The Cell Culture Medium Affects Growth, Phenotype Expression and the Response to Selenium Cytotoxicity in A549 and HepG2 Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culturing and Proliferation Rate
2.3. Viability Measurements
2.4. Intracellular and Extracellular Thiols
2.5. Determination of Protein Concentration
2.6. Thioredoxin Reductase 1 (TrxR1) Expression and Activity
2.7. Western Blot
2.8. Immunocytochemical Staining
2.9. Statistical Analysis
3. Results
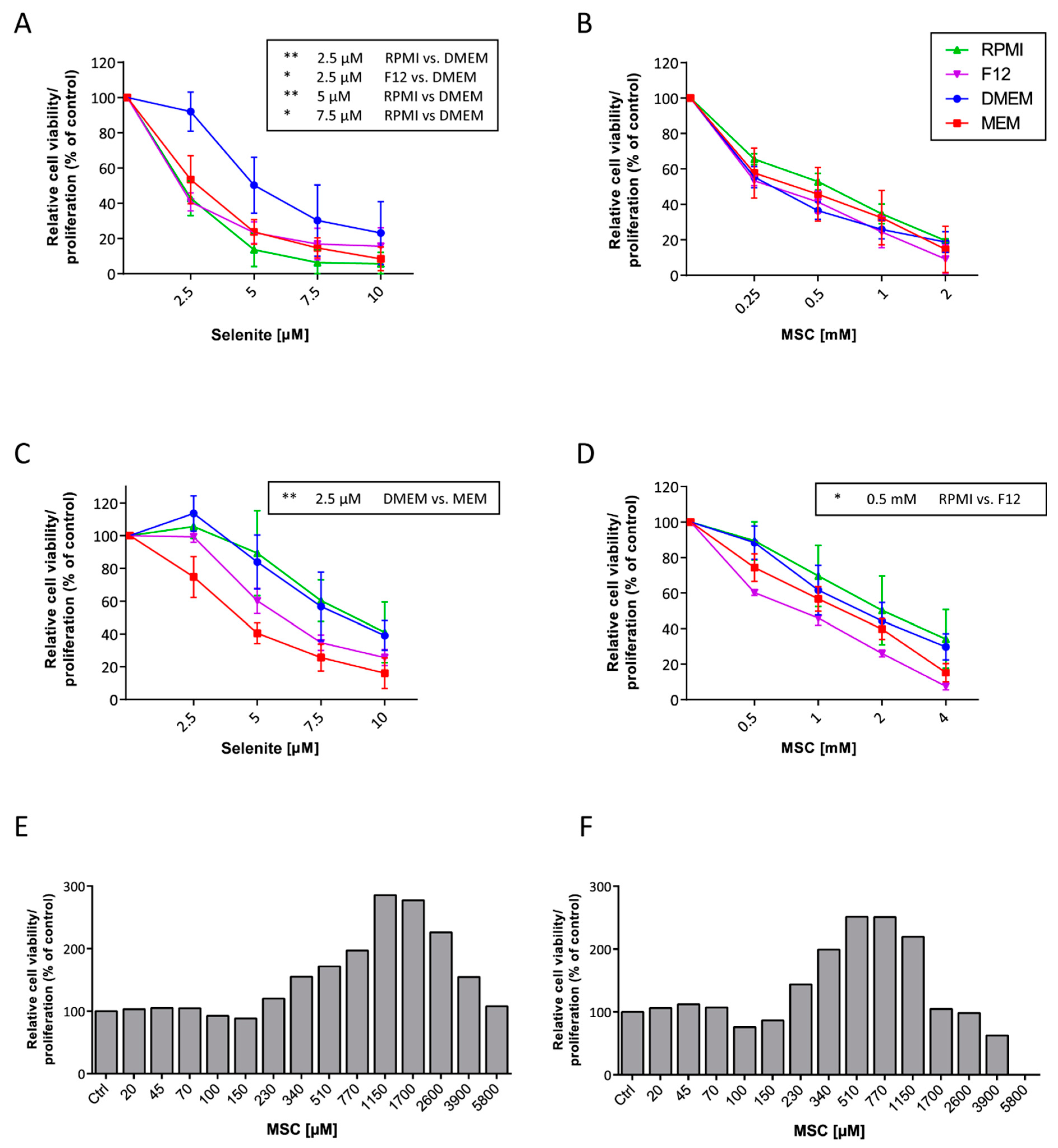
3.1. Cell Culture Media Influence Selenium Cytotoxicity
3.2. MSC Interacts with WST-1
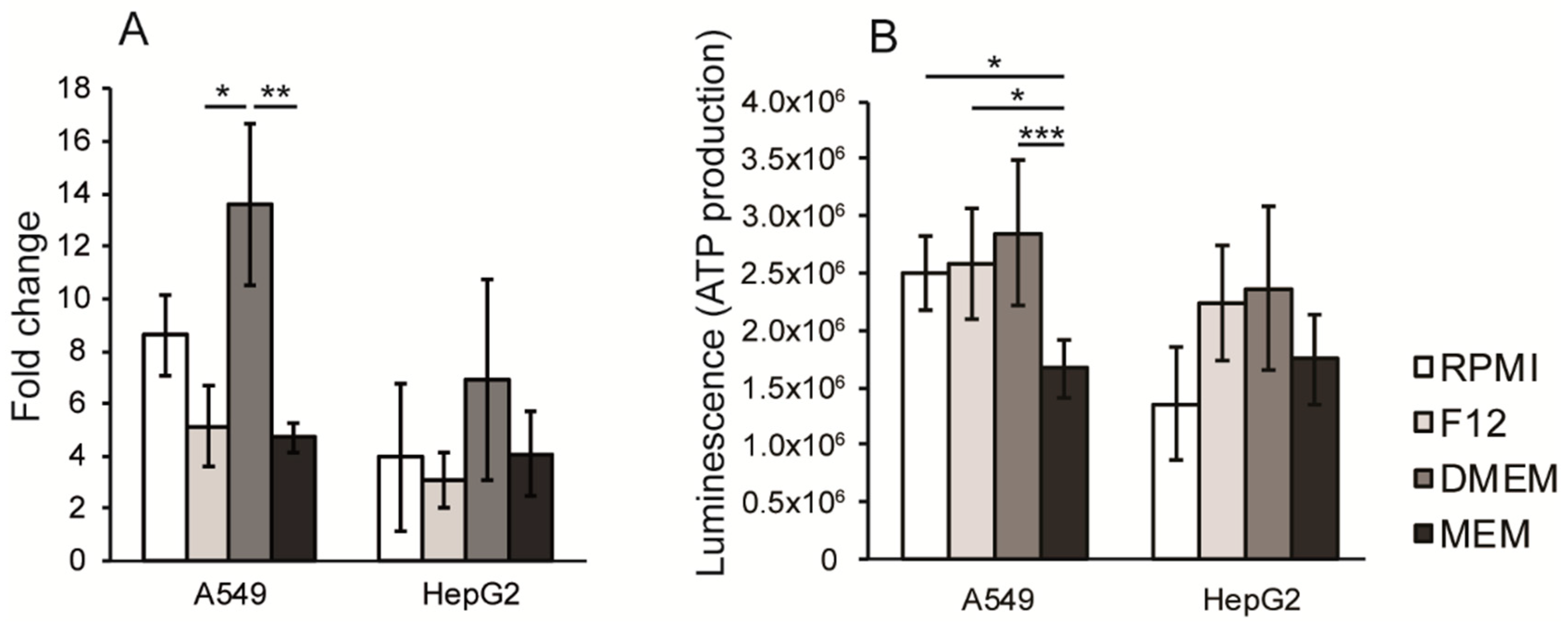
3.3. Proliferation Rate and ATP Production are Dependent on Cell Culture Media Composition
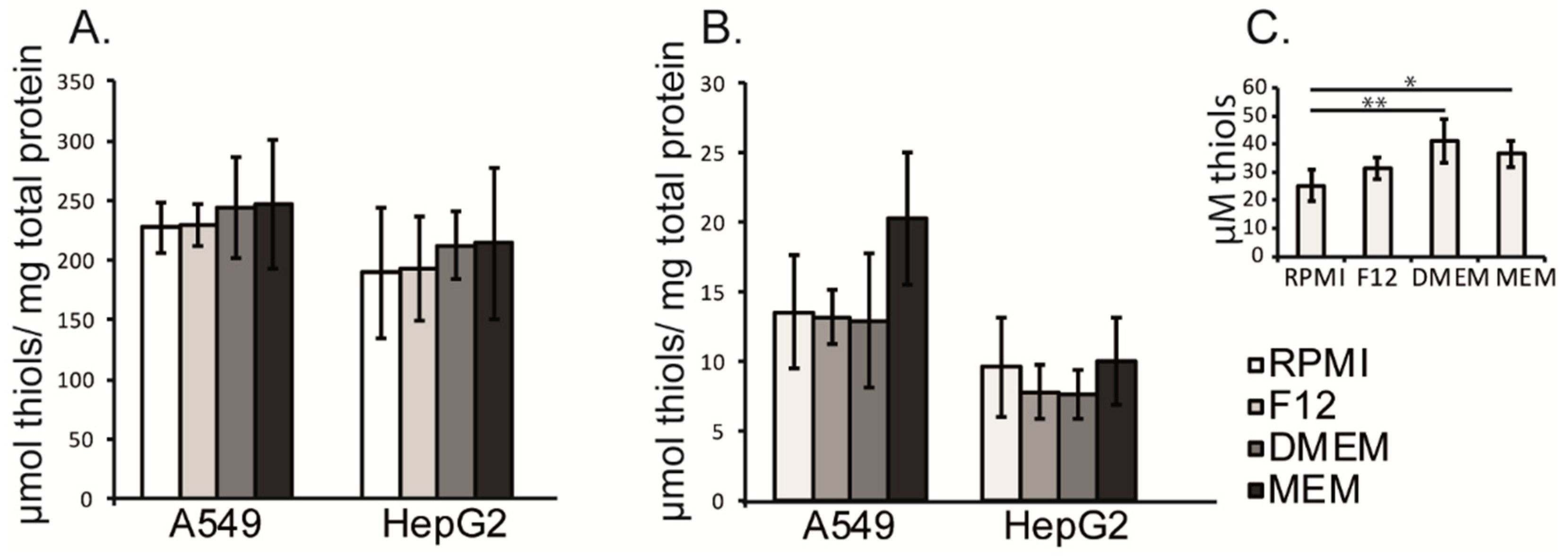
3.4. Intracellular and Extracellular Thiol Content in A549 and HepG2 Cells Were Not Affected by the Different Cell Culture Media
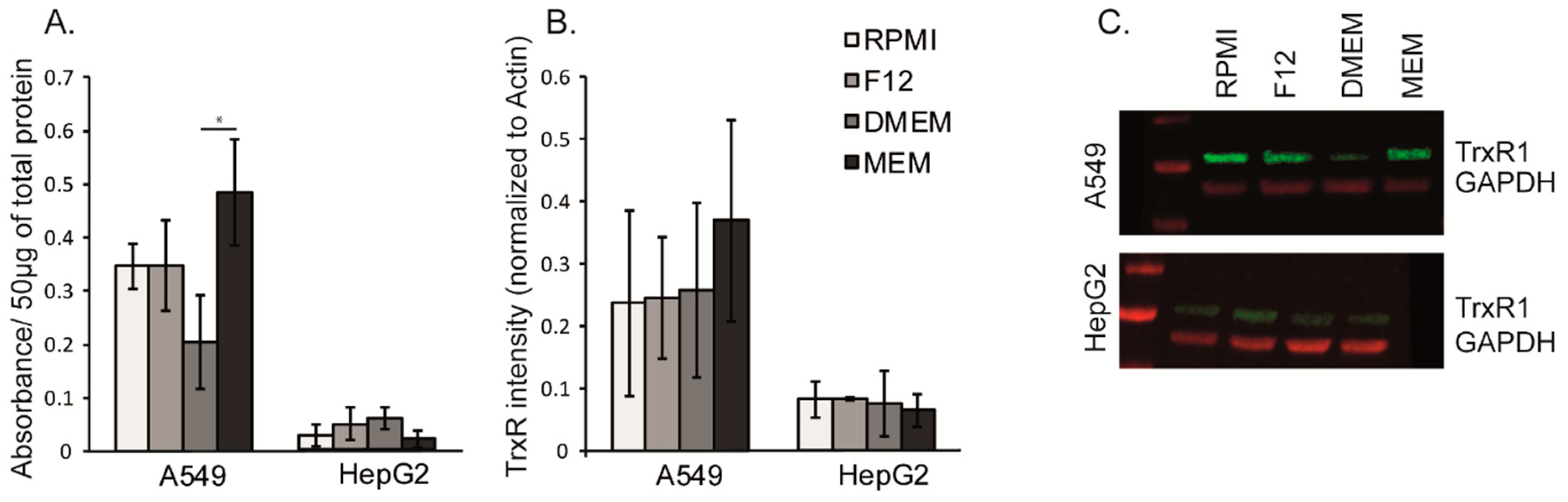
3.5. The Cell Growth Media MEM Effects the Activity and Relative Expression of Thioredoxin Reductase
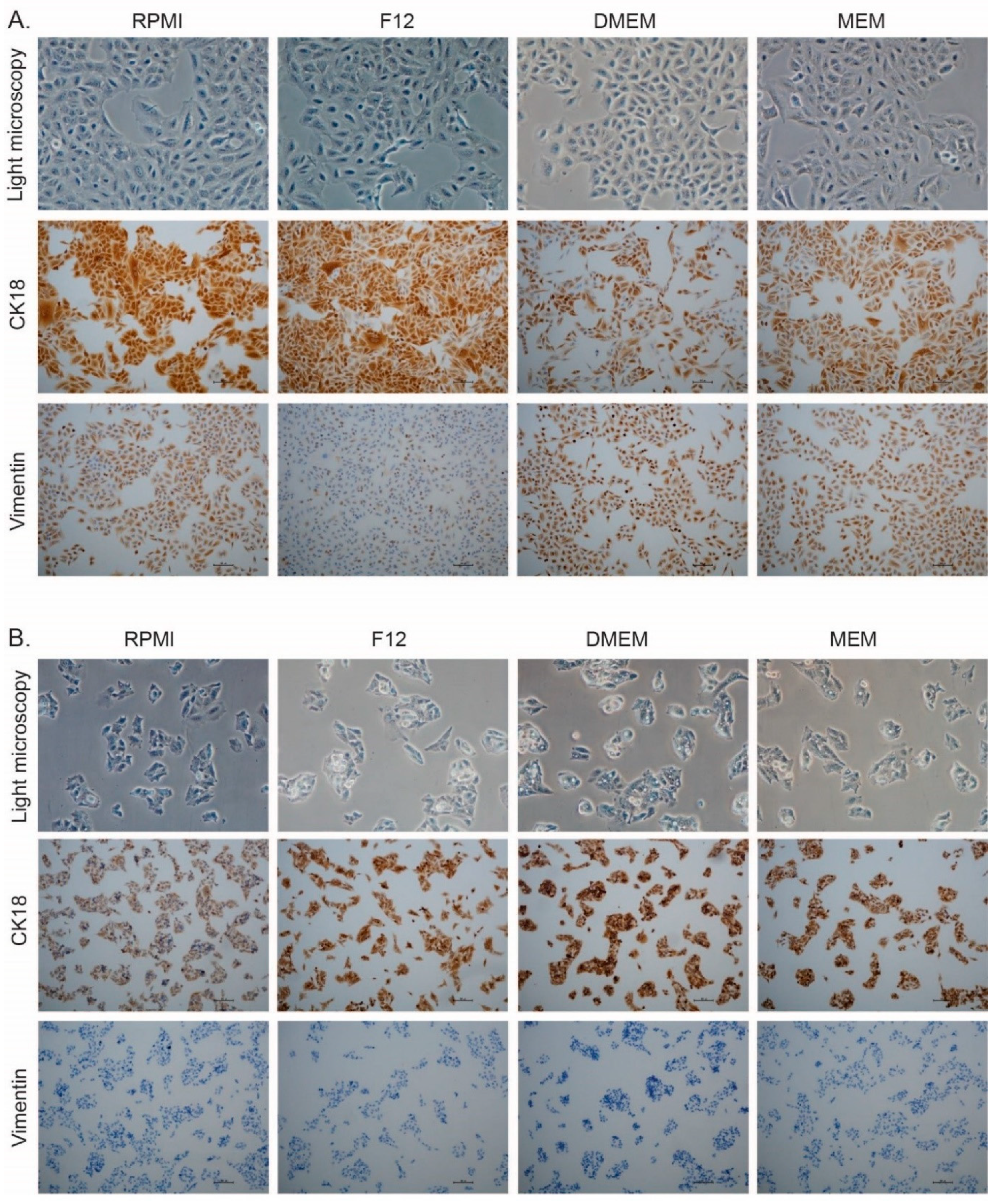
3.6. Selection of Cell Culture Media Changes the Expression of Markers for Epithelial and Mesenchymal Phenotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Misra, S.; Boylan, M.; Selvam, A.; Spallholz, J.E.; Bjornstedt, M. Redox-Active Selenium Compounds—From Toxicity and Cell Death to Cancer Treatment. Nutrients 2015, 7, 3536–3556. [Google Scholar] [CrossRef] [Green Version]
- Brodin, O.; Eksborg, S.; Wallenberg, M.; Asker-Hagelberg, C.; Larsen, E.H.; Mohlkert, D.; Lenneby-Helleday, C.; Jacobsson, H.; Linder, S.; Misra, S.; et al. Pharmacokinetics and Toxicity of Sodium Selenite in the Treatment of Patients with Carcinoma in a Phase I Clinical Trial: The SECAR Study. Nutrients 2015, 7, 4978–4994. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-H.; Kim, J.-H.; Chi, G.Y.; Kim, G.-Y.; Chang, Y.-C.; Moon, S.-K.; Nam, S.-W.; Kim, W.-J.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis and autophagy by sodium selenite in A549 human lung carcinoma cells through generation of reactive oxygen species. Toxicol. Lett. 2012, 212, 252–261. [Google Scholar] [CrossRef]
- Xu, W.; Ma, W.-W.; Zeng, H.-H. Synergistic effect of ethaselen and selenite treatment against A549 human non-small cell lung cancer cells. Asian Pac. J. Cancer Prev. 2014, 15, 7129–7135. [Google Scholar] [CrossRef]
- Weekley, C.M.; Aitken, J.B.; Vogt, S.; Finney, L.A.; Paterson, D.J.; de Jonge, M.D.; Howard, D.L.; Witting, P.K.; Musgrave, I.F.; Harris, H.H. Metabolism of selenite in human lung cancer cells: X-ray absorption and fluorescence studies. J. Am. Chem. Soc. 2011, 133, 18272–18279. [Google Scholar] [CrossRef]
- Branco, V.; Godinho-Santos, A.; Goncalves, J.; Lu, J.; Holmgren, A.; Carvalho, C. Mitochondrial thioredoxin reductase inhibition, selenium status, and Nrf-2 activation are determinant factors modulating the toxicity of mercury compounds. Free Radic. Biol. Med. 2014, 73, 95–105. [Google Scholar] [CrossRef]
- Zou, Y.; Niu, P.; Yang, J.; Yuan, J.; Wu, T.; Chen, X. The JNK signaling pathway is involved in sodium-selenite-induced apoptosis mediated by reactive oxygen in HepG2 cells. Cancer Biol. Ther. 2008, 7, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.M.; Yang, C.F.; Ong, C.N. Sodium selenite-induced oxidative stress and apoptosis in human hepatoma HepG2 cells. Int. J. Cancer 1999, 81, 820–828. [Google Scholar] [CrossRef] [Green Version]
- Chatzakos, V.; Rundlof, A.K.; Ahmed, D.; de Verdier, P.J.; Flygare, J. Inhibition of sphingosine kinase 1 enhances cytotoxicity, ceramide levels and ROS formation in liver cancer cells treated with selenite. Biochem. Pharmacol. 2012, 84, 712–721. [Google Scholar] [CrossRef]
- Kim, T.; Jung, U.; Cho, D.Y.; Chung, A.S. Se-methylselenocysteine induces apoptosis through caspase activation in HL-60 cells. Carcinogenesis 2001, 22, 559–565. [Google Scholar] [CrossRef]
- Li, J.; Zuo, L.; Shen, T.; Xu, C.-M.; Zhang, Z.-N. Induction of apoptosis by sodium selenite in human acute promyelocytic leukemia NB4 cells: involvement of oxidative stress and mitochondria. J. Trace Elem. Med. Biol. 2003, 17, 19–26. [Google Scholar] [CrossRef]
- Schroterova, L.; Kralova, V.; Voracova, A.; Haskova, P.; Rudolf, E.; Cervinka, M. Antiproliferative effects of selenium compounds in colon cancer cells: Comparison of different cytotoxicity assays. Toxicol. In Vitro 2009, 23, 1406–1411. [Google Scholar] [CrossRef]
- Suzuki, M.; Endo, M.; Shinohara, F.; Echigo, S.; Rikiishi, H. Differential apoptotic response of human cancer cells to organoselenium compounds. Cancer Chemother. Pharmacol. 2010, 66, 475–484. [Google Scholar] [CrossRef]
- Okuno, T.; Honda, E.; Arakawa, T.; Ogino, H.; Ueno, H. Glutathione-dependent cell cycle G1 arrest and apoptosis induction in human lung cancer A549 cells caused by methylseleninic acid: comparison with sodium selenite. Biol. Pharm. Bull. 2014, 37, 1831–1837. [Google Scholar] [CrossRef]
- Leclere, L.; Fransolet, M.; Cote, F.; Cambier, P.; Arnould, T.; Van Cutsem, P.; Michiels, C. Heat-modified citrus pectin induces apoptosis-like cell death and autophagy in HepG2 and A549 cancer cells. PLoS ONE 2015, 10, e0115831. [Google Scholar] [CrossRef]
- Speit, G.; Bonzheim, I. Genotoxic and protective effects of hyperbaric oxygen in A549 lung cells. Mutagenesis 2003, 18, 545–548. [Google Scholar] [CrossRef] [Green Version]
- Kipp, A.P.; Frombach, J.; Deubel, S.; Brigelius-Flohe, R. Selenoprotein W as biomarker for the efficacy of selenium compounds to act as source for selenoprotein biosynthesis. Methods Enzymol. 2013, 527, 87–112. [Google Scholar] [PubMed]
- Etebari, M.; Jafarian-Dehkordi, A.; Lame, V. Evaluation of protective effect of amifostine on dacarbazine induced genotoxicity. Res. Pharm. Sci. 2015, 10, 68–74. [Google Scholar]
- Cervello, M.; Notarbartolo, M.; Landino, M.; Cusimano, A.; Virruso, L.; Montalto, G.; D’Alessandro, N. Downregulation of wild-type beta-catenin expression by interleukin 6 in human hepatocarcinoma HepG2 cells: A possible role in the growth-regulatory effects of the cytokine? Eur. J. Cancer 2001, 37, 512–519. [Google Scholar] [CrossRef]
- Liu, H.; Chen, W.; Liang, C.; Chen, B.W.; Zhi, X.; Zhang, S.; Zheng, X.; Bai, X.; Liang, T. WP1130 increases doxorubicin sensitivity in hepatocellular carcinoma cells through usp9x-dependent p53 degradation. Cancer Lett. 2015, 361, 218–225. [Google Scholar] [CrossRef]
- Berger, E.; Vega, N.; Weiss-Gayet, M.; Geloen, A. Gene Network Analysis of Glucose Linked Signaling Pathways and Their Role in Human Hepatocellular Carcinoma Cell Growth and Survival in HuH7 and HepG2 Cell Lines. Biomed. Res. Int. 2015, 2015, 821761. [Google Scholar] [CrossRef]
- Tang, S.Q.; Lee, Y.Y.I.; Packiaraj, D.S.; Ho, H.K.; Chai, C.L.L. Systematic Evaluation of the Metabolism and Toxicity of Thiazolidinone and Imidazolidinone Heterocycles. Chem. Res. Toxicol. 2015, 28, 2019–2033. [Google Scholar] [CrossRef]
- Cuello, S.; Ramos, S.; Mateos, R.; Martin, M.A.; Madrid, Y.; Camara, C.; Bravo, L.; Goya, L. Selenium methylselenocysteine protects human hepatoma HepG2 cells against oxidative stress induced by tert-butyl hydroperoxide. Anal. Bioanal. Chem. 2007, 389, 2167–2178. [Google Scholar] [CrossRef]
- Scientific, T.F. Recommended Media Types for Common Cells. Available online: https://www.thermofisher.com/se/en/home/references/gibco-cell-culture-basics/cell-culture-protocols/maintaining-cultured-cells/recommended-media-types-for-common-cells.html (accessed on 5 April 2015).
- Arora, M. Cell Culture Media: A Review. Mater. Methods. 2013, 3, 24. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, S.J.; Langley, R.R.; Fidler, I.J. Modulation of the cancer cell transcriptome by culture media formulations and cell density. Int. J. Oncol. 2015, 46, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Olm, E.; Fernandes, A.P.; Hebert, C.; Rundlof, A.-K.; Larsen, E.H.; Danielsson, O.; Bjornstedt, M. Extracellular thiol-assisted selenium uptake dependent on the xc− cystine transporter explains the cancer-specific cytotoxicity of selenite. Proc. Natl. Acad. Sci. USA 2009, 106, 11400–11405. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Holmgren, A.; Bjornstedt, M. Thioredoxin and thioredoxin reductase. Methods Enzymol. 1995, 252, 199–208. [Google Scholar]
- Lai, A.G.; Forde, D.; Chang, W.H.; Yuan, F.; Zhuang, X.; Orbegozo Rubio, C.; Song, C.X.; McKeating, J.A. Glucose and glutamine availability regulate HepG2 transcriptional responses to low oxygen. Wellcome Open Res. 2018, 3, 126. [Google Scholar] [CrossRef]
- Hoang, N.A.; Richter, F.; Schubert, M.; Lorkowski, S.; Klotz, L.O.; Steinbrenner, H. Differential capability of metabolic substrates to promote hepatocellular lipid accumulation. Eur. J. Nutr. 2018. [Google Scholar] [CrossRef]
- Han, J.; Zhang, L.; Guo, H.; Wysham, W.Z.; Roque, D.R.; Willson, A.K.; Sheng, X.; Zhou, C.; Bae-Jump, V.L. Glucose promotes cell proliferation, glucose uptake and invasion in endometrial cancer cells via AMPK/mTOR/S6 and MAPK signaling. Gynecol. Oncol. 2015, 138, 668–675. [Google Scholar] [CrossRef] [Green Version]
- Vincent, B.R.; Mousset, S.; Jacquemin-Sablon, A. Cysteine control over glutathione homeostasis in Chinese hamster fibroblasts overexpressing a gamma-glutamylcysteine synthetase activity. Eur. J. Biochem. 1999, 262, 873–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Shi, Y.; Hou, Y.; Ren, Y.; Du, C.; Zhang, L.; Li, Y.; Duan, H. Knockdown of thioredoxin-interacting protein ameliorates high glucose-induced epithelial to mesenchymal transition in renal tubular epithelial cells. Cell. Signal. 2013, 25, 2788–2796. [Google Scholar] [CrossRef]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef]
- Kumar, S.; Bjornstedt, M.; Holmgren, A. Selenite is a substrate for calf thymus thioredoxin reductase and thioredoxin and elicits a large non-stoichiometric oxidation of NADPH in the presence of oxygen. Eur. J. Biochem. 1992, 207, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Bjornstedt, M.; Kumar, S.; Holmgren, A. Selenodiglutathione is a highly efficient oxidant of reduced thioredoxin and a substrate for mammalian thioredoxin reductase. J. Biol. Chem. 1992, 267, 8030–8034. [Google Scholar]
- Bjornstedt, M.; Hamberg, M.; Kumar, S.; Xue, J.; Holmgren, A. Human thioredoxin reductase directly reduces lipid hydroperoxides by NADPH and selenocystine strongly stimulates the reaction via catalytically generated selenols. J. Biol. Chem. 1995, 270, 11761–11764. [Google Scholar] [CrossRef]
- Xia, L.; Nordman, T.; Olsson, J.M.; Damdimopoulos, A.; Bjorkhem-Bergman, L.; Nalvarte, I.; Eriksson, L.C.; Arner, E.S.J.; Spyrou, G.; Bjornstedt, M. The mammalian cytosolic selenoenzyme thioredoxin reductase reduces ubiquinone: A novel mechanism for defense against oxidative stress. J. Biol. Chem. 2003, 278, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Arner, E.S.; Nordberg, J.; Holmgren, A. Efficient reduction of lipoamide and lipoic acid by mammalian thioredoxin reductase. Biochem. Biophys. Res. Commun. 1996, 225, 268–274. [Google Scholar] [CrossRef]
- May, J.M.; Mendiratta, S.; Hill, K.E.; Burk, R.F. Reduction of dehydroascorbate to ascorbate by the selenoenzyme thioredoxin reductase. J. Biol. Chem. 1997, 272, 22607–22610. [Google Scholar] [CrossRef]
- Selenius, M.; Rundlof, A.-K.; Olm, E.; Fernandes, A.P.; Bjornstedt, M. Selenium and the selenoprotein thioredoxin reductase in the prevention, treatment and diagnostics of cancer. Antioxid. Redox Signal. 2010, 12, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Wallenberg, M.; Misra, S.; Bjornstedt, M. Selenium cytotoxicity in cancer. Basic Clin. Pharmacol. Toxicol. 2014, 114, 377–386. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selenite | Medium | FBS | Antibiotics | IC50 | Evaluated by | Ref. |
| Cell Line | ||||||
| A549 | RPMI 1640 | 10% | Yes | ~6 µM | MTT | [3] |
| A549 | DMEM | 10% | No | 8.2 µM | Sulforhodamine | [4] |
| A549 | DMEM+ non ess. AA | 2% | Yes | 5 µM = 85% viability | MTT | [5] |
| HepG2 | 1:1 DMEM:F12 | 10% | Yes | 25.7 µM | MTT | [6] |
| HepG2 | DMEM | 10% | Yes | ~10 µM | MTT (48 h) | [7] |
| HepG2 | MEM | 10% | Yes | ~7.5 µM | Tunnel-assay | [8] |
| Huh7 | DMEM | 10% | Yes | ~20 µM | XTT | [9] |
| HL-60 | RPMI 1640 | 10% | Yes | 20 µM | DNA fragmentation | [10] |
| NB 4 | RPMI 1640 | 10% | Yes | 20 µM | Trypan blue | [11] |
| MSC | Medium | FBS | Antibiotics | IC50 | Evaluated by | Ref. |
| Cell Line | ||||||
| HT 29/ SW480/ SW620 | DMEM | 10% | Yes | ~64/~32/ ~90µM | MTT (48 h) | [12] |
| HL-60 | RPMI 1640 | 10% | Yes | 50 µM | DNA fragmentation | [10] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arodin Selenius, L.; Wallenberg Lundgren, M.; Jawad, R.; Danielsson, O.; Björnstedt, M. The Cell Culture Medium Affects Growth, Phenotype Expression and the Response to Selenium Cytotoxicity in A549 and HepG2 Cells. Antioxidants 2019, 8, 130. https://doi.org/10.3390/antiox8050130
Arodin Selenius L, Wallenberg Lundgren M, Jawad R, Danielsson O, Björnstedt M. The Cell Culture Medium Affects Growth, Phenotype Expression and the Response to Selenium Cytotoxicity in A549 and HepG2 Cells. Antioxidants. 2019; 8(5):130. https://doi.org/10.3390/antiox8050130
Chicago/Turabian StyleArodin Selenius, Lisa, Marita Wallenberg Lundgren, Rim Jawad, Olof Danielsson, and Mikael Björnstedt. 2019. "The Cell Culture Medium Affects Growth, Phenotype Expression and the Response to Selenium Cytotoxicity in A549 and HepG2 Cells" Antioxidants 8, no. 5: 130. https://doi.org/10.3390/antiox8050130
APA StyleArodin Selenius, L., Wallenberg Lundgren, M., Jawad, R., Danielsson, O., & Björnstedt, M. (2019). The Cell Culture Medium Affects Growth, Phenotype Expression and the Response to Selenium Cytotoxicity in A549 and HepG2 Cells. Antioxidants, 8(5), 130. https://doi.org/10.3390/antiox8050130