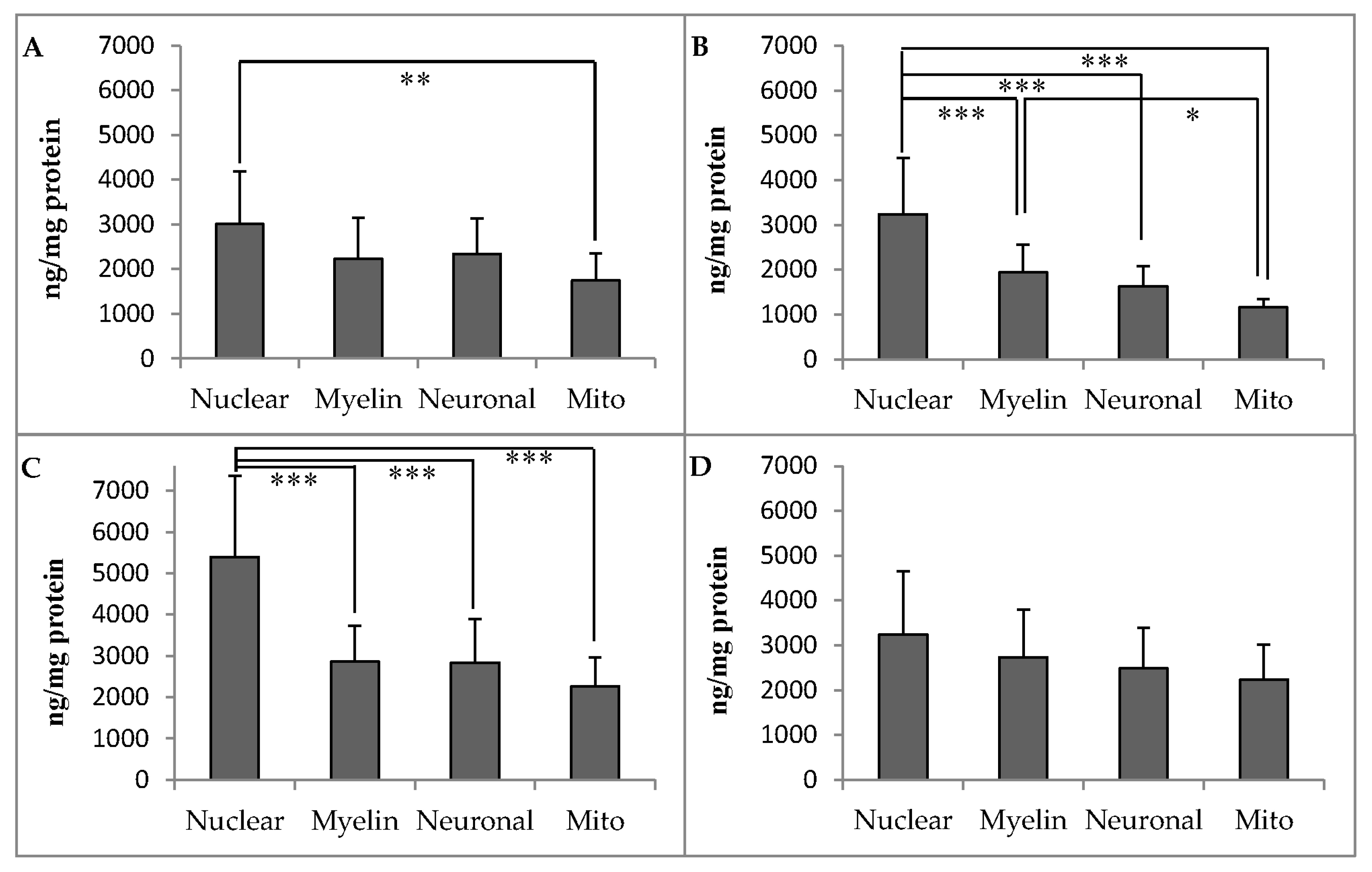
3.2. Distribution of α-Tocopherol in Brain Regions and Membranes of Adult Rhesus Monkeys
3.3. Distribution of Membrane PUFAs in Different Brain Regions of Adult Rhesus Macaques
3.4. Relationship between Membrane α-Tocopherol and PUFA Concentrations in Brain Regions
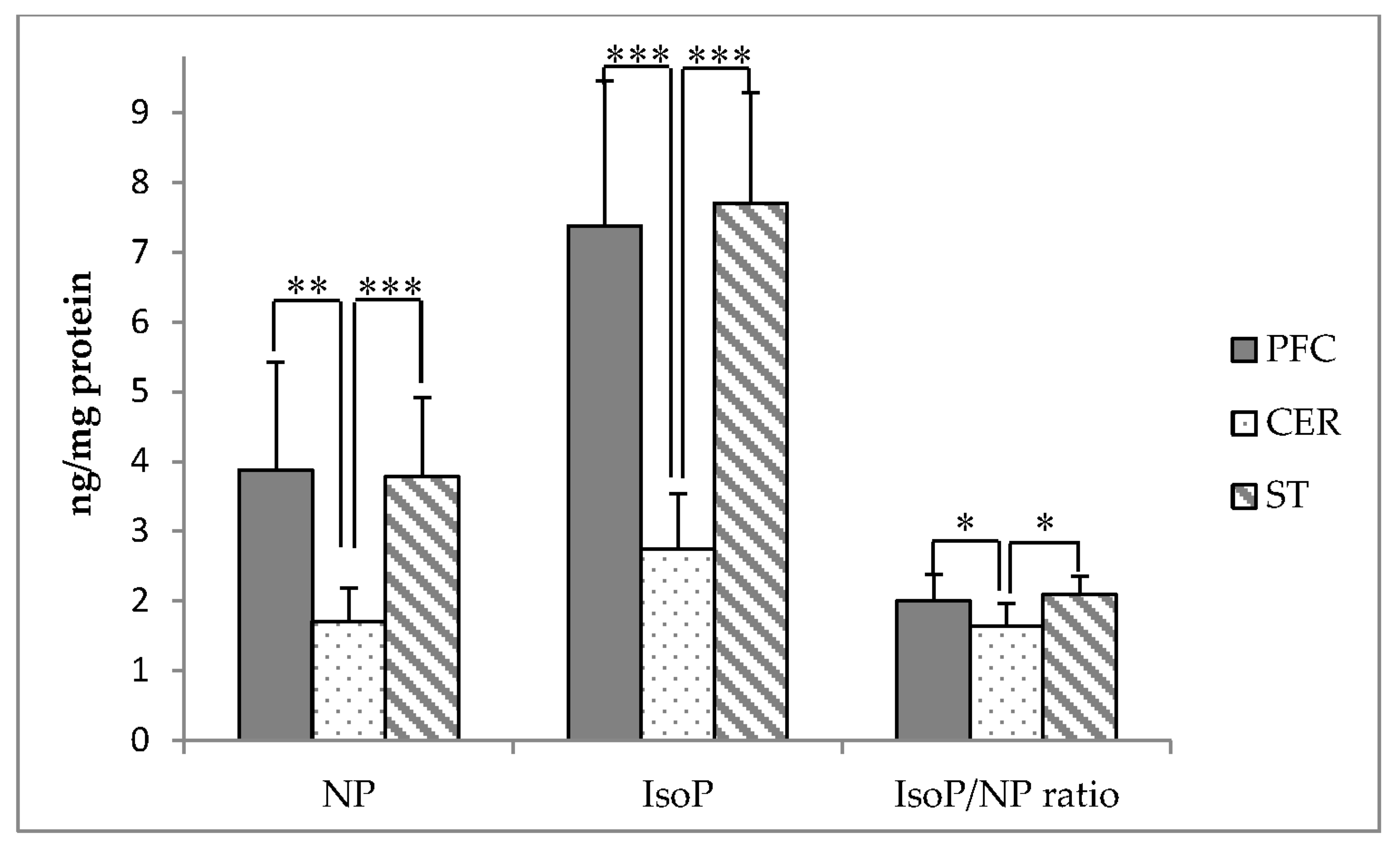
3.5. Relationship between Membrane α-Tocopherol and PUFA Oxidation Products in Brain Regions
4. Discussion
4.1. α-Tocopherol Distribution in Brain Regions of Adult Rhesus Monkeys
4.2. Accumulation of α-Tocopherol in Brain Membranes of Adult Rhesus Monkeys
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Institute of Medicine (US) Panel on Dietary Antioxidants and Related Compounds. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academies Press (US): Washington, DC, USA, 2000; Available online: http://www.ncbi.nlm.nih.gov/books/NBK225483/ (accessed on 1 February 2017).
- Traber, M.G.; Arai, H. Molecular mechanisms of vitamin E transport. Annu. Rev. Nutr. 1999, 19, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, A. Absorption, transport, and tissue delivery of vitamin E. Mol. Asp. Med. 2007, 28, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Grodstein, F.; Chen, J.; Willett, W.C. High-dose antioxidant supplements and cognitive function in community-dwelling elderly women. Am. J. Clin. Nutr. 2003, 77, 975–984. [Google Scholar] [PubMed]
- Mangialasche, F.; Xu, W.; Kivipelto, M.; Costanzi, E.; Ercolani, S.; Pigliautile, M.; Cecchetti, R.; Baglioni, M.; Simmons, A.; Soininen, H.; et al. Tocopherols and tocotrienols plasma levels are associated with cognitive impairment. Neurobiol. Aging 2012, 33, 2282–2290. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.J.; Vishwanathan, R.; Johnson, M.A.; Hausman, D.B.; Davey, A.; Scott, T.M.; Green, R.C.; Miller, L.S.; Gearing, M.; Woodard, J.; et al. Relationship between Serum and Brain Carotenoids, α-Tocopherol, and Retinol Concentrations and Cognitive Performance in the Oldest Old from the Georgia Centenarian Study. J. Aging Res. 2013, 2013, 951786. [Google Scholar] [CrossRef] [PubMed]
- Hensley, K.; Barnes, L.L.; Christov, A.; Tangney, C.; Honer, W.G.; Schneider, J.A.; Bennett, D.A.; Morris, M.C. Analysis of Postmortem Ventricular Cerebrospinal Fluid from Patients with and without Dementia Indicates Association of Vitamin E with Neuritic Plaques and Specific Measures of Cognitive Performance. J. Alzheimers Dis. 2011, 24, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Jiménez, F.J.; de Bustos, F.; Molina, J.A.; Benito-León, J.; Tallón-Barranco, A.; Gasalla, T.; Ortí-Pareja, M.; Guillamón, F.; Rubio, J.C.; Arenas, J.; et al. Cerebrospinal fluid levels of alpha-tocopherol (vitamin E) in Alzheimer’s disease. J. Neural Transm. 1997, 104, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The Role of Oxidative Stress in Neurodegenerative Diseases. Exp. Neurobiol. 2015, 24, 325–340. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Qi, K.; Hall, M.; Deckelbaum, R.J. Long-chain polyunsaturated fatty acid accretion in brain. Curr. Opin. Clin. Nutr. Metab. Care 2002, 5, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Engin, K.N. Alpha-tocopherol: Looking beyond an antioxidant. Mol. Vis. 2009, 15, 855–860. [Google Scholar] [PubMed]
- Quinn, P.J. Is the distribution of alpha-tocopherol in membranes consistent with its putative functions? Biochemistry 2004, 69, 58–66. [Google Scholar] [PubMed]
- Azzi, A.; Gysin, R.; Kempná, P.; Munteanu, A.; Negis, Y.; Villacorta, L.; Visarius, T.; Zingg, J.M. Vitamin E Mediates Cell Signaling and Regulation of Gene Expression. Ann. N. Y. Acad. Sci. 2004, 1031, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Pang, E.; Zingg, J.-M.; Meydani, S.N.; Meydani, M.; Azzi, A. Differential effects of natural and synthetic vitamin E on gene transcription in murine T lymphocytes. Arch. Biochem. Biophys. 2010, 495, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Adolfsson, O.; Lee, C.-K.; Prolla, T.A.; Ordovas, J.; Meydani, S.N. Age and vitamin E-induced changes in gene expression profiles of T cells. J. Immunol. 2006, 177, 6052–6061. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Adolfsson, O.; Lee, C.-K.; Prolla, T.A.; Ordovas, J.; Meydani, S.N. Vitamin E and gene expression in immune cells. Ann. N. Y. Acad. Sci. 2004, 1031, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Quinn, P.J. Vitamin E and its function in membranes. Prog. Lipid Res. 1999, 38, 309–336. [Google Scholar] [CrossRef]
- Vatassery, G.T.; Angerhofer, C.K.; Knox, C.A.; Deshmukh, D.S. Concentrations of vitamin E in various neuroanatomical regions and subcellular fractions, and the uptake of vitamin E by specific areas, of rat brain. Biochim. Biophys. Acta 1984, 792, 118–122. [Google Scholar] [CrossRef]
- Vatassery, G.T.; Smith, W.E.; Quach, H.T. α-Tocopherol in Rat Brain Subcellular Fractions Is Oxidized Rapidly during Incubations with Low Concentrations of Peroxynitrite. J. Nutr. 1998, 128, 152–157. [Google Scholar] [PubMed]
- Perretta, G. Non-Human Primate Models in Neuroscience Research. Scand. J. Lab. Anim. Sci. 2009, 36, 77–85. [Google Scholar]
- Mohn, E.S.; Erdman, J.W.; Kuchan, M.J.; Neuringer, M.; Johnson, E.J. Lutein accumulates in subcellular membranes of brain regions in adult rhesus macaques: Relationship to DHA oxidation products. PLoS ONE 2017, 12, e0186767. [Google Scholar] [CrossRef] [PubMed]
- Muzhingi, T.; Yeum, K.-J.; Russell, R.M.; Johnson, E.J.; Qin, J.; Tang, G. Determination of carotenoids in yellow maize, the effects of saponification and food preparations. Int. J. Vitam. Nutr. Res. 2008, 78, 112–120. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide for the Care and Use of Laboratory Animal, 8th ed.; National Academies Press (US): Washington, DC, USA, 2011; Available online: http://www.ncbi.nlm.nih.gov/books/NBK54050/ (accessed on 15 February 2017).
- Sun, G.Y.; Sun, Y. Phospholipids and acyl groups of synaptosomal and myelin membranes isolated from the cerebral cortex of squirrel monkey (Saimiri sciureus). Biochim. Biophys. Acta 1972, 280, 306–315. [Google Scholar] [CrossRef]
- Sun, G.Y. Phospholipids and acyl groups in subcellular fractions from human cerebral cortex. J. Lipid Res. 1973, 14, 656–663. [Google Scholar] [PubMed]
- Park, J.; Hwang, H.; Kim, M.; Lee-Kim, Y. Effects of dietary fatty acids and vitamin E supplementation on antioxidant vitamin status of the second generation rat brain sections. Korean J. Nutr. 2001, 34, 754–761. [Google Scholar]
- Vishwanathan, R.; Neuringer, M.; Snodderly, D.M.; Schalch, W.; Johnson, E.J. Macular lutein and zeaxanthin are related to brain lutein and zeaxanthin in primates. Nutr. Neurosci. 2013, 16, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.J.; Neuringer, M.; Russell, R.M.; Schalch, W.; Snodderly, D.M. Nutritional manipulation of primate retinas, III: Effects of lutein or zeaxanthin supplementation on adipose tissue and retina of xanthophyll-free monkeys. Investig. Ophthalmol. Vis. Sci. 2005, 46, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Lichtenstein, A.H.; Matthan, N.R.; Jalbert, S.M.; Resteghini, N.A.; Schaefer, E.J.; Ausman, L.M. Novel soybean oils with different fatty acid profiles alter cardiovascular disease risk factors in moderately hyperlipidemic subjects. Am. J. Clin. Nutr. 2006, 84, 497–504. [Google Scholar] [PubMed]
- Walter, M.F.; Blumberg, J.B.; Dolnikowski, G.G.; Handelman, G.J. Streamlined F2-isoprostane analysis in plasma and urine with high-performance liquid chromatography and gas chromatography/mass spectroscopy. Anal. Biochem. 2000, 280, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Arneson, K.O.; Roberts, L.J., 2nd. Measurement of products of docosahexaenoic acid peroxidation, neuroprostanes, and neurofurans. Methods Enzymol. 2007, 433, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Mukaka, M. A guide to appropriate use of Correlation coefficient in medical research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar] [PubMed]
- Craft, N.E.; Haitema, T.B.; Garnett, K.M.; Fitch, K.A.; Dorey, C.K. Carotenoid, tocopherol, and retinol concentrations in elderly human brain. J. Nutr. Health Aging 2004, 8, 156–162. [Google Scholar] [PubMed]
- Kuchan, M.J.; Jensen, S.K.; Johnson, E.J.; Lieblein-Boff, J.C. The naturally occurring α-tocopherol stereoisomer RRR-α-tocopherol is predominant in the human infant brain. Br. J. Nutr. 2016, 116, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Copp, R.P.; Wisniewski, T.; Hentati, F.; Larnaout, A.; Ben Hamida, M.; Kayden, H.J. Localization of alpha-tocopherol transfer protein in the brains of patients with ataxia with vitamin E deficiency and other oxidative stress related neurodegenerative disorders. Brain Res. 1999, 822, 80–87. [Google Scholar] [CrossRef]
- Zimmer, S.; Stocker, A.; Sarbolouki, M.N.; Spycher, S.E.; Sassoon, J.; Azzi, A. A novel human tocopherol-associated protein: cloning, in vitro expression, and characterization. J. Biol. Chem. 2000, 275, 25672–25680. [Google Scholar] [CrossRef] [PubMed]
- Podda, M.; Weber, C.; Traber, M.G.; Packer, L. Simultaneous determination of tissue tocopherols, tocotrienols, ubiquinols, and ubiquinones. J. Lipid Res. 1996, 37, 893–901. [Google Scholar] [PubMed]
- Zingg, J.-M.; Azzi, A. Non-antioxidant activities of vitamin E. Curr. Med. Chem. 2004, 11, 1113–1133. [Google Scholar] [CrossRef] [PubMed]
- Rota, C.; Rimbach, G.; Minihane, A.M.; Stoecklin, E.; Barella, L. Dietary vitamin E modulates differential gene expression in the rat hippocampus: potential implications for its neuroprotective properties. Nutr. Neurosci. 2005, 8, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-K.; Page, G.P.; Kim, K. Differential effect of α- and γ-tocopherol supplementation in age-related transcriptional alterations in heart and brain of B6/C3H F1 mice. J. Nutr. 2008, 138, 1010–1018. [Google Scholar] [PubMed]
- Gohil, K.; Schock, B.C.; Chakraborty, A.A.; Terasawa, Y.; Raber, J.; Farese, R.V., Jr.; Packer, L.; Cross, C.E.; Traber, M.G. Gene expression profile of oxidant stress and neurodegeneration in transgenic mice deficient in alpha-tocopherol transfer protein. Free Radic. Biol. Med. 2003, 35, 1343–1354. [Google Scholar] [CrossRef]
- Berchtold, N.C.; Coleman, P.D.; Cribbs, D.H.; Rogers, J.; Gillen, D.L.; Cotman, C.W. Synaptic genes are extensively downregulated across multiple brain regions in normal human aging and Alzheimer’s disease. Neurobiol. Aging 2013, 34, 1653–1661. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, N.C.; Sabbagh, M.N.; Beach, T.G.; Kim, R.C.; Cribbs, D.H.; Cotman, C.W. Brain gene expression patterns differentiate Mild Cognitive Impairment from normal Aged and Alzheimer Disease. Neurobiol. Aging 2014, 35, 1961–1972. [Google Scholar] [CrossRef] [PubMed]
- Simen, A.A.; Bordner, K.A.; Martin, M.P.; Moy, L.A.; Barry, L.C. Cognitive Dysfunction with Aging and the Role of Inflammation. Ther. Adv. Chronic Dis. 2011, 2, 175–195. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, J.; Harroun, T.; Wassall, S.R.; Stillwell, W.; Katsaras, J. The location and behavior of alpha-tocopherol in membranes. Mol. Nutr. Food Res. 2010, 54, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Raederstorff, D.; Wyss, A.; Calder, P.C.; Weber, P.; Eggersdorfer, M. Vitamin E function and requirements in relation to PUFA. Br. J. Nutr. 2015, 114, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Montine, T.J.; Montine, K.S.; Reich, E.E.; Terry, E.S.; Porter, N.A.; Morrow, J.D. Antioxidants significantly affect the formation of different classes of isoprostanes and neuroprostanes in rat cerebral synaptosomes. Biochem. Pharmacol. 2003, 65, 611–617. [Google Scholar] [CrossRef]
- Reich, E.E.; Montine, K.S.; Gross, M.D.; Roberts, L.J., 2nd; Swift, L.L.; Morrow, J.D.; Montine, T.J. Interactions between Apolipoprotein E Gene and Dietary α-Tocopherol Influence Cerebral Oxidative Damage in Aged Mice. J. Neurosci. 2001, 21, 5993–5999. [Google Scholar] [PubMed]
- Lebold, K.M.; Traber, M.G. Interactions between alpha-tocopherol, polyunsaturated fatty acids, and lipoxygenases during embryogenesis. Free Radic. Biol. Med. 2014, 66, 13–19. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| α-Tocopherol | γ-Tocopherol | |
|---|---|---|
| Stock Diet (µmol/kg) | 23 ± 1.2 | 171 ± 34 |
| Serum (µmol/L) | 20.8 ± 8.3 | 2.5 ± 0.6 |
| Region | Nuclear | Myelin | Neuronal | Mitochondrial | |
|---|---|---|---|---|---|
| Prefrontal Cortex | Arachidonic Acid | 123.5 ± 30.0 a | 99.3 ± 30.5 b | 96.44 ± 20.0 b | 28.0 ± 16.3 c |
| PUFA n-6 | 291.7 ± 77.5 a | 204.9 ± 48.9 b | 201.6 ± 63.5 b | 49.2 ± 27.6 c | |
| Docosahexaenoic acid | 151.9 ± 37.2 a | 180.6 ± 48.9 a | 175.8 ± 54.8 a | 35.3 ± 19.2 b | |
| PUFA n-3 | 164.2 ± 44.6 a | 187.6 ± 51.7 a | 182.9 ± 56.8 a | 36.2 ± 19.8 b | |
| Total PUFA | 455.9 ± 120.6 a | 392.4 ± 99.9 a | 384.5 ± 120.1 a | 85.4 ± 47.1 b | |
| Cerebellum | Arachidonic Acid | 91.4 ± 19.1 a | 84.4 ± 32.8 a | 95.9 ± 20.4 a | 15.1 ± 5.8 b |
| PUFAn-6 | 213.0 ± 40.2 a | 166.1 ± 59.3 b | 187.6 ± 38.0 a,b | 30.3 ± 11.4 c | |
| Docosahexaenoic acid | 136.0 ± 26.5 a | 178.2 ± 59.9 b | 199.8 ± 39.7 b | 27.8 ± 10.2 c | |
| PUFA n-3 | 148.8 ± 30.0 a | 185.9 ± 63.3 b | 207.3 ± 40.9 b | 28.7 ± 10.5 c | |
| Total PUFA | 361.8 ± 69.4 a | 352.0 ± 121.4 a | 394.9 ± 76.7 a | 59.0 ± 21.9 b | |
| Striatum | Arachidonic Acid | 221.5 ± 65.4 a | 156.2 ± 20.5 b | 143.9 ± 26.5 b | 30.5 ± 13.3 c |
| PUFA n-6 | 525.2 ± 177.7 a | 315.5 ± 46.4 b | 289.0 ± 47.6 b | 52.9 ± 22.1 c | |
| Docosahexaenoic acid | 196.9 ± 36.1 a | 257.7 ± 35.0 b | 201.0 ± 37.2 a | 37.3 ± 17.4 c | |
| PUFA n-3 | 224.0 ± 43.9 a | 268.7 ± 36.7 b | 211.7 ± 38.5 a | 38.4 ± 17.8 c | |
| Total PUFA | 749.2 ± 217.5 a | 584.2 ± 81.6 b | 500.7 ± 85.1 c | 91.4 ± 39.6 d | |
| Hippocampus | Arachidonic Acid | 208.0 ± 54.6 a,b | 247.1 ± 119.7 a | 170.6 ± 50.4 b | 21.6 ± 6.6 c |
| PUFA n-6 | 452.0 ± 126.0 a | 519.7 ± 226.7 a | 340.0 ± 104.9 b | 37.0 ± 10.9 c | |
| Docosahexaenoic acid | 144.2 ± 40.9 a | 325.2 ± 118.7 b | 201.5 ± 63.4 c | 16.7 ± 5.5 d | |
| PUFA n-3 | 191.6 ± 54.2 a | 347.7 ± 127.5 b | 219.4 ± 69.7 a | 18.2 ± 5.5 c | |
| Total PUFA | 643.6 ± 177.5 a,b | 867.5 ± 351.2 a | 559.5 ± 173.8 b | 55.2 ± 16.1 c |
| Region | Nuclear | Myelin | Neuronal | Mitochondrial | |
|---|---|---|---|---|---|
| Prefrontal Cortex | Arachidonic acid | 0.68 ** | 0.69 ** | 0.78 ** | -- |
| PUFA n-6 | 0.70 ** | 0.61 | 0.82 ** | -- | |
| Docosahexaenoic acid | 0.70 ** | 0.55 | 0.94 *** | -- | |
| PUFA n-3 | 0.71 ** | 0.54 | 0.94 *** | -- | |
| Total PUFA | 0.71 ** | 0.58 | 0.91 *** | -- | |
| Cerebellum | Arachidonic acid | 0.78 ** | 0.43 | 0.52 | 0.53 |
| PUFA n-6 | 0.85 *** | 0.47 | 0.58 | 0.57 | |
| Docosahexaenoic acid | 0.61 | 0.40 | 0.63 | 0.56 | |
| PUFA n-3 | 0.70 ** | 0.41 | 0.62 | 0.56 | |
| Total PUFA | 0.76 ** | 0.44 | 0.60 | 0.57 | |
| Striatum | Arachidonic acid | 0.89 *** | 0.81 ** | 0.86 *** | 0.80 ** |
| PUFA n-6 | 0.81 ** | 0.76 ** | 0.84 *** | 0.75 ** | |
| Docosahexaenoic acid | 0.89 *** | 0.81 ** | 0.80 ** | 0.80 ** | |
| PUFA n-3 | 0.91 *** | 0.80 ** | 0.79 ** | 0.79 ** | |
| Total PUFA | 0.85 *** | 0.79 ** | 0.83 ** | 0.77 ** | |
| Hippocampus | Arachidonic acid | 0.60 | 0.71 ** | 0.48 | 0.61 |
| PUFA n-6 | 0.64 | 0.79 ** | 0.50 | 0.58 | |
| Docosahexaenoic acid | 0.61 | 0.70 ** | 0.57 | 0.79 ** | |
| PUFA n-3 | 0.67 | 0.70 ** | 0.56 | 0.77 ** | |
| Total PUFA | 0.66 | 0.76 ** | 0.53 | 0.66 |
| Total | Nuclear | Myelin | Neuronal | Mitochondrial | |
|---|---|---|---|---|---|
| Neuroprostanes | |||||
| Prefrontal Cortex | −0.66 * | -- | -- | -- | -- |
| Cerebellum | -- | -- | -- | -- | -- |
| Striatum | −0.43 | −0.62 * | -- | -- | -- |
| Isoprostanes | |||||
| Prefrontal Cortex | −0.74 ** | −0.47 | -- | -- | -- |
| Cerebellum | -- | −0.66 * | -- | -- | −0.61 * |
| Striatum | −0.73 ** | -- | −0.64 * | -0.50 | -- |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohn, E.S.; Kuchan, M.J.; Erdman, J.W.; Neuringer, M.; Matthan, N.R.; Chen, C.-Y.O.; Johnson, E.J. The Subcellular Distribution of Alpha-Tocopherol in the Adult Primate Brain and Its Relationship with Membrane Arachidonic Acid and Its Oxidation Products. Antioxidants 2017, 6, 97. https://doi.org/10.3390/antiox6040097
Mohn ES, Kuchan MJ, Erdman JW, Neuringer M, Matthan NR, Chen C-YO, Johnson EJ. The Subcellular Distribution of Alpha-Tocopherol in the Adult Primate Brain and Its Relationship with Membrane Arachidonic Acid and Its Oxidation Products. Antioxidants. 2017; 6(4):97. https://doi.org/10.3390/antiox6040097
Chicago/Turabian StyleMohn, Emily S., Matthew J. Kuchan, John W. Erdman, Martha Neuringer, Nirupa R. Matthan, Chung-Yen Oliver Chen, and Elizabeth J. Johnson. 2017. "The Subcellular Distribution of Alpha-Tocopherol in the Adult Primate Brain and Its Relationship with Membrane Arachidonic Acid and Its Oxidation Products" Antioxidants 6, no. 4: 97. https://doi.org/10.3390/antiox6040097
APA StyleMohn, E. S., Kuchan, M. J., Erdman, J. W., Neuringer, M., Matthan, N. R., Chen, C. -Y. O., & Johnson, E. J. (2017). The Subcellular Distribution of Alpha-Tocopherol in the Adult Primate Brain and Its Relationship with Membrane Arachidonic Acid and Its Oxidation Products. Antioxidants, 6(4), 97. https://doi.org/10.3390/antiox6040097