Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Retinal Pigment Epithelial Cell Culture Maintenance
2.2. Methylthiazolyldiphenyl-Tetrazolium Bromide Assay to Test Cell Viability
2.3. RNA Extraction and Whole Transcriptome Analysis
2.4. FASTQ Data Quality Control and Read Mapping
2.5. Gene Expression Quantification, Differential Gene Expression (DGE) and Statistical Analysis
2.6. Functional Gene Annotation, Enrichment Analysis and Gene Prioritization
2.7. Quantitative RT-PCR Data Validation
2.8. Statistical Analysis
3. Results
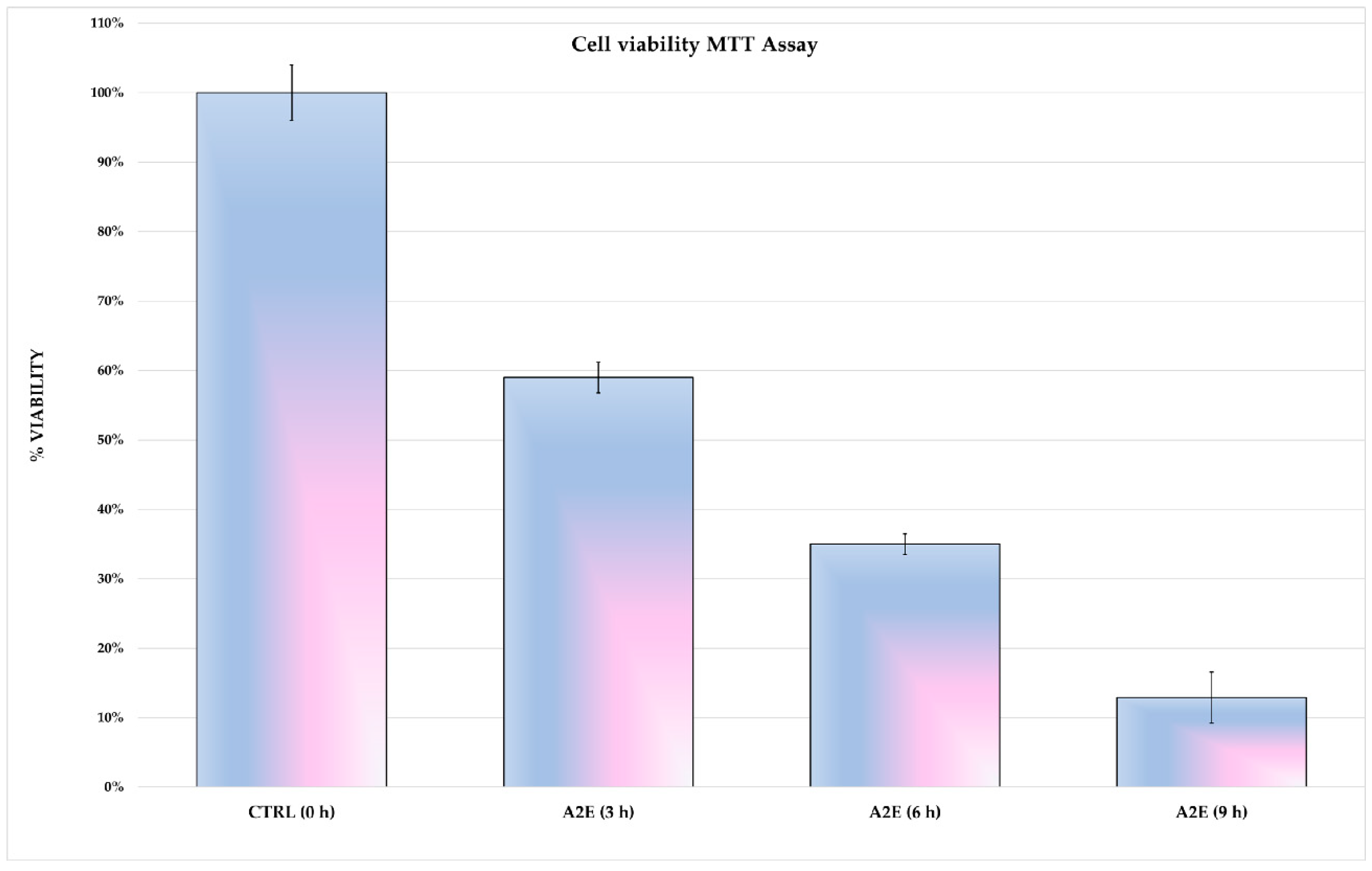
3.1. A2E Affects H-RPE Cell Viability
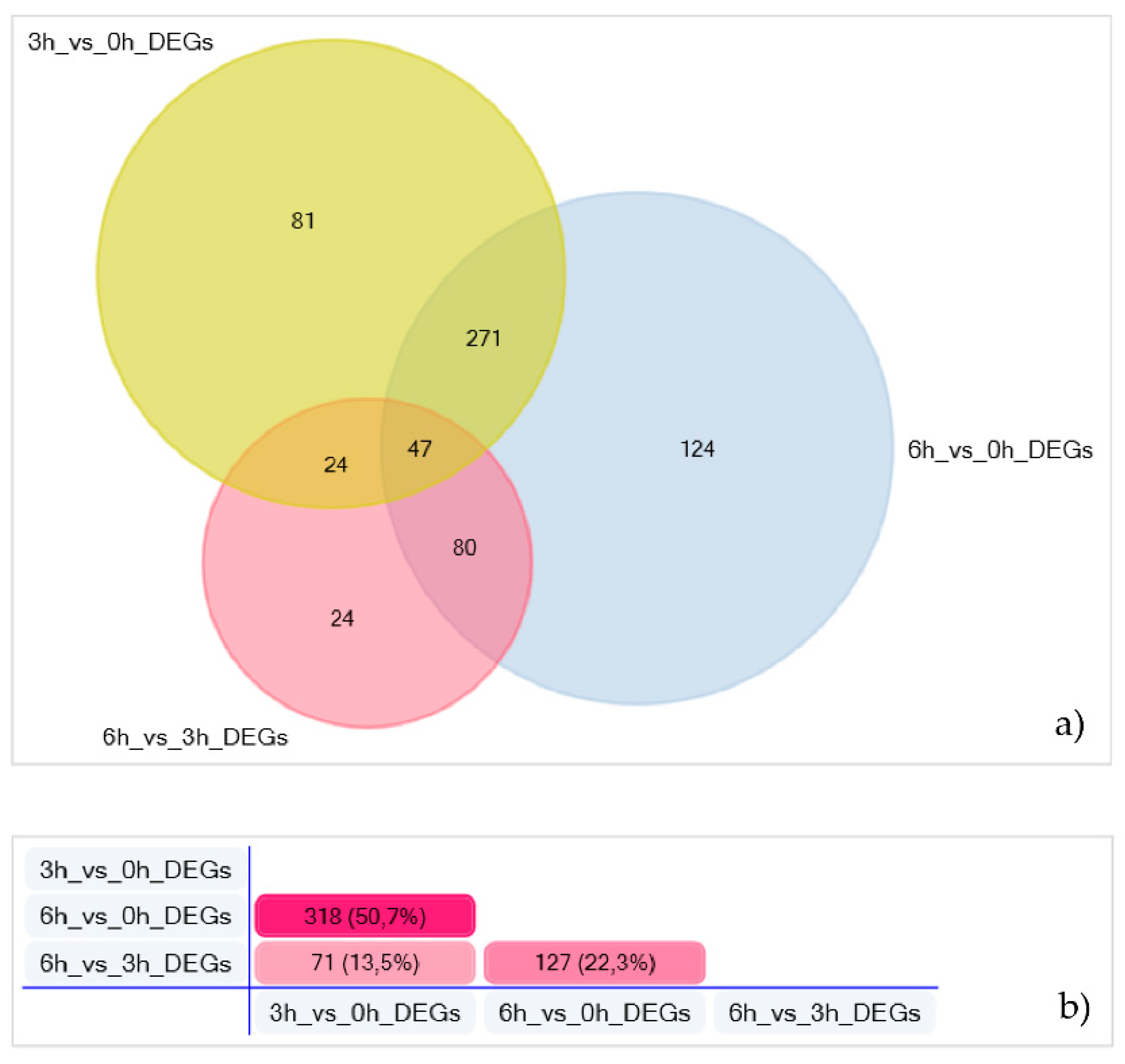
3.2. Transcriptome Analysis and Differential Gene Expression
3.3. Functional Pathway Analysis
3.4. A2E Treatment CAUSES Loss of Retinal Blood Barrier Properties
3.5. A2E Treatment Impairs Expression of AHR and ROM1
3.6. Quantitative RT-PCR Data Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Deposition
References
- Fuhrmann, S.; Zou, C.; Levine, E.M. Retinal pigment epithelium development, plasticity, and tissue homeostasis. Exp. Eye Res. 2014, 123, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Muniz, A.; Betts, B.S.; Trevino, A.R.; Buddavarapu, K.; Roman, R.; Ma, J.X.; Tsin, A.T. Evidence for two retinoid cycles in the cone-dominated chicken eye. Biochemistry 2009, 48, 6854–6863. [Google Scholar] [CrossRef] [Green Version]
- Fields, M.A.; Del Priore, L.V.; Adelman, R.A.; Rizzolo, L.J. Interactions of the choroid, Bruch’s membrane, retinal pigment epithelium, and neurosensory retina collaborate to form the outer blood-retinal-barrier. Prog. Retin. Eye Res. 2020, 76, 100803. [Google Scholar] [CrossRef] [PubMed]
- Hellinen, L.; Sato, K.; Reinisalo, M.; Kidron, H.; Rilla, K.; Tachikawa, M.; Uchida, Y.; Terasaki, T.; Urtti, A. Quantitative Protein Expression in the Human Retinal Pigment Epithelium: Comparison Between Apical and Basolateral Plasma Membranes with Emphasis on Transporters. Investig. Ophthalmol. Vis. Sci. 2019, 60, 5022–5034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichhart, N.; Strauss, O. Ion channels and transporters of the retinal pigment epithelium. Exp. Eye Res. 2014, 126, 27–37. [Google Scholar] [CrossRef]
- Zhang, H.; Wong, C.L.; Shan, S.W.; Li, K.K.; Cheng, A.K.; Lee, K.L.; Ge, J.; To, C.H.; Do, C.W. Characterisation of Cl⁻ transporter and channels in experimentally induced myopic chick eyes. Clin. Exp. Optom. 2011, 94, 528–535. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Leung, K.W.; Ren, Y.; Pei, J.; Ge, J.; Tombran-Tink, J. PEDF improves mitochondrial function in RPE cells during oxidative stress. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6742–6755. [Google Scholar] [CrossRef]
- Hernández, C.; Simó-Servat, O.; Simó, R. Somatostatin and diabetic retinopathy: Current concepts and new therapeutic perspectives. Endocrine 2014, 46, 209–214. [Google Scholar] [CrossRef]
- Samson, F.P.; He, W.; Sripathi, S.R.; Patrick, A.T.; Madu, J.; Chung, H.; Frost, M.C.; Jee, D.; Gutsaeva, D.R.; Jahng, W.J. Dual Switch Mechanism of Erythropoietin as an Antiapoptotic and Pro-Angiogenic Determinant in the Retina. ACS Omega 2020, 5, 21113–21126. [Google Scholar] [CrossRef]
- Farnoodian, M.; Kinter, J.B.; Yadranji Aghdam, S.; Zaitoun, I.; Sorenson, C.M.; Sheibani, N. Expression of pigment epithelium-derived factor and thrombospondin-1 regulate proliferation and migration of retinal pigment epithelial cells. Physiol. Rep. 2015, 3, e12266. [Google Scholar] [CrossRef]
- Zadeh, J.K.; Zhutdieva, M.B.; Laspas, P.; Yuksel, C.; Musayeva, A.; Pfeiffer, N.; Brochhausen, C.; Oelze, M.; Daiber, A.; Xia, N.; et al. Apolipoprotein E Deficiency Causes Endothelial Dysfunction in the Mouse Retina. Oxid. Med. Cell Longev. 2019, 2019, 5181429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Kozak, Y.; Cotinet, A.; Goureau, O.; Hicks, D.; Thillaye-Goldenberg, B. Tumor necrosis factor and nitric oxide production by resident retinal glial cells from rats presenting hereditary retinal degeneration. Ocul. Immunol. Inflamm. 1997, 5, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Parete, G.; Flati, V.; Maccarone, R. Up-regulation of pro-angiogenic pathways and induction of neovascularization by an acute retinal light damage. Sci. Rep. 2020, 10, 6376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Cai, X.; Wu, Y.; Liu, Y.; Deng, L.; Chen, H. Insights from Genetic Model Systems of Retinal Degeneration: Role of Epsins in Retinal Angiogenesis and VEGFR2 Signaling. J. Nat. Sci. 2017, 3, e281. [Google Scholar]
- Shen, C.; Li, Y.; Wang, Q.; Chen, Y.N.; Li, W.; Wei, W.B. Choroidal vascular changes in retinitis pigmentosa patients detected by optical coherence tomography angiography. BMC Ophthalmol. 2020, 20, 384. [Google Scholar] [CrossRef]
- Garafalo, A.V.; Cideciyan, A.V.; Héon, E.; Sheplock, R.; Pearson, A.; WeiYang Yu, C.; Sumaroka, A.; Aguirre, G.D.; Jacobson, S.G. Progress in treating inherited retinal diseases: Early subretinal gene therapy clinical trials and candidates for future initiatives. Prog. Retin. Eye Res. 2020, 77, 100827. [Google Scholar] [CrossRef]
- Strobbe, E.; Cellini, M.; Fresina, M.; Campos, E.C. ET-1 Plasma Levels, Aqueous Flare, and Choroidal Thickness in Patients with Retinitis Pigmentosa. J. Ophthalmol. 2015, 2015, 292615. [Google Scholar] [CrossRef] [Green Version]
- Narayan, S.; Prasanna, G.; Tchedre, K.; Krishnamoorthy, R.; Yorio, T. Thrombin-induced endothelin-1 synthesis and secretion in retinal pigment epithelial cells is rho kinase dependent. J. Ocul. Pharmacol. Ther. 2010, 26, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.Y.; Possin, D.E.; Milam, A.H. Histopathology of bone spicule pigmentation in retinitis pigmentosa. Ophthalmology 1995, 102, 805–816. [Google Scholar] [CrossRef]
- Farnoodian, M.; Halbach, C.; Slinger, C.; Pattnaik, B.R.; Sorenson, C.M.; Sheibani, N. High glucose promotes the migration of retinal pigment epithelial cells through increased oxidative stress and PEDF expression. Am. J. Physiol. Cell Physiol. 2016, 311, C418–C436. [Google Scholar] [CrossRef] [Green Version]
- Marano, F.; Deutman, A.F.; Leys, A.; Aandekerk, A.L. Hereditary retinal dystrophies and choroidal neovascularization. Graefes Arch. Clin. Exp. Ophthalmol. 2000, 238, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Sood, S.; Narang, S. Successful treatment of choroidal neovascular membrane in retinitis pigmentosa with intravitreal bevacizumab. Int. Ophthalmol. 2010, 30, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Campochiaro, P.A.; Mir, T.A. The mechanism of cone cell death in Retinitis Pigmentosa. Prog. Retin. Eye Res. 2018, 62, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced multi-sample quality control for high-throughput sequencing data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Ruotti, V.; Stewart, R.M.; Thomson, J.A.; Dewey, C.N. RNA-Seq gene expression estimation with read mapping uncertainty. Bioinformatics 2010, 26, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving Coverage, Classification and Access to Protein Sequence Annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A Pathology Atlas of the Human Cancer Transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orchard, S.; Ammari, M.; Aranda, B.; Breuza, L.; Briganti, L.; Broackes-Carter, F.; Campbell, N.H.; Chavali, G.; Chen, C.; del-Toro, N.; et al. The MIntAct project—IntAct as a Common Curation Platform for 11 Molecular Interaction Databases. Nucleic Acids Res. 2014, 42, D358–D563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aken, B.L.; Ayling, S.; Barrell, D.; Clarke, L.; Curwen, V.; Fairley, S.; Banet, J.F.; Billis, K.; García Girón, C.; Hourlier, T.; et al. The Ensembl Gene Annotation System. Database 2016, 2016, baw093. [Google Scholar] [CrossRef] [PubMed]
- Wain, H.M.; Bruford, E.A.; Lovering, R.C.; Lush, M.J.; Wright, M.W.; Povey, S. Guidelines for Human Gene Nomenclature. Genomics 2002, 79, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.S.; Askenase, P.; Batagov, A.O.; Benito-Martin, A.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of extracellular vesicles data. J. Extracell. Vesicles 2017, 6, 1321455. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef]
- Choudhary, M.; Kazmin, D.; Hu, P.; Thomas, R.S.; McDonnell, D.P.; Malek, G. Aryl hydrocarbon receptor knock-out exacerbates choroidal neovascularization via multiple pathogenic pathways. J. Pathol. 2015, 235, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, M.; Binder, A.; Menzel, U.; Zeiter, S.; Alini, M.; Verrier, S. CD34/CD133 enriched bone marrow progenitor cells promote neovascularization of tissue engineered constructs in vivo. Stem Cell Res. 2014, 13, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Novitskiy, S.V.; Zaynagetdinov, R.; Vasiukov, G.; Gutor, S.; Han, W.; Serezani, A.; Matafonov, A.; Gleaves, L.A.; Sherrill, T.P.; Polosukhin, V.V.; et al. Gas6/MerTK signaling is negatively regulated by NF-κB and supports lung carcinogenesis. Oncotarget 2019, 10, 7031–7042. [Google Scholar] [CrossRef]
- Liu, S.; Biesemeier, A.K.; Tschulakow, A.V.; Thakkar, H.V.; Julien-Schraermeyer, S.; Schraermeyer, U. A new rat model of treatment-naive quiescent choroidal neovascularization induced by human VEGF165 overexpression. Biol. Open. 2020, 9, bio048736. [Google Scholar] [CrossRef] [Green Version]
- Massengill, M.T.; Young, B.; Patel, D.; Jafri, F.; Sabogal, E.; Ash, N.; Li, H.; Ildefonso, C.J.; Lewin, A.S. Clinically Relevant Outcome Measures for the I307N Rhodopsin Mouse: A Model of Inducible Autosomal Dominant Retinitis Pigmentosa. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5417–5430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, L.A.; Morrison, M.A.; Ahn, J.; Woo, S.J.; Sato, H.; Robinson, R.; Morgan, D.J.; Zacharaki, F.; Simeonova, M.; Uehara, H.; et al. FLT1 genetic variation predisposes to neovascular AMD in ethnically diverse populations and alters systemic FLT1 expression. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3543–3554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, D.; Kumanogoh, A. The role of Sema4A in angiogenesis, immune responses, carcinogenesis, and retinal systems. Cell Adhes. Migr. 2016, 10, 692–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagani, F.; Trivedi, A.; Khatri, D.; Zizioli, D.; Garrafa, E.; Mitola, S.; Finazzi, D. Silencing of pantothenate kinase 2 reduces endothelial cell angiogenesis. Mol. Med. Rep. 2018, 18, 4739–4746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, P.; Becerra, S.P. Role of the PNPLA2 Gene in the Regulation of Oxidative Stress Damage of RPE. Adv. Exp. Med. Biol. 2019, 1185, 377–382. [Google Scholar]
- Marie, M.; Bigot, K.; Angebault, C.; Barrau, C.; Gondouin, P.; Pagan, D.; Fouquet, S.; Villette, T.; Sahel, J.A.; Lenaers, G.; et al. Light action spectrum on oxidative stress and mitochondrial damage in A2E-loaded retinal pigment epithelium cells. Cell Death Dis. 2018, 9, 287. [Google Scholar] [CrossRef]
- Nakagami, Y.; Hatano, E.; Inoue, T.; Yoshida, K.; Kondo, M.; Terasaki, H. Cytoprotective Effects of a Novel Nrf2 Activator, RS9, in Rhodopsin Pro347Leu Rabbits. Curr. Eye Res. 2016, 41, 1123–1126. [Google Scholar] [CrossRef]
- Donato, L.; D’Angelo, R.; Alibrandi, S.; Rinaldi, C.; Sidoti, A.; Scimone, C. Effects of A2E-Induced Oxidative Stress on Retinal Epithelial Cells: New Insights on Differential Gene Response and Retinal Dystrophies. Antioxidants 2020, 9, 307. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.S.; Hussein, K.; Wang, F.; Wan, M.; Saad, N.; Essa, M.; Kim, I.; Shakoor, A.; Owen, L.A.; DeAngelis, M.M.; et al. Bone Morphogenetic Protein (BMP)4 But Not BMP2 Disrupts the Barrier Integrity of Retinal Pigment Epithelia and Induces Their Migration: A Potential Role in Neovascular Age-Related Macular Degeneration. J. Clin. Med. 2020, 9, 2293. [Google Scholar] [CrossRef]
- Vogt, R.R.; Unda, R.; Yeh, L.C.; Vidro, E.K.; Lee, J.C.; Tsin, A.T. Bone morphogenetic protein-4 enhances vascular endothelial growth factor secretion by human retinal pigment epithelial cells. J. Cell. Biochem. 2006, 98, 1196–1202. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.A.; Kim, H.S.; Vargas, A.; Yu, W.Q.; Eom, Y.S.; Craft, C.M.; Lee, E.J. Inhibition of Matrix Metalloproteinase 9 Enhances Rod Survival in the S334ter-line3 Retinitis Pigmentosa Model. PLoS ONE 2016, 11, e0167102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Vargas, A.; Eom, Y.S.; Li, J.; Yamamoto, K.L.; Craft, C.; Lee, E.J. Tissue inhibitor of metalloproteinases 1 enhances rod survival in the rd1 mouse retina. PLoS ONE 2018, 13, e0197322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Liu, X.; Wang, S.; Miao, J.; Wu, L.; Yang, X.; Wang, Y.; Kang, L.; Li, W.; Cui, C.; et al. PKR promotes choroidal neovascularization via upregulating the PI3K/Akt signaling pathway in VEGF expression. Mol. Vis. 2016, 22, 1361–1374. [Google Scholar] [PubMed]
- Domènech, E.B.; Marfany, G. The Relevance of Oxidative Stress in the Pathogenesis and Therapy of Retinal Dystrophies. Antioxidants 2020, 9, 347. [Google Scholar] [CrossRef] [Green Version]
- Vila, N.; Siblini, A.; Esposito, E.; Bravo-Filho, V.; Zoroquiain, P.; Aldrees, S.; Logan, P.; Arias, L.; Burnier, M.N. Blue-light filtering alters angiogenic signaling in human retinal pigmented epithelial cells culture model. BMC Ophthalmol. 2017, 17, 198. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Sparrow, J.R. Light filtering in a retinal pigment epithelial cell culture model. Optom. Vis. Sci. 2011, 88, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, C.; Lei, W.; Wang, K.; Zheng, J. Roles of Aryl Hydrocarbon Receptor in Endothelial Angiogenic Responses. Biol. Reprod. 2020, 103, 927–937. [Google Scholar] [CrossRef]
- Ziegler, N.; Awwad, K.; Fisslthaler, B.; Reis, M.; Devraj, K.; Corada, M.; Minardi, S.P.; Dejana, E.; Plate, K.H.; Fleming, I.; et al. β-Catenin Is Required for Endothelial Cyp1b1 Regulation Influencing Metabolic Barrier Function. J. Neurosci. 2016, 36, 8921–8935. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, T.; Ikeda, K.; Masuya, H.; Sezutsu, H.; Kaneda, H.; Kobayashi, K.; Miura, I.; Kurihara, Y.; Yokokura, S.; et al. A monogenic dominant mutation in Rom1 generated by N-ethyl-N-nitrosourea mutagenesis causes retinal degeneration in mice. Mol. Vis. 2010, 16, 378–391. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HGNC Gene Name (ID) | Ensembl Transcript ID | 3 h_vs._0 h RNAseq log2 FC | 6 h_vs._0 h RNAseq log2 FC | Primer Pair | Fragment Length (bp) |
|---|---|---|---|---|---|
| OCLN (8104) | ENST00000355237.2 | −0.873423348 | −1.391662424 | ACTTCGCCTGTGGATGACTT GACCTTCCTGCTCTTCCCTT | 101 |
| RICTOR (28611) | ENST00000357387.8 | −0.940830609 | −0.705087984 | GCTCTCTGAAGAACCTCCGA CCTGCAATCTGGCCACATTT | 126 |
| TJP1 (11827) | ENST00000356107.10 | −0.694212076 | −1.059176648 | TCTTCGCAGCTCCAAGAGAA AGGCCTCAGAAATCCAGCTT | 121 |
| FZD4 (4042) | ENST00000531380.2 | 0.37699785 | 0.532274651 | GGTTTGGTGGCCTTGTTCAA ATCACACACGTTGCAGGAAC | 131 |
| KIFC3 (6326) | ENST00000445690.7 | 0.995878391 | 1.422298598 | GACCCTCACCAACGACTACA TTGCTGTTGACCTCCTCGAT | 123 |
| VEGFA (12628) | ENST00000372055.8 | 0.137384927 | 1.016674127 | CTGTCTTGGGTGCATTGGAG TGATGATTCTGCCCTCCTCC | 101 |
| 3 h_vs._0 h | 6 h_vs._0 h | 6 h_vs._3 h | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Pathway | % Enriched Genes | Fold Enrichment | p-Value (Bonferroni-Adjusted) | % Enriched Genes | Fold Enrichment | p-Value (Bonferroni-Adjusted) | % Enriched Genes | Fold Enrichment | p-Value (Bonferroni-Adjusted) |
| Beta1 integrin cell surface interactions | 25.4 | 1.1854 | 0.0146 | 26.9 | 1.2561 | 1.0167 × 10−7 | 34.1 | 1.5901 | 1.441 × 10−7 |
| EGFR-dependent Endothelin signalling events | 24.2 | 1.1825 | 0.03440 | 25.9 | 1.2690 | 4.5939 × 10−8 | 33.6 | 1.645 | 1.3314 × 10−8 |
| Endothelins | Not significant | 26.1 | 1.26 | 1.3783 × 10−7 | 33.6 | 1.6222 | 4.1668 × 10−8 | ||
| HIF-1-alpha transcription factor network | Not significant | Not significant | 5.1 | 4.8831 | 8.0756 × 10−8 | ||||
| IGF1 pathway | 24.2 | 1.1838 | 0.0297 | 25.9 | 1.2670 | 6.044 × 10−8 | 33.6 | 1.6424 | 1.5141 × 10−8 |
| LKB1 signalling events | 24.8 | 1.1964 | 0.0070 | 26.5 | 1.2809 | 4.0980 × 10−9 | 33.6 | 1.6210 | 4.4347 × 10−8 |
| N-cadherin signalling events | Not significant | Not significant | 8.2 | 2.0655 | 0.0252 | ||||
| Nectin adhesion pathway | 24.2 | 1.1801 | 0.0409 | 26.0 | 1.2659 | 6.6551 × 10−8 | 34.1 | 1.6590 | 4.2976 × 10−9 |
| PDGF receptor signalling network | 24.3 | 1.1851 | 0.0257 | 26.06 | 1.2707 | 3.2165 × 10−8 | 33.8 | 1.6507 | 8.0949 × 10−9 |
| Posttranslational regulation of adherens junction stability and disassembly | Not significant | Not significant | 7.8 | 2.1230 | 0.0235 | ||||
| Proteoglycan syndecan-mediated signalling events | 25.3 | 1.1846 | 0.0167 | 27.1 | 1.2724 | 7.3776 × 10−9 | 35.0 | 1.6389 | 5.2725 × 10−9 |
| RAC1 signalling pathway | Not significant | Not significant | 7.3 | 2.2556 | 0.0113 | ||||
| Regulation of CDC42 activity | Not significant | 15.2 | 1.2478 | 0.0083 | 22.7 | 1.8607 | 1.1663 × 10−7 | ||
| RhoA signalling pathway | Not significant | Not significant | 7.3 | 2.2556 | 0.0113 | ||||
| Signalling events mediated by focal adhesion kinase | 24.2 | 1.1834 | 0.0317 | 25.9 | 1.27 | 4.0022 × 10−8 | 33.6 | 1.6462 | 1.2483 × 10−8 |
| Signalling events mediated by Hepatocyte Growth Factor Receptor (c-Met) | 24.3 | 1.1851 | 0.0257 | 26.0 | 1.2679 | 5.0634 × 10−8 | 33.6 | 1.6398 | 1.7211 × 10−8 |
| Signalling events mediated by VEGFR1 and VEGFR2 | 24.7 | 1.1855 | 0.0240 | 26.1 | 1.2705 | 3.0898 × 10−8 | 33.8 | 1.6469 | 9.8334 × 10−9 |
| Stabilization and expansion of the E-cadherin adherens junction | Not significant | Not significant | 9.3 | 2.1399 | 0.0025 | ||||
| Syndecan-1-mediated signalling events | 24.5 | 1.1881 | 0.0178 | 26.2 | 1.2721 | 2.1586 × 10−8 | 33.6 | 1.6310 | 2.6854 × 10−8 |
| VEGF and VEGFR signalling network | 24.4 | 1.1782 | 0.0455 | 26.2 | 1.2655 | 5.8984 × 10−8 | 33.8 | 1.6368 | 1.6437 × 10−8 |
| Locus (HGCN ID) | RP Phenotype | Angiogenesis | Log2 FC 3 h_vs._0 h | Log2 FC 6 h_vs._0 h |
|---|---|---|---|---|
| AHR (348) | recessive | If depleted, enhances retinal angiogenesis [37] | −0.78173956 | −1.52949254 |
| PROM1 (9454) | recessive with macular degeneration | Expressed on EPCS; promotes neovascularization [38] | not significant | |
| MERTK (7027) | recessive | Promotes EC survival by inhibiting apoptosis [39] | not significant | |
| RPE65 (10294) | recessive | Overexpressed in RPE cells surrounding CNV [40] | not determined | |
| RHO (10012) | - dominant; - recessive | If mutated, leads to retinal vessels atrophy following light exposure [41] | not significant | |
| NR2E3 (7974) | - recessive in Portuguese Crypto Jews; - dominant | Regulates retinal neovascularization by targeting FLT1 [42] | not significant | |
| SEMA4A (10729) | dominant | In ECs, suppresses VEGF-mediated EC migration and proliferation by binding Plexin-D1 [43] | not significant | |
| ROM1 (10254) | dominant | GO Biological process: retina vasculature development in camera-type eye (GO:0061298) | 1.256611685 | 1.123121917 |
| PANK2 (15894) | recessive HARP (hypoprebetalipoproteinemia, acanthocytosis, retinitis pigmentosa, and palladial degeneration) | Required for normal development of angiogenetic properties in ECs [44] | not significant | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scimone, C.; Alibrandi, S.; Scalinci, S.Z.; Trovato Battagliola, E.; D’Angelo, R.; Sidoti, A.; Donato, L. Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells. Antioxidants 2020, 9, 1154. https://doi.org/10.3390/antiox9111154
Scimone C, Alibrandi S, Scalinci SZ, Trovato Battagliola E, D’Angelo R, Sidoti A, Donato L. Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells. Antioxidants. 2020; 9(11):1154. https://doi.org/10.3390/antiox9111154
Chicago/Turabian StyleScimone, Concetta, Simona Alibrandi, Sergio Zaccaria Scalinci, Edoardo Trovato Battagliola, Rosalia D’Angelo, Antonina Sidoti, and Luigi Donato. 2020. "Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells" Antioxidants 9, no. 11: 1154. https://doi.org/10.3390/antiox9111154
APA StyleScimone, C., Alibrandi, S., Scalinci, S. Z., Trovato Battagliola, E., D’Angelo, R., Sidoti, A., & Donato, L. (2020). Expression of Pro-Angiogenic Markers Is Enhanced by Blue Light in Human RPE Cells. Antioxidants, 9(11), 1154. https://doi.org/10.3390/antiox9111154