Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence



Abstract
:
1. Introduction
2. Carotenoids in Biomembrane: Why Different Carotenoids Behave Differently?
3. Carotenoids-Triggered ROS Production in Cancer Cells
4. Enhanced ROS as a Cancer Cure: Mechanism of Carotenoid Triggered Apoptosis
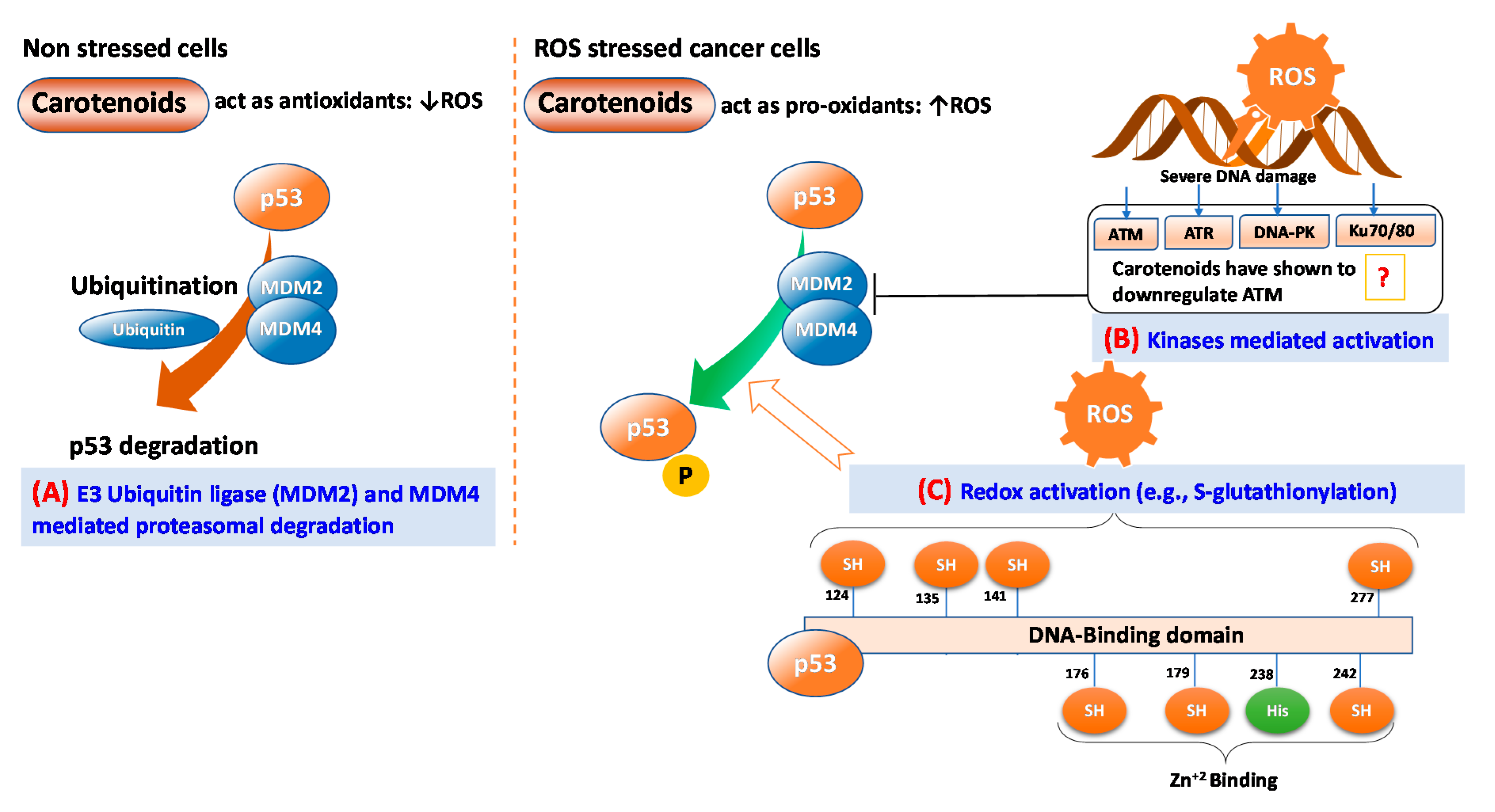
5. p53: A Key Mediator of ROS-Induced Apoptosis
6. Carotenoids Act Synergistically with Ros-Inducing Anticancer Drugs
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Milisav, I. The neglected significance of “antioxidative stress”. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthil, K.; Aranganathan, S.; Nalini, N. Evidence of oxidative stress in the circulation of ovarian cancer patients. Clin. Chim. Acta 2004, 339, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Santoro, M.M. ROS homeostasis and metabolism: A dangerous liason in cancer cells. Cell Death Dis. 2016, 7, 2253. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.; Mumper, R.J. Elevated copper and oxidative stress in cancer cells as a target for cancer treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from fruits and vegetables: Chemistry, analysis, occurrence, bioavailability and biological activities. Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Jarvik, T. Lateral transfer of genes from fungi underlies carotenoid production in aphids. Science 2010, 328, 624–627. [Google Scholar] [CrossRef] [Green Version]
- Maiani, G.; Caston, M.J.; Catasta, G.; Toti, E.; Cambrodon, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food. Res. 2009, 53, 194–218. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Al-Delaimy, W.K.; van Kappel, A.L.; Ferrari, P.; Slimani, N.; Steghens, J.P.; Bingham, S.; Johansson, I.; Wallstrom, P.; Overvad, K.; Tjonneland, A.; et al. Plasma levels of six carotenoids in nine European countries: Report from the European Prospective Investigation into Cancer and Nutrition (EPIC). Public Health Nutr. 2004, 7, 713–722. [Google Scholar] [CrossRef]
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Acad. Nutr. Diet. 2001, 101, 294. [Google Scholar]
- Toti, E.; Chen, C.Y.O.; Palmery, M.; Villaño Valencia, D.; Peluso, I. Non-Provitamin A and Provitamin A Carotenoids as Immunomodulators: Recommended Dietary Allowance, Therapeutic Index, or Personalized Nutrition? Oxid. Med. Cell. Longev. 2018, 2018, 20. [Google Scholar] [CrossRef] [PubMed]
- Lupton, J.R.; Atkinson, S.A.; Chang, N.; Fraga, C.G.; Levy, J.; Messina, M.; Richardson, D.P.; Van Ommen, B.; Yang, Y.; Griffiths, J.C.; et al. Exploring the benefits and challenges of establishing a DRI-like process for bioactives. Eur. J. Nutr. 2014, 53, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arathi, B.P.; Sowmya, P.R.; Kuriakose, G.C.; Vijay, K.; Baskaran, V.; Jayabaskaran, C.; Lakshminarayana, R. Enhanced cytotoxic and apoptosis inducing activity of lycopene oxidation products in different cancer cell lines. Food Chem. Toxicol. 2016, 97, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Gansukh, E.; Mya, K.K.; Jung, M.; Keum, Y.-S.; Kim, D.H.; Saini, R.K. Lutein derived from marigold (Tagetes erecta) petals triggers ROS generation and activates Bax and caspase-3 mediated apoptosis of human cervical carcinoma (HeLa) cells. Food Chem. Toxicol. 2019, 127, 11–18. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Structures and analysis of carotenoid molecules. In Carotenoids in Nature; Springer: Cham, Switzerland, 2016; pp. 71–108. [Google Scholar]
- Saini, R.K.; Keum, Y.S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Honda, M.; Kageyama, H.; Hibino, T.; Zhang, Y.; Diono, W.; Kanda, H.; Yamaguchi, R.; Takemura, R.; Fukaya, T.; Goto, M. Improved Carotenoid Processing with Sustainable Solvents Utilizing Z-Isomerization-Induced Alteration in Physicochemical Properties: A Review and Future Directions. Molecules 2019, 24, 2149. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Bekhit, A.E.-D.A.; Roohinejad, S.; Rengasamy, K.R.R.; Keum, Y.-S. Chemical Stability of Lycopene in Processed Products: A Review of the Effects of Processing Methods and Modern Preservation Strategies. J. Agric. Food Chem. 2020, 68, 712–726. [Google Scholar] [CrossRef]
- McNulty, H.P.; Byun, J.; Lockwood, S.F.; Jacob, R.F.; Mason, R.P. Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Lowe, G.M.; Vlismas, K.; Young, A.J. Carotenoids as prooxidants? Mol. Asp. Med. 2003, 24, 363–369. [Google Scholar] [CrossRef]
- Sowmya, P.R.-R.; Arathi, B.P.; Vijay, K.; Baskaran, V.; Lakshminarayana, R. Astaxanthin from shrimp efficiently modulates oxidative stress and allied cell death progression in MCF-7 cells treated synergistically with β-carotene and lutein from greens. Food Chem. Toxicol. 2017, 106, 58–69. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1810, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Gloria, N.F.; Soares, N.; Brand, C.; Oliveira, F.L.; Borojevic, R.; Teodoro, A.J. Lycopene and beta-carotene induce cell-cycle arrest and apoptosis in human breast cancer cell lines. Anticancer Res. 2014, 34, 1377–1386. [Google Scholar] [PubMed]
- Chisté, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids inhibit lipid peroxidation and hemoglobin oxidation, but not the depletion of glutathione induced by ROS in human erythrocytes. Life Sci. 2014, 99, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht-Jam, I.; Khademi, M.; Nosrati, M.; Eslami, S.; Foroutan-Tanha, M.; Sahebkar, A.; Tavalaie, S.; Ghayour-Mobarhan, M.; Ferns, G.A.A.; Hadizadeh, F.; et al. Effect of crocin extracted from saffron on pro-oxidant–anti-oxidant balance in subjects with metabolic syndrome: A randomized, placebo-controlled clinical trial. Eur. J. Integr. Med. 2016, 8, 307–312. [Google Scholar] [CrossRef]
- Park, J.; Chyun, J.; Kim, Y.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. 2010, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Keum, Y.-S.; Daglia, M.; Rengasamy, K.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Saini, R.K.; Rengasamy, K.R.R.; Mahomoodally, F.M.; Keum, Y.-S. Protective effects of lycopene in cancer, cardiovascular, and neurodegenerative diseases: An update on epidemiological and mechanistic perspectives. Pharmacol. Res. 2020, 155, 104730. [Google Scholar] [CrossRef]
- El-Agamey, A.; McGarvey, D.J. The reactivity of carotenoid radicals with oxygen. Free Radic. Res. 2007, 41, 295–302. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Beutner, S.; Bloedorn, B.; Frixel, S.; Hernández Blanco, I.; Hoffmann, T.; Martin, H.-D.; Mayer, B.; Noack, P.; Ruck, C.; Schmidt, M.; et al. Quantitative assessment of antioxidant properties of natural colorants and phytochemicals: Carotenoids, flavonoids, phenols and indigoids. The role of β-carotene in antioxidant functions. J. Sci. Food Agric. 2001, 81, 559–568. [Google Scholar] [CrossRef]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids that Exhibit both Anti- and Pro-Oxidative Mechanisms—A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jomova, K.; Valko, M. Health protective effects of carotenoids and their interactions with other biological antioxidants. Eur. J. Med. Chem. 2013, 70, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.D. Do carotenoids serve as transmembrane radical channels? Free Radic. Biol. Med. 2009, 47, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Arathi, B.P.; Raghavendra-Rao Sowmya, P.; Kuriakose, G.C.; Shilpa, S.; Shwetha, H.J.; Kumar, S.; Raju, M.; Baskaran, V.; Lakshminarayana, R. Fractionation and Characterization of Lycopene-Oxidation Products by LC-MS/MS (ESI)(+): Elucidation of the Chemopreventative Potency of Oxidized Lycopene in Breast-Cancer Cell Lines. J. Agric. Food Chem. 2018, 66, 11362–11371. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef]
- Chen, Q.; Espey, M.G.; Sun, A.Y.; Pooput, C.; Kirk, K.L.; Krishna, M.C.; Khosh, D.B.; Drisko, J.; Levine, M. Pharmacologic doses of ascorbate act as a prooxidant and decrease growth of aggressive tumor xenografts in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 11105–11109. [Google Scholar] [CrossRef] [Green Version]
- Carocho, M.; Ferreira, I. A review on antioxidants, prooxidants and related controversy: Natural and synthetic compounds, screening and analysis methodologies and future perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef]
- Yeum, K.-J.; Aldini, G.; Russell, R.M.; Krinsky, N.I. Antioxidant/pro-oxidant actions of carotenoids. In Carotenoids; Springer: Cham, Switzerland, 2009; pp. 235–268. [Google Scholar]
- Siems, W.; Wiswedel, I.; Salerno, C.; Crifò, C.; Augustin, W.; Schild, L.; Langhans, C.-D.; Sommerburg, O. β-Carotene breakdown products may impair mitochondrial functions—potential side effects of high-dose β-carotene supplementation. J. Nutr. Biochem. 2005, 16, 385–397. [Google Scholar] [CrossRef]
- Vrolijk, M.F.; Opperhuizen, A.; Jansen, E.H.J.M.; Godschalk, R.W.; Van Schooten, F.J.; Bast, A.; Haenen, G.R.M.M. The shifting perception on antioxidants: The case of vitamin E and β-carotene. Redox Biol. 2015, 4, 272–278. [Google Scholar] [CrossRef] [Green Version]
- Reczek, C.R.; Chandel, N.S. The Two Faces of Reactive Oxygen Species in Cancer. Annu. Rev. Cancer Biol. 2017, 1, 79–98. [Google Scholar] [CrossRef]
- De Oliveira Júnior, R.G.; Bonnet, A.; Braconnier, E.; Groult, H.; Prunier, G.; Beaugeard, L.; Grougnet, R.; da Silva Almeida, J.R.G.; Ferraz, C.A.A.; Picot, L. Bixin, an apocarotenoid isolated from Bixa orellana L., sensitizes human melanoma cells to dacarbazine-induced apoptosis through ROS-mediated cytotoxicity. Food Chem. Toxicol. 2019, 125, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Gansukh, E.; Nile, A.; Sivanesan, I.; Rengasamy, K.R.R.; Kim, D.H.; Keum, Y.S.; Saini, R.K. Chemopreventive Effect of beta-Cryptoxanthin on Human Cervical Carcinoma (HeLa) Cells Is Modulated through Oxidative Stress-Induced Apoptosis. Antioxidants 2019, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-N.; Heo, S.-J.; Kang, S.-M.; Ahn, G.; Jeon, Y.-J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. In Vitro 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Wu, H.-L.; Fu, X.-Y.; Cao, W.-Q.; Xiang, W.-Z.; Hou, Y.-J.; Ma, J.-K.; Wang, Y.; Fan, C.-D. Induction of Apoptosis in Human Glioma Cells by Fucoxanthin via Triggering of ROS-Mediated Oxidative Damage and Regulation of MAPKs and PI3K–AKT Pathways. J. Agric. Food Chem. 2019, 67, 2212–2219. [Google Scholar] [CrossRef]
- Kavalappa, Y.P.; Udayawara Rudresh, D.; Gopal, S.S.; Haranahalli Shivarudrappa, A.; Stephen, N.M.; Rangiah, K.; Ponesakki, G. β-carotene isolated from the marine red alga, Gracillaria sp. potently attenuates the growth of human hepatocellular carcinoma (HepG2) cells by modulating multiple molecular pathways. J. Funct. Foods 2019, 52, 165–176. [Google Scholar] [CrossRef]
- Vijay, K.; Sowmya, P.R.-R.; Arathi, B.P.; Shilpa, S.; Shwetha, H.J.; Raju, M.; Baskaran, V.; Lakshminarayana, R. Low-dose doxorubicin with carotenoids selectively alters redox status and upregulates oxidative stress-mediated apoptosis in breast cancer cells. Food Chem. Toxicol. 2018, 118, 675–690. [Google Scholar] [CrossRef]
- Gong, X.; Smith, J.; Swanson, H.; Rubin, L. Carotenoid Lutein Selectively Inhibits Breast Cancer Cell Growth and Potentiates the Effect of Chemotherapeutic Agents through ROS-Mediated Mechanisms. Molecules 2018, 23, 905. [Google Scholar] [CrossRef] [Green Version]
- Ezeriņa, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-Acetyl Cysteine Functions as a Fast-Acting Antioxidant by Triggering Intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 2018, 25, 447–459.e4. [Google Scholar] [CrossRef] [Green Version]
- Kotake-Nara, E.; Terasaki, M.; Nagao, A. Characterization of apoptosis induced by fucoxanthin in human promyelocytic leukemia cells. Biosci. Biotechnol. Biochem. 2005, 69, 224–227. [Google Scholar] [CrossRef]
- Liu, B.; Chen, Y.; Clair, D.K.S. ROS and p53: A versatile partnership. Free Radic. Biol. Med. 2008, 44, 1529–1535. [Google Scholar] [CrossRef] [Green Version]
- Muller, P.A.J.; Vousden, K.H. Mutant p53 in Cancer: New Functions and Therapeutic Opportunities. Cancer Cell 2014, 25, 304–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitolli, C.; Wang, Y.; Candi, E.; Shi, Y.; Melino, G.; Amelio, I. P53-Mediated Tumor Suppression: DNA-Damage Response and Alternative Mechanisms. Cancers 2019, 11, 1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogosawa, S.; Yoshida, K. Tumor suppressive role for kinases phosphorylating p53 in DNA damage-induced apoptosis. Cancer Sci. 2018, 109, 3376–3382. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menolfi, D.; Zha, S. ATM, ATR and DNA-PKcs kinases—The lessons from the mouse models: Inhibition ≠ deletion. Cell Biosci. 2020, 10. [Google Scholar] [CrossRef]
- Jang, S.H.; Lim, J.W.; Kim, H. Mechanism of β-Carotene-Induced Apoptosis of Gastric Cancer Cells: Involvement of Ataxia-Telangiectasia-Mutated. Ann. N. Y. Acad. Sci. 2009, 1171, 156–162. [Google Scholar] [CrossRef]
- Takeshima, M.; Ono, M.; Higuchi, T.; Chen, C.; Hara, T.; Nakano, S. Anti-proliferative and apoptosis-inducing activity of lycopene against three subtypes of human breast cancer cell lines. Cancer Sci. 2014, 105, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Sowmya Shree, G.; Prasad, K.Y.; Arpitha, H.; Deepika, U.; Kumar, K.N.; Mondal, P.; Ganesan, P. β-carotene at physiologically attainable concentration induces apoptosis and down-regulates cell survival and antioxidant markers in human breast cancer (MCF-7) cells. Mol. Cell. Biochem. 2017, 436, 1–12. [Google Scholar] [CrossRef]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Choi, J.; Lim, J.W.; Kim, H. β-Carotene-induced apoptosis is mediated with loss of Ku proteins in gastric cancer AGS cells. Genes Nutr. 2015, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular Advances and Pharmacologic Developments in Antitumor Activity and Cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, D.B.S. Ser392 Phosphorylation Regulates the Oncogenic Function of Mutant p53. Cancer Res. 2004, 64, 4749–4754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda-Vilela, A.L.; Grisolia, C.K.; Longo, J.P.F.; Peixoto, R.C.A.; de Almeida, M.C.; Barbosa, L.C.P.; Roll, M.M.; Portilho, F.A.; Estevanato, L.L.C.; Bocca, A.L.; et al. Oil rich in carotenoids instead of vitamins C and E as a better option to reduce doxorubicin-induced damage to normal cells of Ehrlich tumor-bearing mice: Hematological, toxicological and histopathological evaluations. J. Nutr. Biochem. 2014, 25, 1161–1176. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental System | Carotenoid | Major Outcome | Conclusion | References |
|---|---|---|---|---|
| U251-human-glioma-cell | Fucoxanthin (20 μM) | ↑ROS, ↑DNA damage, ↑MAPKs, ↑Ser428-ATR, Ser1981-ATM, Ser15-p53, and Ser139-histone ↑Thr183-JNK, Thr180-p38, ↓Ser-AKT, ↓PI3K−AKT | Fucoxanthin-induced oxidation of thiol-containing intracellular antioxidant (e.g., GSH and thioredoxin) or blocked the activities of antioxidant enzymes | [47] |
| Human hepatocellular carcinoma (HepG2) cells | β-carotene (1–5 μM) | ↑MOMP, ↓Bcl-2, PARP, and NF-kB, ↓Akt and ERK1/2, ↓SOD-2 and HO-1, Nrf-2, ↑Bax and cleaved PARP, (  ) ROS ) ROS | β-carotene suppresses the growth of HepG2 cells by activating the intrinsic apoptotic pathway modulated by intracellular antioxidant status (independent of ROS induction) | [48] |
| Human melanoma A2058 cells | Bixin (50–100) µM, IC50 of 31.85 µM with bixin + dacarbazine | ↑ROS and MDA (a lipid peroxidation marker) | Bixin sensitizes A2058 cells to dacarbazine-induced cytotoxicity through ROS elevations | [44] |
| HeLa cells | Lutein (1 to 10 µM), IC50 of 7.9 and 3.7 µM after 24 and 48 h of treatments | ↑ROS (three times higher ROS in cell treated with 10 µM, compared to control), ↑Bax, p53, caspase-3 mRNA, ↑nuclear DNA damage (97% TUNEL-positive cells in cell treated with 10 µM), ↓Bcl-2 | Lutein triggers a ROS-mediated intrinsic apoptotic pathway in HeLa cells | [15] |
| HeLa cells | β-Cryptoxanthin (1 to 10 µM), IC50 of 4.5 and 3.7 µM after 24 and 48h of treatments | ↑ROS, ↑caspase-3, -7, and -9, p53, and Bax mRNA, ↑cleaved caspase-3, ↓Bcl-2 mRNA, ↑MOMP, ↑nuclear DNA damage (52% TUNEL-positive cells in cell treated with 1 µM) | β-Cryptoxanthin triggers ROS-mediated intrinsic apoptotic pathway in HeLa cells | [45] |
| Breast-cancer MCF-7 cell lines | Autoxidation or KMnO4 oxidized products of lycopene, mainly apo-8,6′-carotendial in fraction II of silica column (IC50 value of 64.5 μM) | ↑ROS, ↑mitochondrial dysfunction | ROS-dependent (at 50 µM concentration) and ROS-independent (at higher concentrations of 100 µM) apoptosis of MCF-7 cells | [36] |
| Estrogen receptor (ER)-positive MCF-7 and ER-negative MDA-MB-231 cells | Doxorubicin (0.2–3.2 µM) + carotenoids (lutein, astaxanthin, β-carotene, and fucoxanthin; 2–10 µM) | ↓GSH (29.5–48.8%), ↑LPx, ↑ROS (43.2–65%), ↑mitochondrial dysfunction, ↓Bcl-2, ↑Bax, p53, caspase -3, -8, and -9), p21, and, p27, Cytotoxicity to MCF-7 cells: β-carotene > lutein > fucoxanthin > astxanthin | Carotenoids deliver the synergetic effects towards the cytotoxic killing of cancer cells by ROS-inducing anti-cancer drugs without influencing the redox status and proliferation of normal breast epithelial MCF 10A cells | [49] |
| Human prostate (PC-3), cervical (HeLa), and breast adenocarcinoma (MCF-7) cells | Autoxidation or chemically (induced by KMnO4) oxidized products (e.g., Apo-1,6’-carotendial and Apo-5,6’-carotendial) of lycopene (1–50 μM) | ↑DNA condensations, ↑ROS, ↑MDA, ↓GSH | Chemically oxidized products are more efficient than autoxidation products in ROS generation and enhanced apoptosis of cancer cells | [14] |
| HL-60 | Fucoxanthin (8–30 µM) | ↑ROS (fluorescence intensity from 99 (vehicle-treated cells) to 132 (correspondent to superoxide anion, O2•−), ↑cleaved caspases-3 and -7, PARP, ↓Bcl-xL proteins | ROS-induced enhanced cleavage of caspases-3 and -7, and PARP | [46] |
| MCF-7 | Astaxanthin co-treatment with β-carotene and lutein (equimolar 5 µM each) | ↑ROS, MDA (≈2.5 fold higher), ↓GSH (≈2 fold lower), ↑p53 and Bax proteins, ↓Bcl-2 proteins | Astaxanthin works synergistically with β-carotene and lutein to trigger ROS production and the apoptosis of MCF-7 cells | [22] |
| MCF-7 and triple-negative MDA-MB-468 breast cancer cells | Lutein (2 µM) + chemotherapeutic drugs (e.g., taxanes, paclitaxel and docetaxel; 0.5 µM). | ↑ROS (1.9-fold in cells treated with 2 µM lutein compared to control),p53 protein, ↑phosphorylated p53 (at serine residues Ser15, Ser46, and Ser392), ↑cellular HSP60 | Lutein inhibits the proliferation of breast cancer cells through ROS-mediated activation of p53 protein | [50] |
arrow. Akt: Protein kinase B (PKB); ATM: ataxia telangiectasia mutated; ATR: ataxia telangiectasia and Rad3 related (a serine/threonine-protein kinase); Bax: Bcl-2-associated X protein; Bcl-2: B-cell lymphoma 2; Bcl-xL: Bcl-extra-large; DNA-PK: DNA-dependent protein kinases; ERK: signal-regulated kinases; GSH: Reduced glutathione; HO-1: Heme oxygenase-1; HSP: Heat shock protein; JNK: c-Jun N-terminal kinases; LPx: Lipid peroxides; MAPKs mitogen-activated protein kinases; MDA: Malondialdehyde; MOMP: Mitochondrial outer membrane permeabilization; NF-kB: Nuclear factor-κB; Nrf-2: Nuclear factor erythroid 2–related factor 2; PARP: Poly (ADP-ribose) polymerase; PI3K: Phosphoinositide 3-kinases; SOD: Superoxide dismutase.© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, J.; Song, M.-H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532. https://doi.org/10.3390/antiox9060532
Shin J, Song M-H, Oh J-W, Keum Y-S, Saini RK. Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants. 2020; 9(6):532. https://doi.org/10.3390/antiox9060532
Chicago/Turabian StyleShin, Juhyun, Min-Ho Song, Jae-Wook Oh, Young-Soo Keum, and Ramesh Kumar Saini. 2020. "Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence" Antioxidants 9, no. 6: 532. https://doi.org/10.3390/antiox9060532
APA StyleShin, J., Song, M.-H., Oh, J.-W., Keum, Y.-S., & Saini, R. K. (2020). Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants, 9(6), 532. https://doi.org/10.3390/antiox9060532