
Comparative Transcriptome Profiling of mRNA and lncRNA of Mouse Spleens Inoculated with the Group ACYW135 Meningococcal Polysaccharide Vaccine

 ,
,
Abstract
:1. Introduction
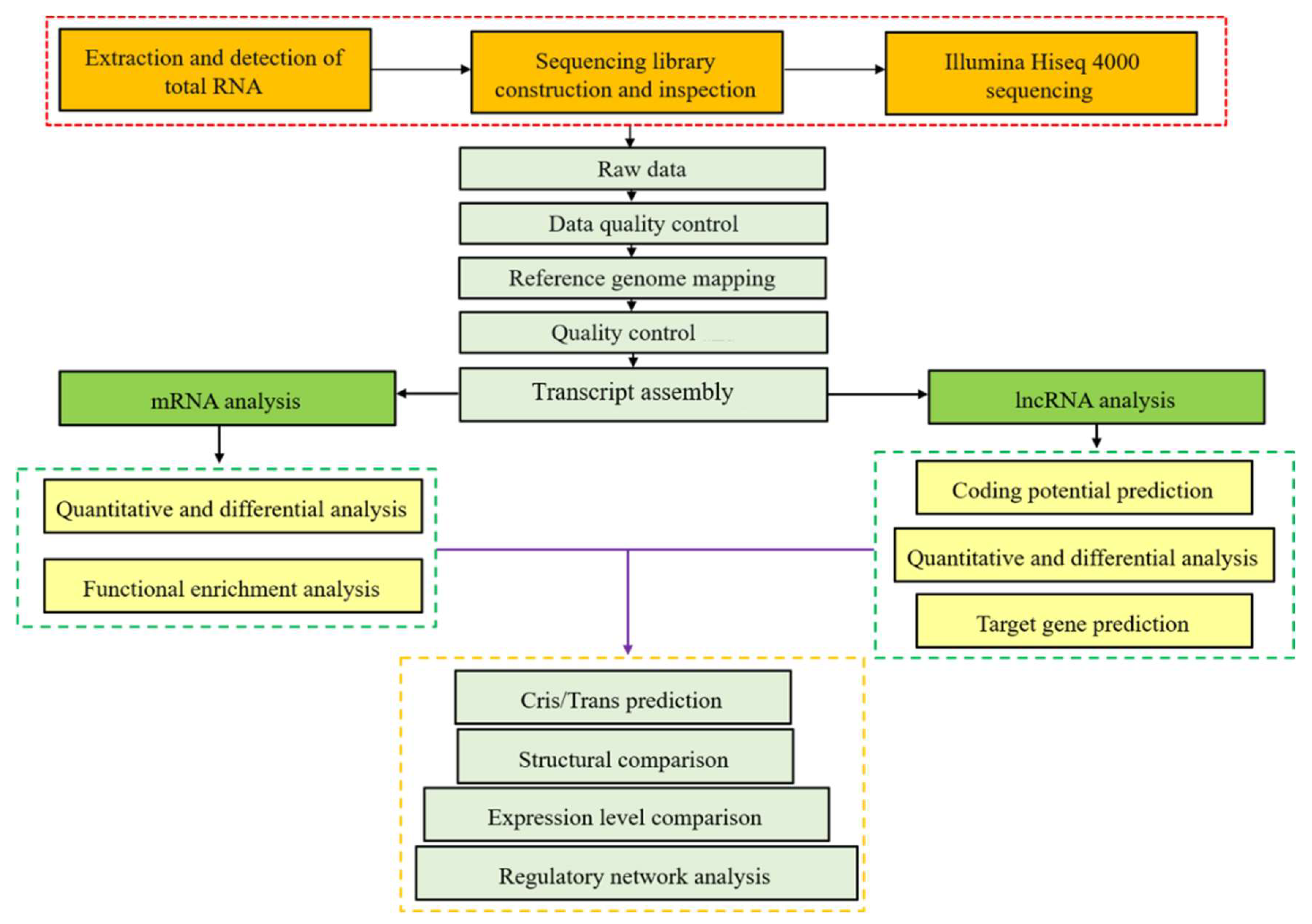
2. Materials and Methods
2.1. Animal Treatment and Sample Collection
2.2. RNA Isolation, Library Preparation and Sequencing
2.3. Quality Control and Mapping
2.4. Identification of lncRNA
2.5. DE mRNAs and DE lncRNAs Identification
2.6. Target Gene Prediction of lncRNAs and Functional Analysis
2.7. Results of GO Enrichment and KEGG Enrichment Analysis
2.8. RNA Sequencing Result Validation by RT-qPCR
2.9. Statistical Analyses
3. Results
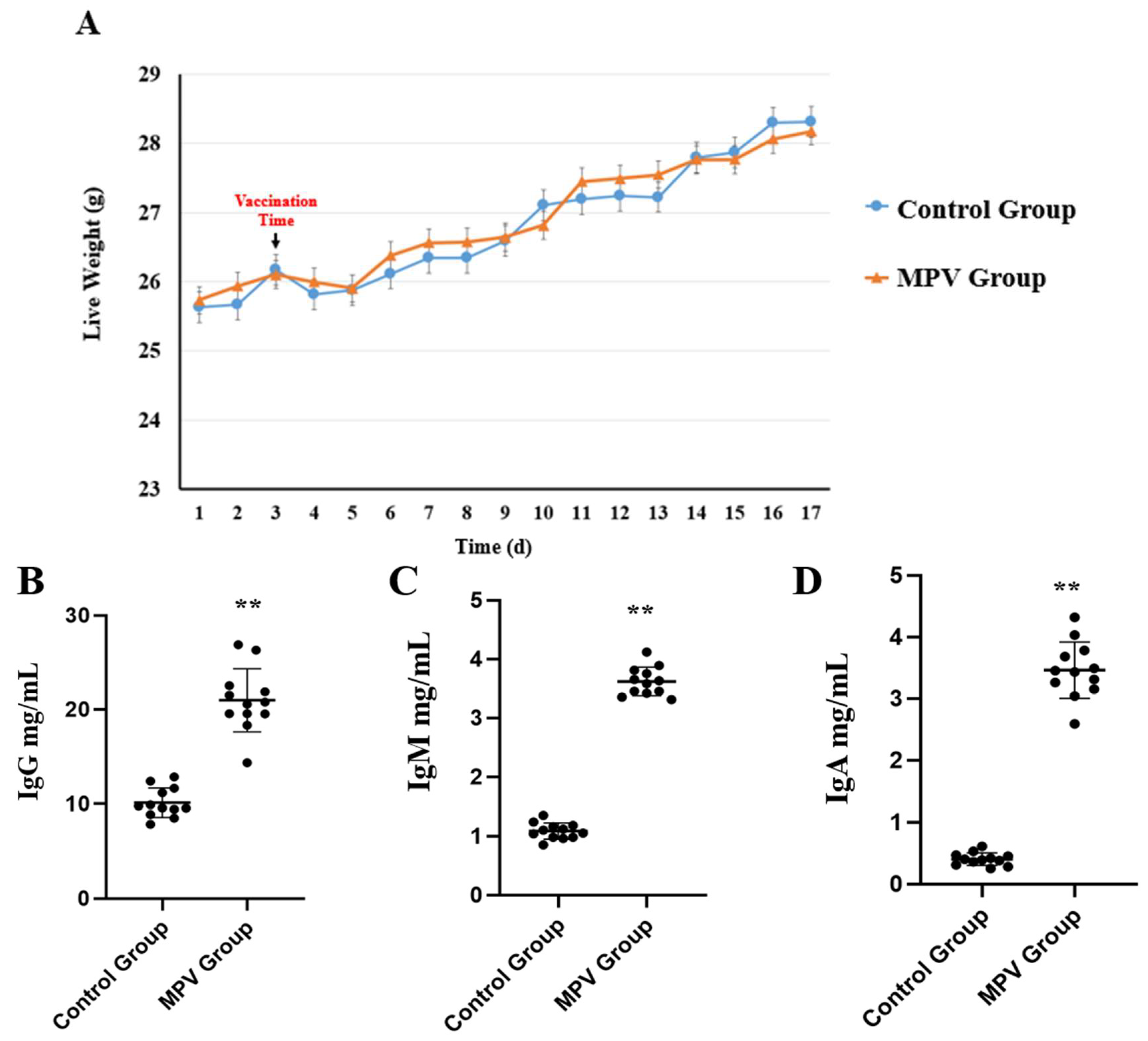
3.1. Phenotypic Data Analysis
3.2. Sequencing Data Summary
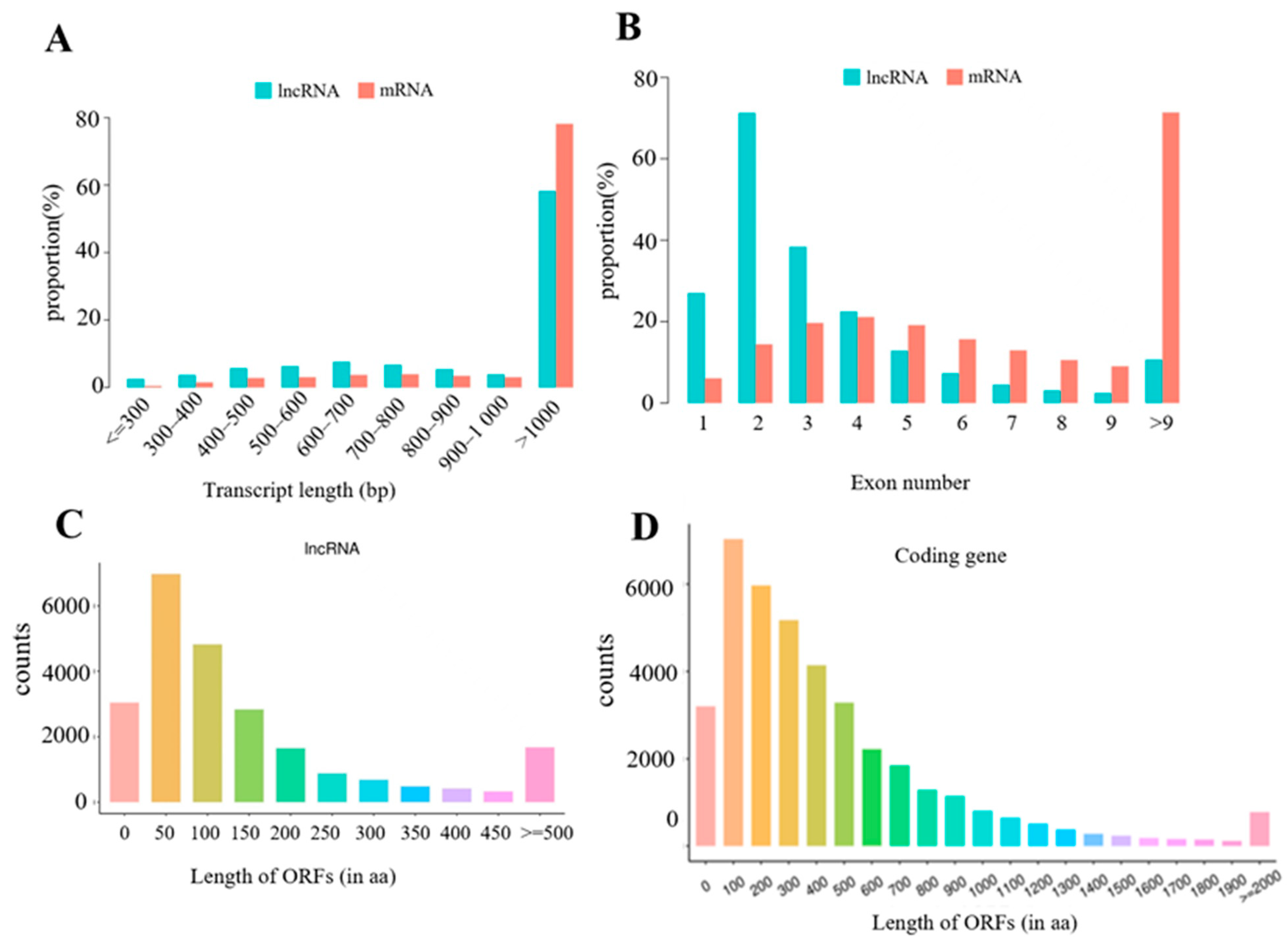
3.3. Identification of lncRNAs and mRNAs in Mouse Spleens
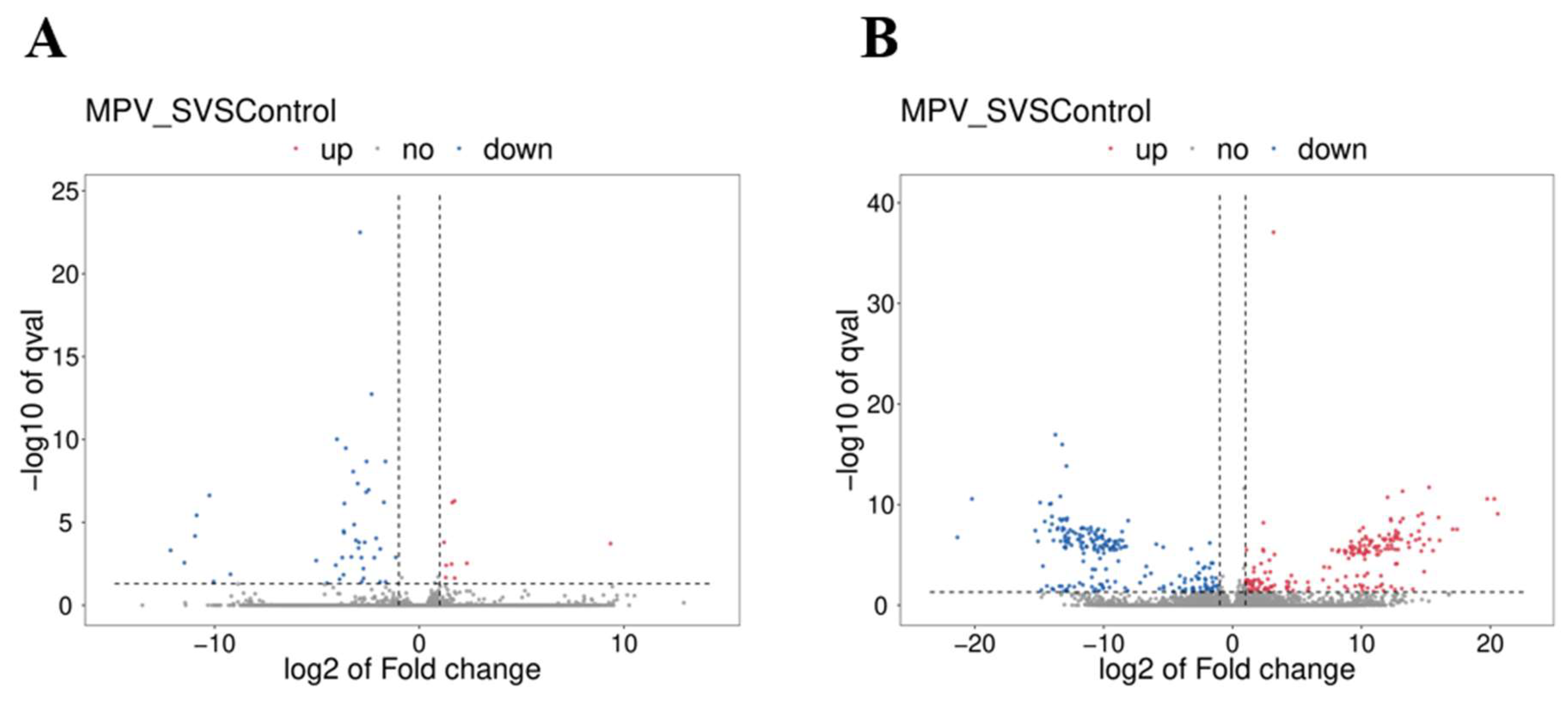
3.4. Determination of DE mRNAs and DE lncRNAs
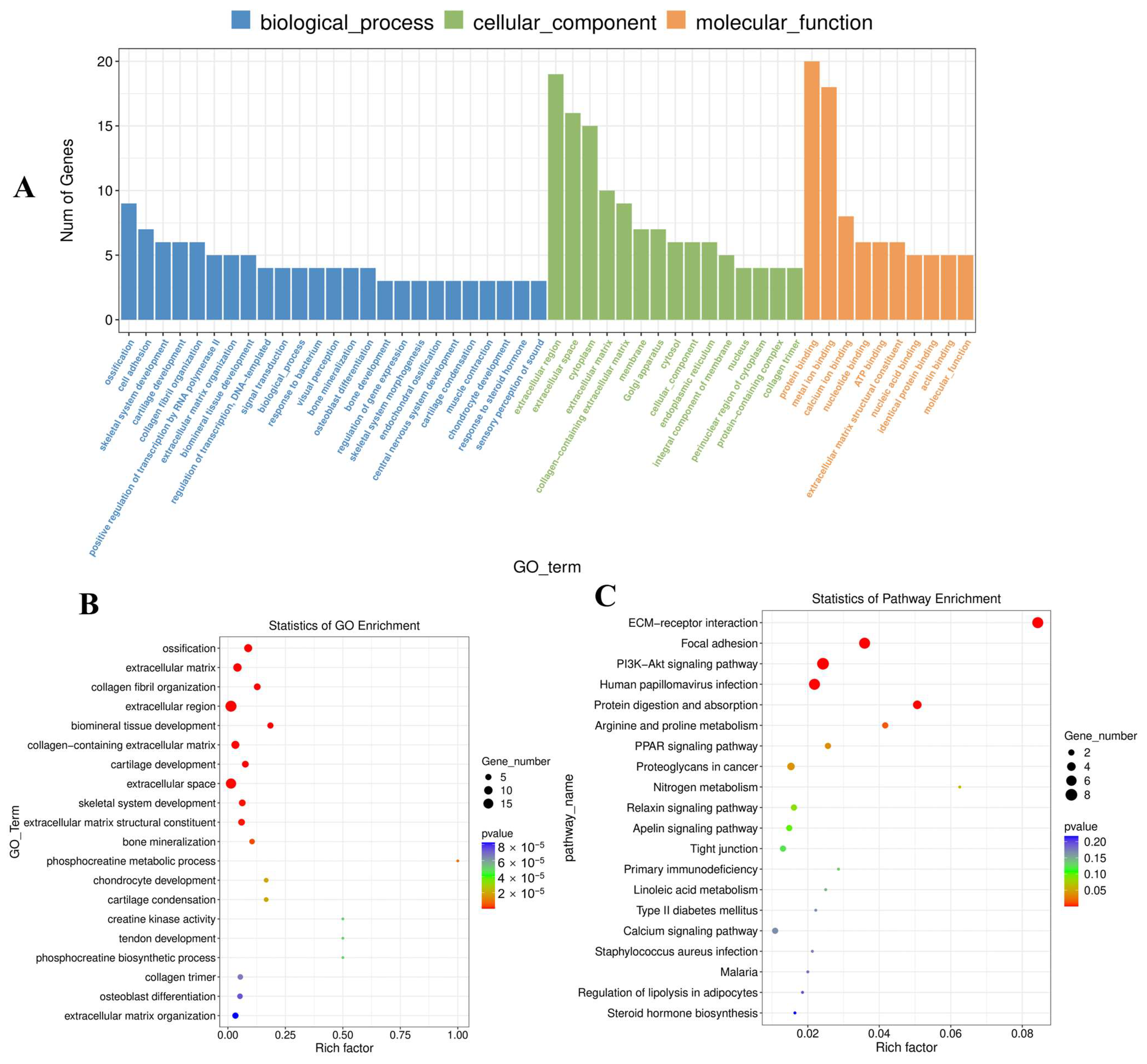
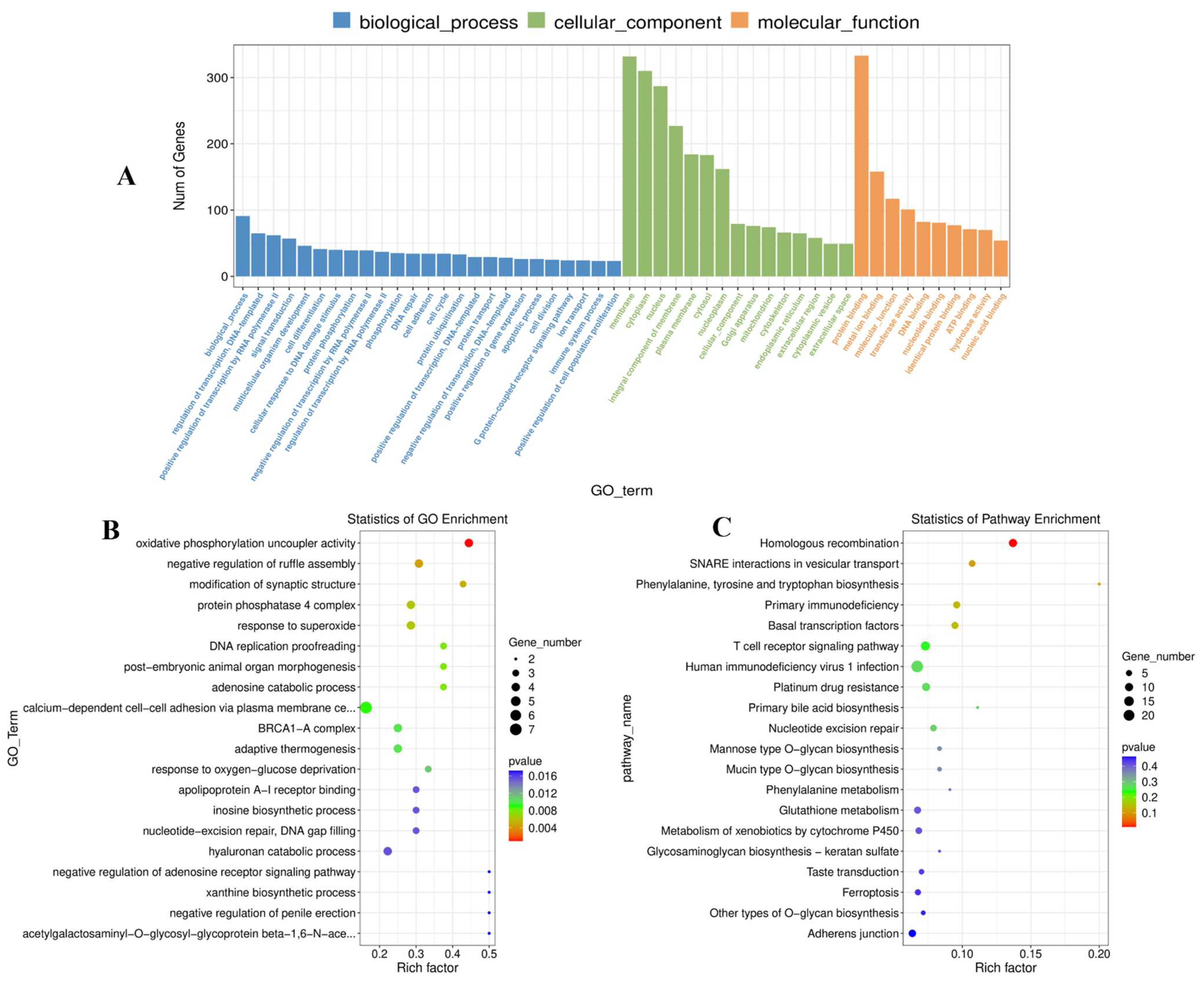
3.5. DE mRNA Functional Enrichment
3.6. Cis-Regulatory Functions of DE lncRNAs in Mouse Spleen Tissues
3.7. Analysis of DE lncRNAs and DE mRNAs by Co-Enriched GO Terms
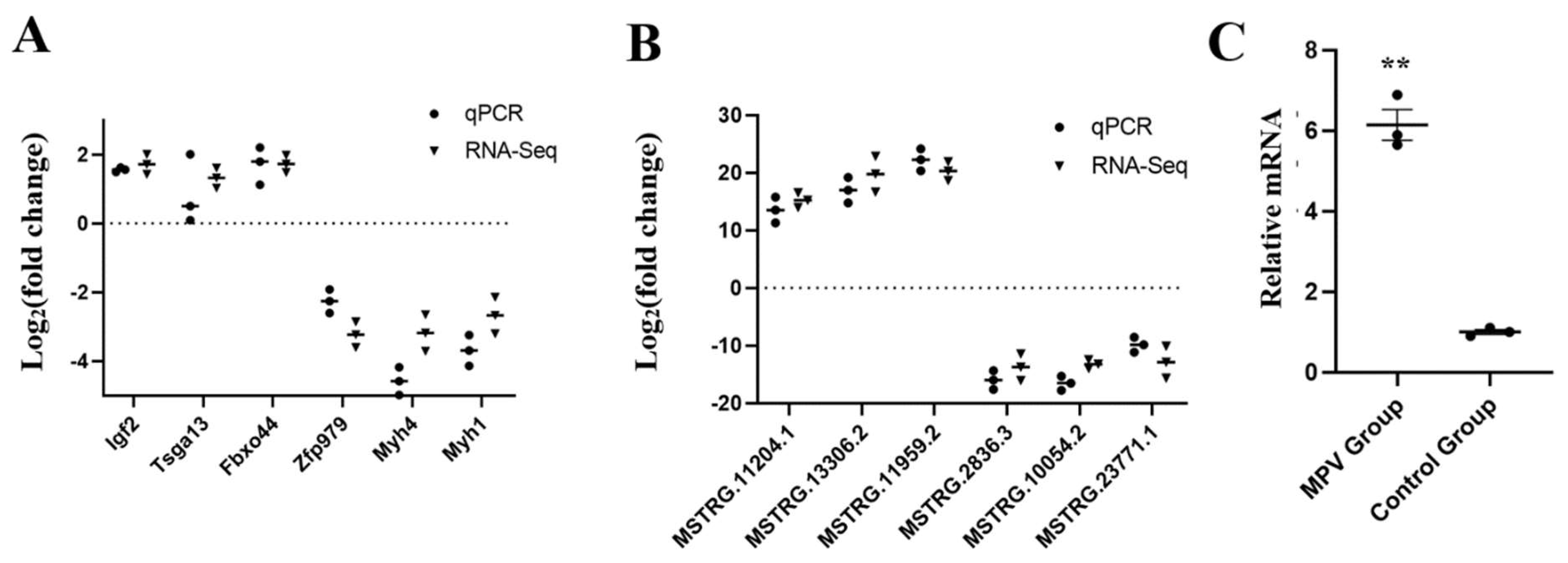
3.8. DE lncRNAs and DE mRNAs Verification Using RT-qPCR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zahlanie, Y.C.; Hammadi, M.M.; Ghanem, S.T.; Dbaibo, G.S. Review of meningococcal vaccines with updates on immunization in adults. Hum. Vacc. Immunother. 2014, 10, 995–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhd Yusoff, N.H.; Zainol Rashid, Z.; Sulong, A.; Shafiee, M.N.; Ismail, Z. Cellulitis: An unusual manifestation of Neisseria meningitidis infection. Malays. J. Pathol. 2020, 41, 315–354. [Google Scholar]
- Mikucki, A.; McCluskey, N.R.; Kahler, C.M. The Host-Pathogen Interactions and Epicellular Lifestyle of Neisseria meningitidis. Front. Cell. Infect. Microbiol. 2022, 12, 862935. [Google Scholar] [CrossRef]
- Caugant, D.A.; Brynildsrud, O.B. Neisseria meningitidis: Using genomics to understand diversity, evolution and pathogenesis. Nat. Rev. Microbiol. 2020, 18, 84–96. [Google Scholar] [CrossRef]
- Ghimire, S.; McCarthy, P.C. Capture of Pb2+ and Cu2+ Metal Cations by Neisseria meningitidis-type Capsular Polysaccharides. Biomolecules 2018, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, D.S.; Greenwood, B.; Brandtzaeg, P. Epidemic meningitis, meningococcaemia, and Neisseria meningitidis. Lancet 2007, 369, 2196–2210. [Google Scholar] [CrossRef] [PubMed]
- van de Beek, D.; Brouwer, M.C.; Thwaites, G.E.; Tunkel, A.R. Advances in treatment of bacterial meningitis. Lancet 2012, 380, 1693–1702. [Google Scholar] [CrossRef]
- Nadine, G.R.; David, S.S. Neisseria meningitidis: Biology, microbiology, and epidemiology. Methods Mol. Biol. 2012, 799, 1–20. [Google Scholar]
- AlMazrou, Y.; Khalil, M.; Borrow, R.; Balmer, P.; Bramwell, J.; Lal, G.; Andrews, N.; AlJeffri, M. Serologic responses to ACYW135 polysaccharide meningococcal vaccine in Saudi children under 5 years of age. Infect. Immun. 2005, 73, 2932–2939. [Google Scholar] [CrossRef] [Green Version]
- Masomian, M.; Ahmad, Z.; Gew, L.T.; Poh, C.L. Development of Next Generation Streptococcus pneumoniae Vaccines Conferring Broad Protection. Vaccines 2020, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Nagano, T.; Fraser, P. No-Nonsense Functions for Long Noncoding RNAs. Cell 2011, 145, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Shkumatava, A. LncRNAs in vertebrates: Advances and challenges. Biochimie 2015, 117, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Nojima, T.; Proudfoot, N.J. Mechanisms of lncRNA biogenesis as revealed by nascent transcriptomics. Nat. Rev. Mol. Cell. Biol. 2022, 23, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.B.; Yang, L.; Wang, J.; Hao, Y.Y.; Wang, C.Y.; Lu, Z.M. The Involvement of Long Non-Coding RNAs in Glioma: From Early Detection to Immunotherapy. Front. Immunol. 2022, 13, 897754. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids. Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Ma, J.Y.; Yu, Z.Y.; Li, M.C.; Zhang, W.C.; Sun, H.Q. Epidemiological characteristics and antibiotic resistance mechanisms of Streptococcus pneumoniae: An updated review. Microbiol. Res. 2023, 226, 127221. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.R.; Park, M.A.; Harding, S.; Abraham, R.S. The total IgM, IgA and IgG antibody responses to pneumococcal polysaccharide vaccination (Pneumovax (R) 23) in a healthy adult population and patients diagnosed with primary immunodeficiencies. Vaccine 2019, 37, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Martinez-Hernandez, E.; Arino, H.; Armangue, T.; Spatola, M.; Petit-Pedrol, M.; Saiz, A.; Rosenfeld, M.R.; Graus, F.; Dalmau, J. Clinical and pathogenic significance of IgG, IgA, and IgM antibodies against the NMDA receptor. Neurology 2018, 90, E1386–E1394. [Google Scholar] [CrossRef]
- Singh, K.; Chang, C.; Gershwin, M.E. IgA deficiency and autoimmunity. Autoimmun. Rev. 2013, 13, 163–177. [Google Scholar] [CrossRef]
- Liu, D.; Duan, L.H.; Cyster, J.G. Chemo- and mechanosensing by dendritic cells facilitate antigen surveillance in the spleen. Immunol. Rev. 2022, 306, 25–42. [Google Scholar] [CrossRef]
- Han, S.; Liang, Y.; Li, Y.; Du, W. Long Noncoding RNA Identification: Comparing Machine Learning Based Tools for Long Noncoding Transcripts Discrimination. BioMed Res. Int. 2016, 2016, 8496165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochiai, K.; Igarashi, K. Exploring novel functions of BACH2 in the acquisition of antigen-specific antibodies. Int. Immunol. 2022, 35, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Itoh-Nakadai, A.; Hikota, R.; Muto, A.; Kometani, K.; Watanabe-Matsui, M.; Sato, Y.; Kobayashi, M.; Nakamura, A.; Miura, Y.; Yano, Y.; et al. The transcription repressors Bach2 and Bach1 promote B cell development by repressing the myeloid program. Nat. Immunol. 2014, 15, 1171. [Google Scholar] [CrossRef] [PubMed]
- Itoh-Nakadai, A.; Matsumoto, M.; Kato, H.; Sasaki, J.; Uehara, Y.; Sato, Y.; Ebina-Shibuya, R.; Morooka, M.; Funayama, R.; Nakayama, K.; et al. A Bach2-Cebp Gene Regulatory Network for the Commitment of Multipotent Hematopoietic Progenitors. Cell Rep. 2017, 18, 2401. [Google Scholar] [CrossRef]
- Kato, H.; Itoh Nakadai, A.; Matsumoto, M.; Ishii, Y.; Watanabe Matsui, M.; Ikeda, M.; Ebina Shibuya, R.; Sato, Y.; Kobayashi, M.; Nishizawa, H.; et al. Infection perturbs Bach2 and Bach1 dependent erythroid lineage ‘choice’ to cause anemia. Nat. Immunol. 2018, 19, 1059. [Google Scholar] [CrossRef]
- Roychoudhuri, R.; Clever, D.; Li, P.; Wakabayashi, Y.; Quinn, K.M.; Klebanoff, C.A.; Ji, Y.; Sukumar, M.; Eil, R.L.; Yu, Z.; et al. BACH2 regulates CD8(+) T cell differentiation by controlling access of AP-1 factors to enhancers. Nat. Commun. 2016, 17, 851. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, M.; Ise, W.; Ochi, M.; Suzuki, J.; Kometani, K.; Maruyama, S.; Izumoto, M.; Matsumoto, A.; Takemori, N.; Takemori, A.; et al. Bach2-Batf interactions control Th2-type immune response by regulating the IL-4 amplification loop. Nat. Commun. 2016, 7, 12596. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, K.; Ochiai, K.; Itoh-Nakadai, A.; Muto, A. Orches-tration of plasma cell differentiation by Bach 2 and its gene regulatory network. Immunol. Rev. 2014, 261, 16–25. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, G.; Huang, Y.; Huang, Y.C.; Wei, X.; Shi, J.Y. Polysaccharide conjugate vaccine: A kind of vaccine with great development potential. Chin. Chem Lett. 2021, 32, 331–1340. [Google Scholar] [CrossRef]
- Yamashita, M.; Kuwahara, M. The critical role of Bach 2 in regulating type 2 chronic airway inflammation. Int. Immunol. 2018, 30, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034.1–0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.Y.; Zhan, Y.Q.; Zhao, J.X.; Wang, J.M.; Ma, H.L. Effects of Fluoride on the Expression of p38MAPK Signaling Pathway-Related Genes and Proteins in Spleen Lymphocytes of Mice. Biol Trace Elem. Res. 2016, 173, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Erturk-Hasdemir, D.; Kasper, D.L. Finding a needle in a haystack: Bacteroides fragilis polysaccharide A as the archetypical symbiosis factor. Ann. N. Y. Acad. Sci. 2018, 1417, 116–129. [Google Scholar] [CrossRef]
- Hsieh, S.A.; Allen, P.M. Immunomodulatory roles of polysaccharide capsules in the intestine. Front. Immunol. 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Blandford, L.E.; Johnston, E.L.; Sanderson, J.D.; Wade, W.G.; Lax, A.J. Promoter orientation of the immunomodulatory Bacteroides fragilis capsular polysaccharide A (PSA) is off in individuals with inflammatory bowel disease (IBD). Gut Microbes 2019, 10, 569–577. [Google Scholar] [CrossRef]
- Johnson, J.L.; Jones, M.B.; Cobb, B.A. Polysaccharide-experienced effector T cells induce IL-10 in FoxP3+ regulatory T cells to prevent pulmonary inflammation. Glycobiology 2018, 28, 50–58. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | lncRNA Transcript Name | Pearson Correlation Coefficient |
|---|---|---|
| Zfp384 | ENSMUST00000203287 | 1 |
| Fus | ENSMUST00000128851 | 1 |
| Acy1 | ENSMUST00000187798 | 0.93 |
| Selenop | MSTRG.9430.8 | 0.90 |
| Igf2 | ENSMUST00000136359 | 0.90 |
| Tmem106a | MSTRG.5353.1 | −0.20 |
| Slc30a5 | MSTRG.8116.2 | −0.20 |
| Slc30a5 | MSTRG.8116.4 | −0.20 |
| Ube2d3 | ENSMUST00000181619 | −0.19 |
| GO Term | GO Function | p-Value |
|---|---|---|
| immunoglobulin production in mucosal tissue | biological process | 0.00 |
| negative regulation of CD8-positive, alpha-beta T cell proliferation | biological process | 0.04 |
| positive regulation of T cell activation | biological process | 0.02 |
| negative regulation of interleukin-17 production | biological process | 0.04 |
| negative regulation of CD4-positive, alpha-beta T cell proliferation | biological process | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, N.; Hu, L.; Hu, W.; Li, Q.; Mao, H.; Wang, M.; Ke, Z.; Qi, L.; Wang, J. Comparative Transcriptome Profiling of mRNA and lncRNA of Mouse Spleens Inoculated with the Group ACYW135 Meningococcal Polysaccharide Vaccine. Vaccines 2023, 11, 1295. https://doi.org/10.3390/vaccines11081295
Zhu N, Hu L, Hu W, Li Q, Mao H, Wang M, Ke Z, Qi L, Wang J. Comparative Transcriptome Profiling of mRNA and lncRNA of Mouse Spleens Inoculated with the Group ACYW135 Meningococcal Polysaccharide Vaccine. Vaccines. 2023; 11(8):1295. https://doi.org/10.3390/vaccines11081295
Chicago/Turabian StyleZhu, Nan, Liping Hu, Wenlong Hu, Qiang Li, Haiguang Mao, Mengting Wang, Zhijian Ke, Lili Qi, and Jinbo Wang. 2023. "Comparative Transcriptome Profiling of mRNA and lncRNA of Mouse Spleens Inoculated with the Group ACYW135 Meningococcal Polysaccharide Vaccine" Vaccines 11, no. 8: 1295. https://doi.org/10.3390/vaccines11081295
APA StyleZhu, N., Hu, L., Hu, W., Li, Q., Mao, H., Wang, M., Ke, Z., Qi, L., & Wang, J. (2023). Comparative Transcriptome Profiling of mRNA and lncRNA of Mouse Spleens Inoculated with the Group ACYW135 Meningococcal Polysaccharide Vaccine. Vaccines, 11(8), 1295. https://doi.org/10.3390/vaccines11081295