Andrographis paniculata and Its Bioactive Diterpenoids Against Inflammation and Oxidative Stress in Keratinocytes

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparation of Andrographis paniculata Extract
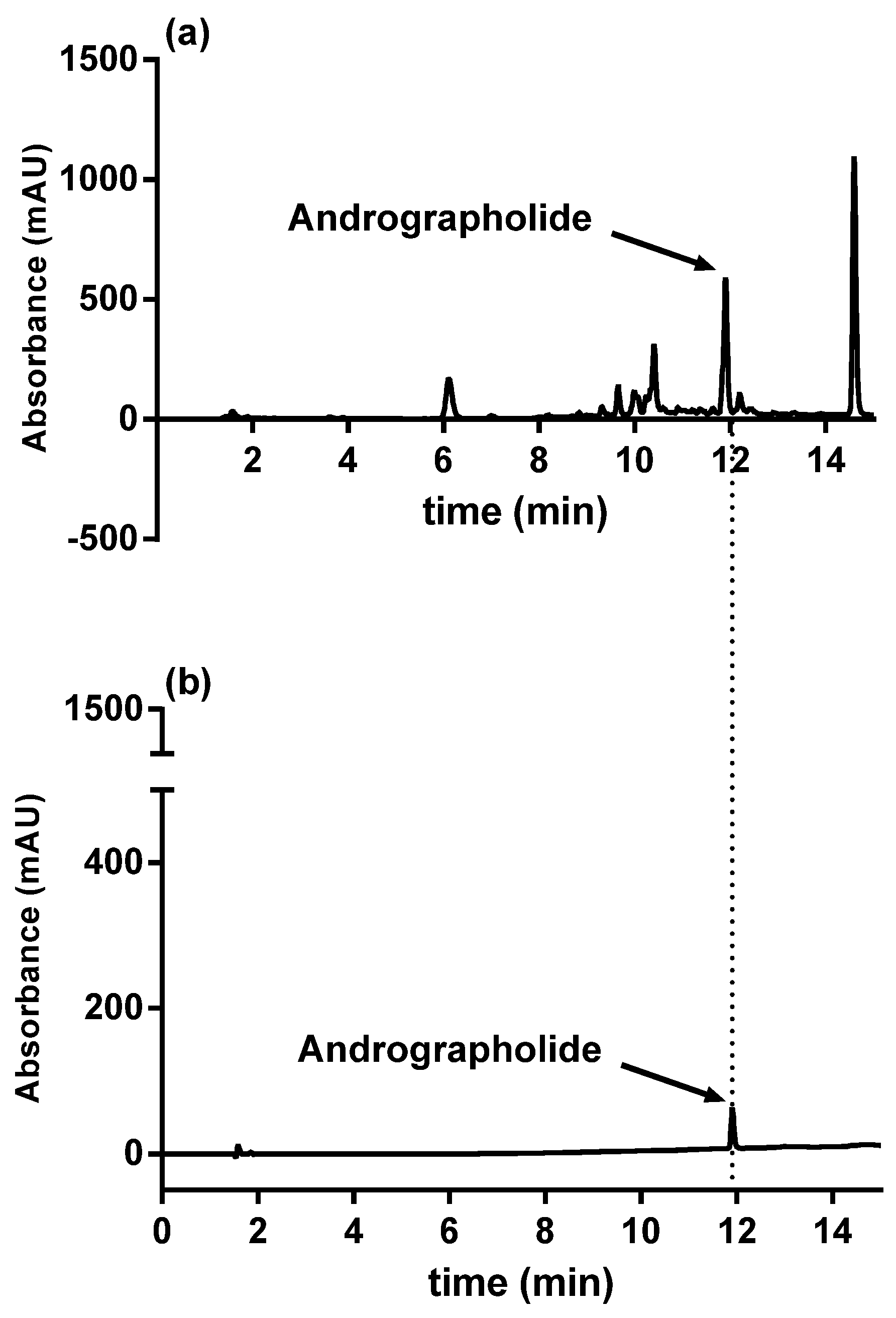
2.3. HPLC Analysis
2.4. Cell Culture
2.5. Cell Treatment
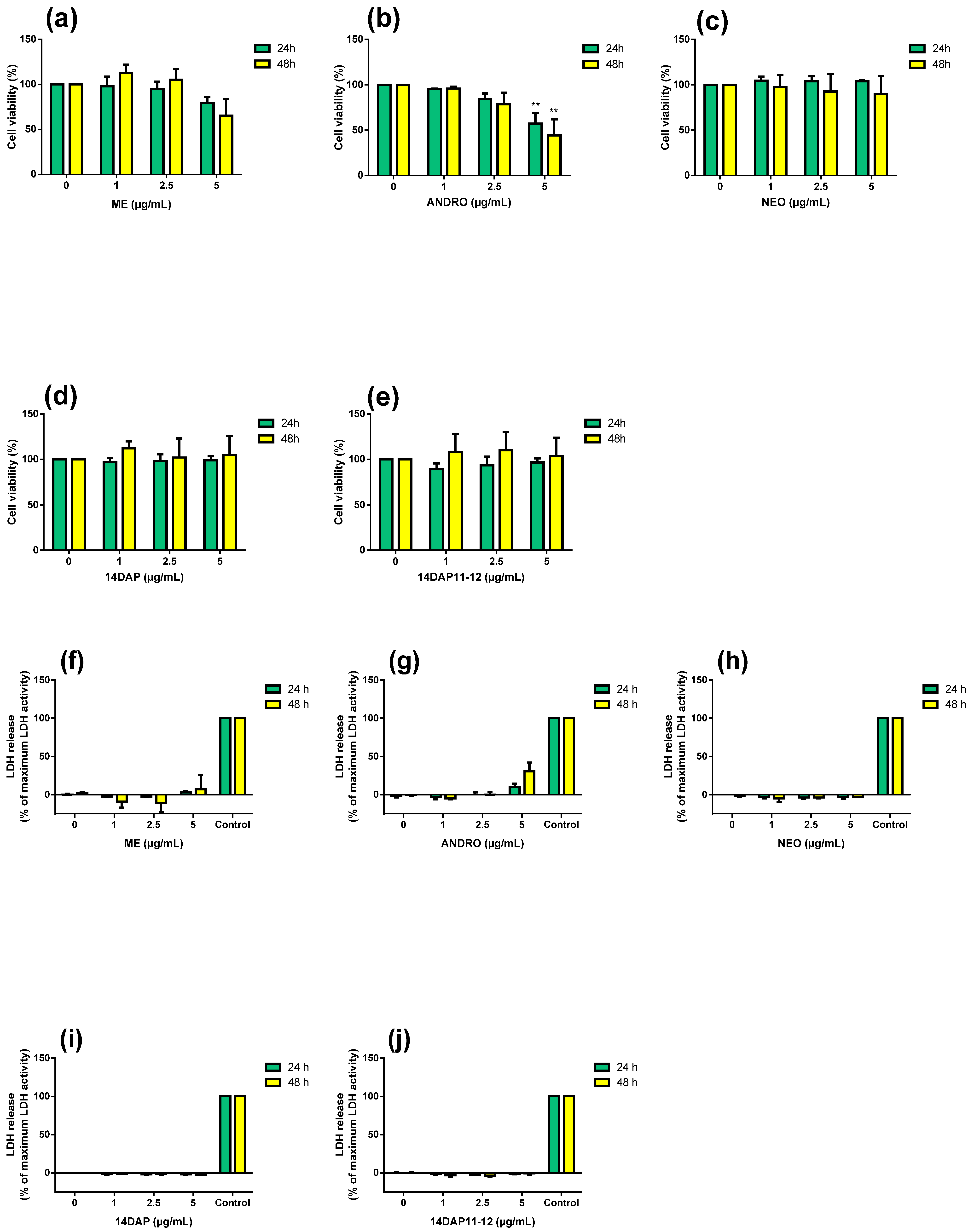
2.6. MTT Assay
2.7. Lactate Dehydrogenase Activity
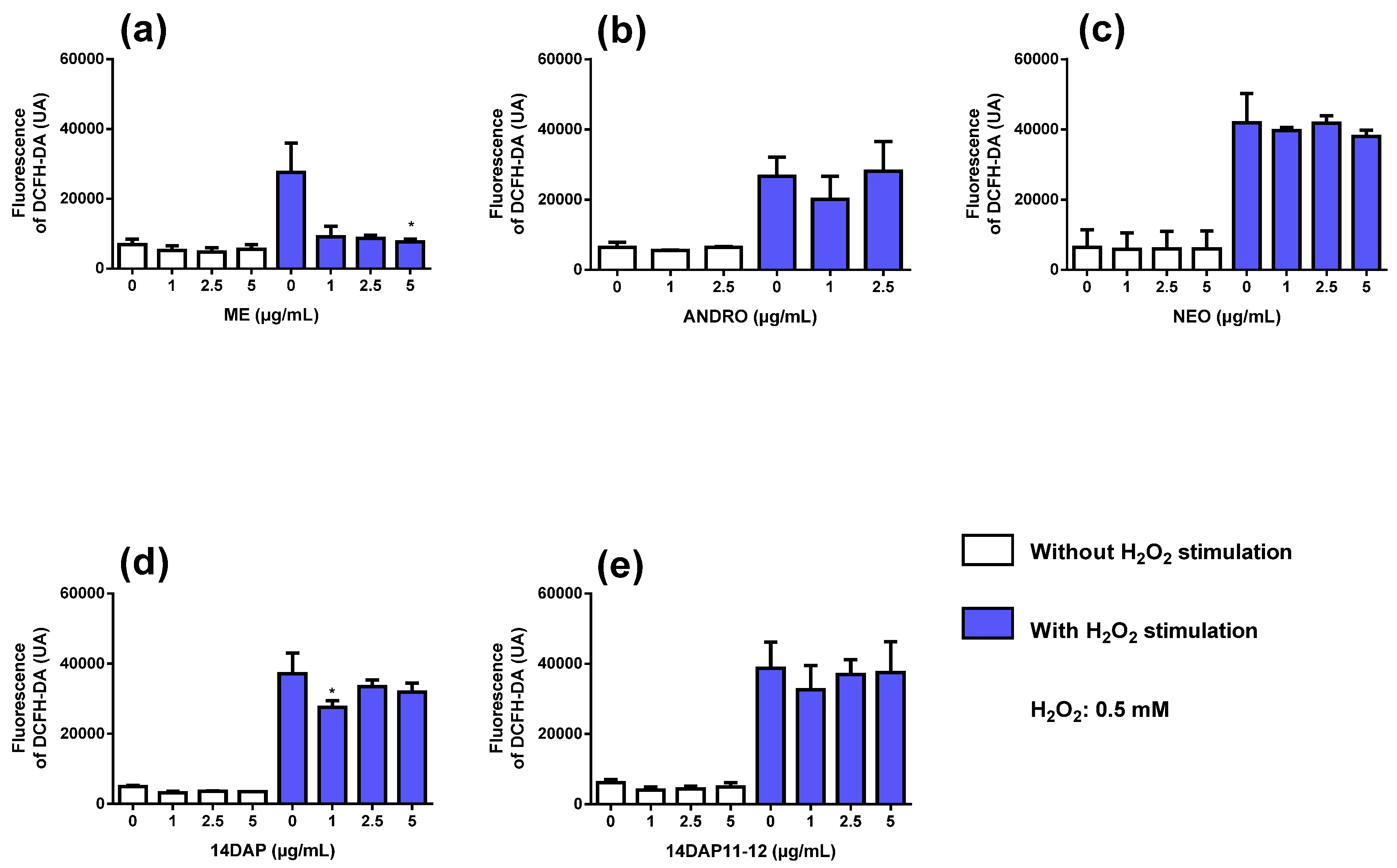
2.8. Intracellular Reactive Oxygen Species
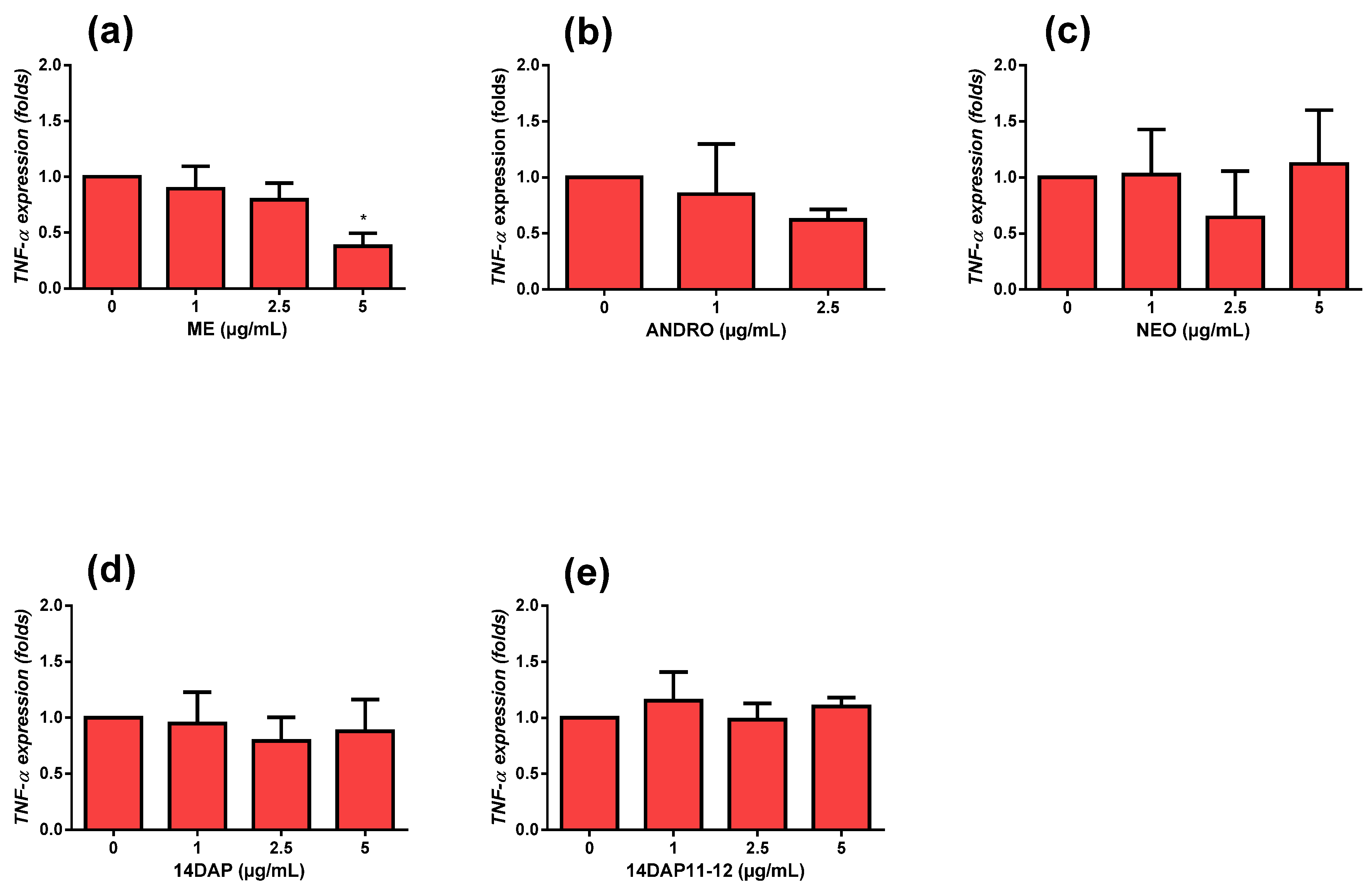
2.9. Quantitative RT-PCR
2.10. Measurement of IL-8 Secretion
2.11. Measurement of Hyaluronic Acid Secretion
2.12. Statistical Tests
3. Results
3.1. Analysis of Methanolic Extract from Andrographis paniculata
3.2. Cytotoxicity Assays
3.3. Antioxidant Activity
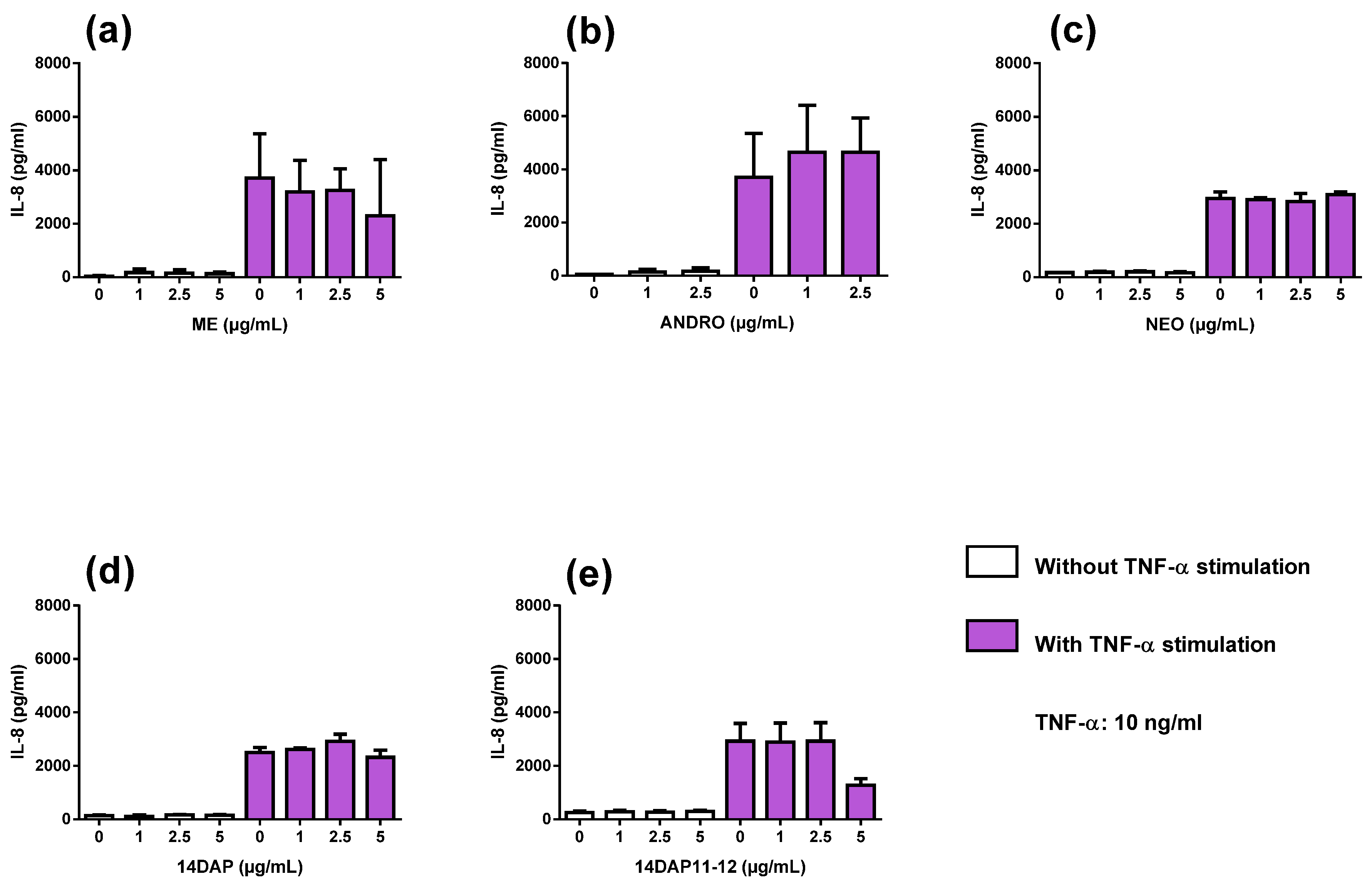
3.4. Anti-Inflammatory Activities
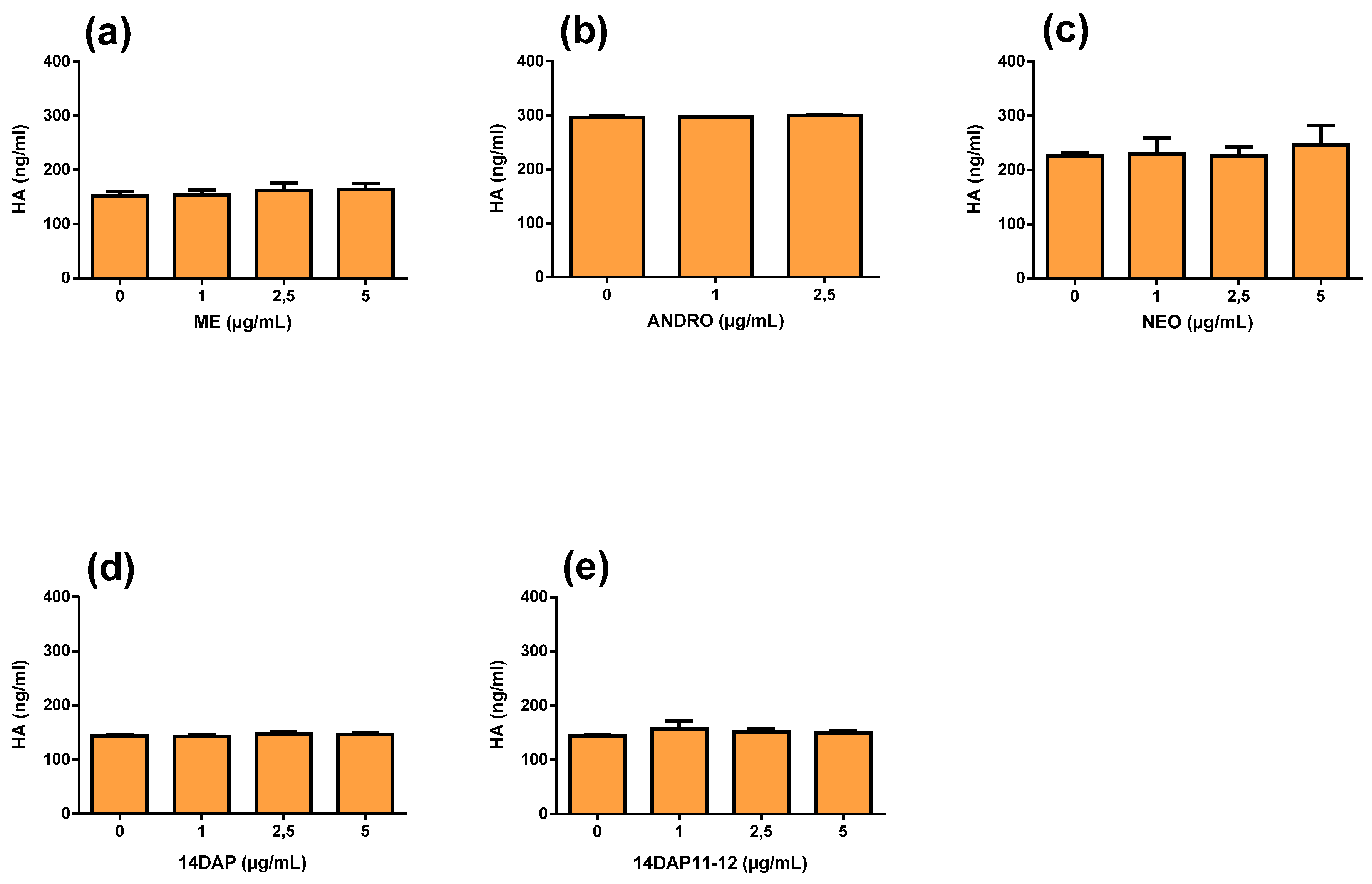
3.5. Hyaluronic Acid Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 14DAP | 14-deoxyandrographolide |
| 14DAP11-12 | 14-deoxy-11,12-didehydroandrographolide |
| ANDRO | andrographolide |
| ECM | extracellular matrix |
| HA | hyaluronic acid |
| HDFa | human dermal fibroblasts, adult |
| IL-8 | interleukin-8 |
| LDH | lactate dehydrogenase |
| ME | methanolic extract |
| MMPs | matrix metalloproteinases |
| NEO | neoandrographolide |
| ROS | reactive oxygen species |
| TNF-α | tumor necrosis factor-α |
References
- Cavinato, M.; Jansen-Dürr, P. Molecular mechanisms of UVB-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Farage, M.A.; Miller, K.W.; Elsner, P.; Maibach, H.I. Intrinsic and extrinsic factors in skin ageing: A review. Int. J. Cosmet. Sci. 2008, 30, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Oresajo, C.; Hayward, J. Ultraviolet radiation and skin aging: Roles of reactive oxygen species, inflammation and protease activation, and strategies for prevention of inflammation-induced matrix degradation—A review. Int. J. Cosmet. Sci. 2005, 27, 17–34. [Google Scholar] [CrossRef]
- Rinnerthaler, M.; Bischof, J.; Streubel, M.K.; Trost, A.; Richter, K. Oxidative Stress in Aging Human Skin. Biomolecules 2015, 5, 545–589. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-M.; Tsen, J.-H.; Yen, H.; Yang, T.-Y.; Huang, H.-C. Extract fromPeriostracum cicadaeInhibits Oxidative Stress and Inflammation Induced by Ultraviolet B Irradiation on HaCaT Keratinocytes. Evidence-Based Complement. Altern. Med. ECAM 2017, 2017, 8325049. [Google Scholar] [CrossRef] [Green Version]
- Kondo, S.; Kono, T.; Sauder, D.N.; McKenzie, R.C. IL-8 Gene Expression and Production in Human Keratinocytes and Their Modulation by UVB. J. Investig. Dermatol. 1993, 101, 690–694. [Google Scholar] [CrossRef] [Green Version]
- Morisaki, N.; Moriwaki, S.; Sugiyama-Nakagiri, Y.; Haketa, K.; Takema, Y.; Imokawa, G. Neprilysin Is Identical to Skin Fibroblast Elastase its role in skin aging and UV responses. J. Biol. Chem. 2010, 285, 39819–39827. [Google Scholar] [CrossRef] [Green Version]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinology 2012, 4, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.S.D.; Liao, W.; Zhou, S.; Wong, W.S.F. Is there a future for andrographolide to be an anti-inflammatory drug? Deciphering its major mechanisms of action. Biochem. Pharmacol. 2017, 139, 71–81. [Google Scholar] [CrossRef]
- Hussain, R.M.; Razak, Z.N.R.A.; Saad, W.M.M.; Mustakim, M. Mechanism of antagonistic effects of Andrographis paniculata methanolic extract against Staphylococcus aureus. Asian Pac. J. Trop. Med. 2017, 10, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Radhika, P.; Annapurna, A.; Rao, S.N. Immunostimulant, cerebroprotective & nootropic activities of Andrographis paniculata leaves extract in normal & type 2 diabetic rats. Indian J. Med. Res. 2012, 135, 636–641. [Google Scholar] [PubMed]
- Sheeja, K.; Shihab, P.; Kuttan, G. Antioxidant and Anti-Inflammatory Activities of the PlantAndrographis PaniculataNees. Immunopharmacol. Immunotoxicol. 2006, 28, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Mussard, E.; Jousselin, S.; Cesaro, A.; Legrain, B.; Lespessailles, E.; Esteve, E.; Berteina-Raboin, S.; Toumi, H. Andrographis paniculata and Its Bioactive Diterpenoids Protect Dermal Fibroblasts against Inflammation and Oxidative Stress. Antioxidants 2020, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Sareer, O.; Ahmad, S.; Umar, S. Andrographis paniculata: A critical appraisal of extraction, isolation and quantification of andrographolide and other active constituents. Nat. Prod. Res. 2014, 28, 2081–2101. [Google Scholar] [CrossRef]
- Mussard, E.; Cesaro, A.; Lespessailles, E.; Legrain, B.; Berteina-Raboin, S.; Toumi, H. Andrographolide, a Natural Antioxidant: An Update. Antioxidants 2019, 8, 571. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.S.D.; Liao, W.; Peh, H.Y.; Vila, M.; Dong, J.; Shen, H.-M.; Wong, W.S.F. Andrographolide simultaneously augments Nrf2 antioxidant defense and facilitates autophagic flux blockade in cigarette smoke-exposed human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2018, 360, 120–130. [Google Scholar] [CrossRef]
- Yang, M.-Y.; Yu, Q.-L.; Huang, Y.-S.; Yang, G. Neuroprotective effects of andrographolide derivative CX-10 in transient focal ischemia in rat: Involvement of Nrf2/AE and TLR/NF-κB signaling. Pharmacol. Res. 2019, 144, 227–234. [Google Scholar] [CrossRef]
- Batkhuu, J.; Hattori, K.; Takano, F.; Fushiya, S.; Oshiman, K.-I.; Fujimiya, Y. Suppression of NO production in activated macrophages in vitro and ex vivo by neoandrographolide isolated from Andrographis paniculata. Biol. Pharm. Bull. 2002, 25, 1169–1174. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, Z.; Ji, L. In Vivo and In Vitro Anti-inflammatory Activities of Neoandrographolide. Am. J. Chin. Med. 2007, 35, 317–328. [Google Scholar] [CrossRef]
- Mandal, S.; Nelson, V.K.; Mukhopadhyay, S.; Bandhopadhyay, S.; Maganti, L.; Ghoshal, N.; Sen, G.; Biswas, T. 14-Deoxyandrographolide targets adenylate cyclase and prevents ethanol-induced liver injury through constitutive NOS dependent reduced redox signaling in rats. Food Chem. Toxicol. 2013, 59, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Parichatikanond, W.; Suthisisang, C.; Dhepakson, P.; Herunsalee, A. Study of anti-inflammatory activities of the pure compounds from Andrographis paniculata (burm.f.) Nees and their effects on gene expression. Int. Immunopharmacol. 2010, 10, 1361–1373. [Google Scholar] [CrossRef] [PubMed]
- Villedieu-Percheron, E.; Ferreira, V.; Campos, J.F.; Destandau, E.; Pichon, C.; Berteina-Raboin, S. Quantitative Determination of Andrographolide and Related Compounds in Andrographis paniculata Extracts and Biological Evaluation of Their Anti-Inflammatory Activity. Foods 2019, 8, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negi, A.S.; Kumar, J.; Luqman, S.; Shanker, K.; Gupta, M.; Khanuja, S. Recent advances in plant hepatoprotectives: A chemical and biological profile of some important leads. Med. Res. Rev. 2008, 28, 746–772. [Google Scholar] [CrossRef] [PubMed]
- Kligler, B.; Ulbricht, C.; Basch, E.; Kirkwood, C.D.; Abrams, T.R.; Miranda, M.; Khalsa, K.P.S.; Giles, M.; Boon, H.; Woods, J. Andrographis Paniculata for the Treatment of Upper Respiratory Infection: A Systematic Review by the Natural Standard Research Collaboration. Explore (NY) 2006, 2, 25–29. [Google Scholar] [CrossRef]
- Jayakumar, T.; Hsieh, C.-Y.; Lee, J.-J.; Sheu, J.-R. Experimental and Clinical Pharmacology ofAndrographis paniculataand Its Major Bioactive Phytoconstituent Andrographolide. Evidence-Based Complement. Altern. Med. 2013, 2013, 846740. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.Y.-X.; Wang, X.-F.; Liu, Y.-H.; Zhang, Z.-B.; Wang, L.; Chen, J.-N.; Huang, S.; Zeng, H.-F.; Lai, X.-P. Andrographolide Sodium Bisulfate Prevents UV-Induced Skin Photoaging through Inhibiting Oxidative Stress and Inflammation. Mediat. Inflamm. 2016, 2016, 3271451. [Google Scholar] [CrossRef] [Green Version]
- Chao, W.-W.; Lin, B.-F. Isolation and identification of bioactive compounds in Andrographis paniculata (Chuanxinlian). Chin. Med. 2010, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Pholphana, N.; Rangkadilok, N.; Thongnest, S.; Ruchirawat, S.; Ruchirawat, M.; Satayavivad, J. Determination and variation of three active diterpenoids inAndrographis paniculata (Burm.f.) Nees. Phytochem. Anal. 2004, 15, 365–371. [Google Scholar] [CrossRef]
- Akowuah, G.; Zhari, I.; Mariam, A.; Yam, M. Absorption of andrographolides from Andrographis paniculata and its effect on CCl4-induced oxidative stress in rats. Food Chem. Toxicol. 2009, 47, 2321–2326. [Google Scholar] [CrossRef]
- Akowuah, G.; Zhari, I.; Norhayati, I.; Mariam, A. HPLC and HPTLC densitometric determination of andrographolides and antioxidant potential of Andrographis paniculata. J. Food Compos. Anal. 2006, 19, 118–126. [Google Scholar] [CrossRef]
- Wang, M.; Zhong, Q.; Lin, B.; Liu, Y.; Huang, Y.; Chen, Y.; Yuan, J.; Su, Z.; Zhan, J.Y. Andrographolide sodium bisulfate attenuates UV-induced photo-damage by activating the keap1/Nrf2 pathway and downregulating the NF-κB pathway in HaCaT keratinocytes. Int. J. Mol. Med. 2019, 45, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, M.; Khoo, C.S.; Müench, G.; Govindaraghavan, S.; Sucher, N. An in vitro study of anti-inflammatory activity of standardised Andrographis paniculata extracts and pure andrographolide. BMC Complement. Altern. Med. 2015, 15, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Meng, W.; Liao, W.; Lian, S. Andrographolide Antagonizes TNF-α-Induced IL-8 via Inhibition of NADPH Oxidase/ROS/NF-κB and Src/MAPKs/AP-1 Axis in Human Colorectal Cancer HCT116 Cells. J. Agric. Food Chem. 2018, 66, 5139–5148. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name. | Structure and Details |
|---|---|
| Andrographolide (ANDRO) |  |
| Neoandrographolide (NEO) |  |
| 14-deoxyandrographolide (14DAP) |  |
| 14-deoxy-11,12-didehydroandrographolide (14DAP11-12) |  |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mussard, E.; Jousselin, S.; Cesaro, A.; Legrain, B.; Lespessailles, E.; Esteve, E.; Berteina-Raboin, S.; Toumi, H. Andrographis paniculata and Its Bioactive Diterpenoids Against Inflammation and Oxidative Stress in Keratinocytes. Antioxidants 2020, 9, 530. https://doi.org/10.3390/antiox9060530
Mussard E, Jousselin S, Cesaro A, Legrain B, Lespessailles E, Esteve E, Berteina-Raboin S, Toumi H. Andrographis paniculata and Its Bioactive Diterpenoids Against Inflammation and Oxidative Stress in Keratinocytes. Antioxidants. 2020; 9(6):530. https://doi.org/10.3390/antiox9060530
Chicago/Turabian StyleMussard, Eugenie, Sundy Jousselin, Annabelle Cesaro, Brigitte Legrain, Eric Lespessailles, Eric Esteve, Sabine Berteina-Raboin, and Hechmi Toumi. 2020. "Andrographis paniculata and Its Bioactive Diterpenoids Against Inflammation and Oxidative Stress in Keratinocytes" Antioxidants 9, no. 6: 530. https://doi.org/10.3390/antiox9060530
APA StyleMussard, E., Jousselin, S., Cesaro, A., Legrain, B., Lespessailles, E., Esteve, E., Berteina-Raboin, S., & Toumi, H. (2020). Andrographis paniculata and Its Bioactive Diterpenoids Against Inflammation and Oxidative Stress in Keratinocytes. Antioxidants, 9(6), 530. https://doi.org/10.3390/antiox9060530