Nora Virus VP4b and ORF1 Circulate in Hemolymph of Infected D. melanogaster with Coordinate Expression of Vago and Vir-1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Husbandry and Time Course
2.2. Hemolymph Sampling
2.3. Preparation of Mouse Antisera against Recombinant VP4b, Vago, and Vir-1 Proteins
2.4. Protein Extraction and Western Blot Analysis
2.5. RNA Extraction, RT-PCR, and qRT-PCR Analyses of Nora Virus, Vago, and Vir-1
3. Results
3.1. Validation of Nora Virus and DCV Infection Using RT-PCR
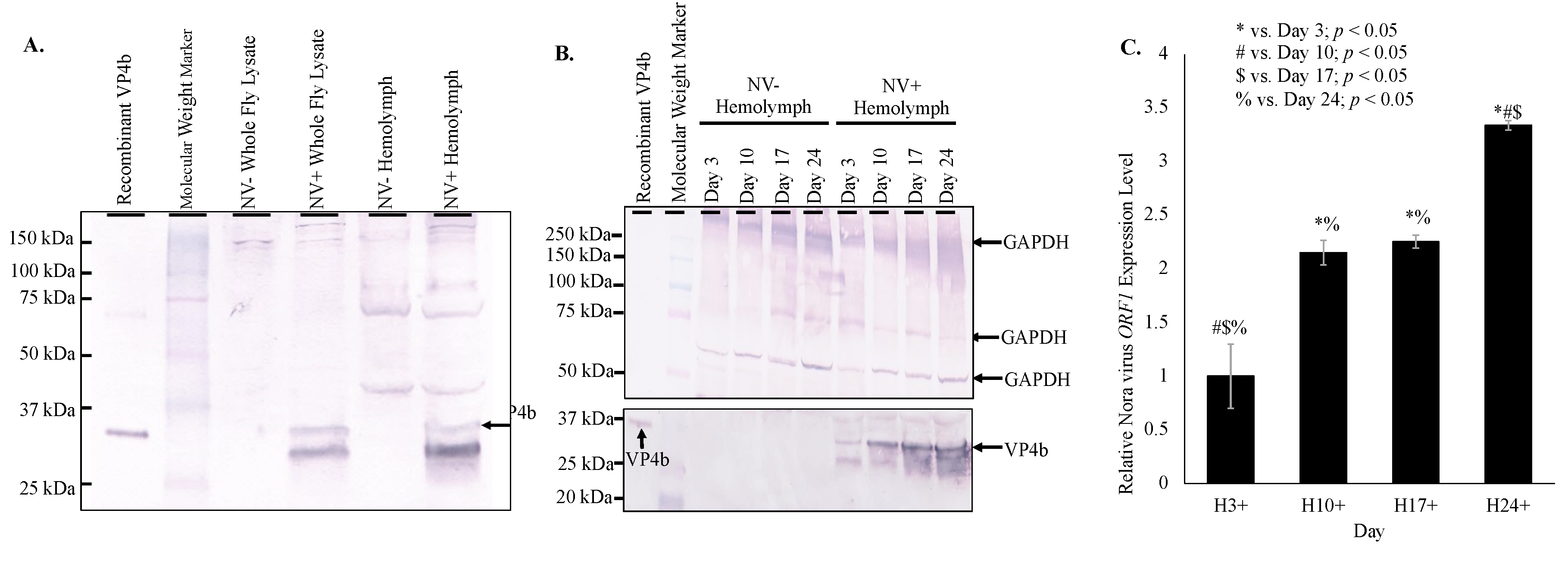
3.2. Nora Virus is Present in the Hemolymph of Infected D. melanogaster
3.3. Viral Load Follows a Biphasic Pattern
3.4. Vago Expression Is Upregulated in Nora Virus-Infected D. melanogaster
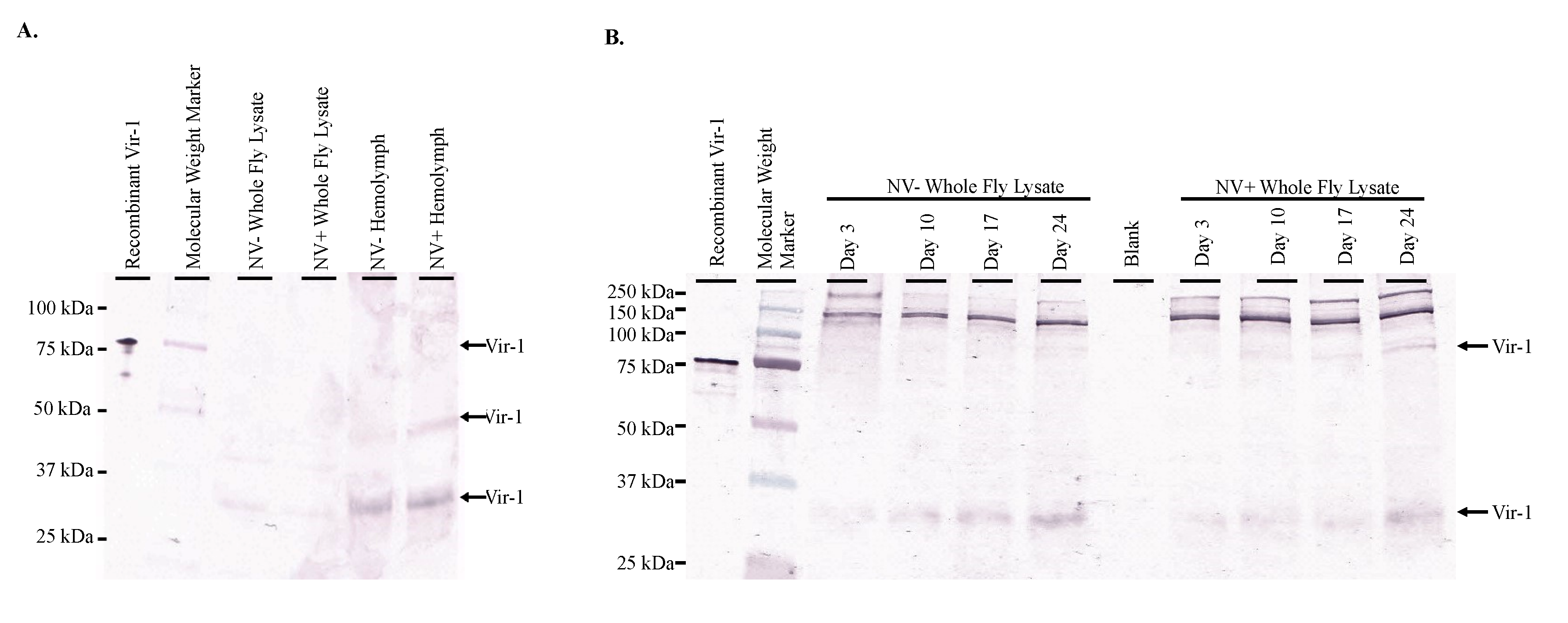
3.5. Vir-1 Presence Does Not Require Viral Infection
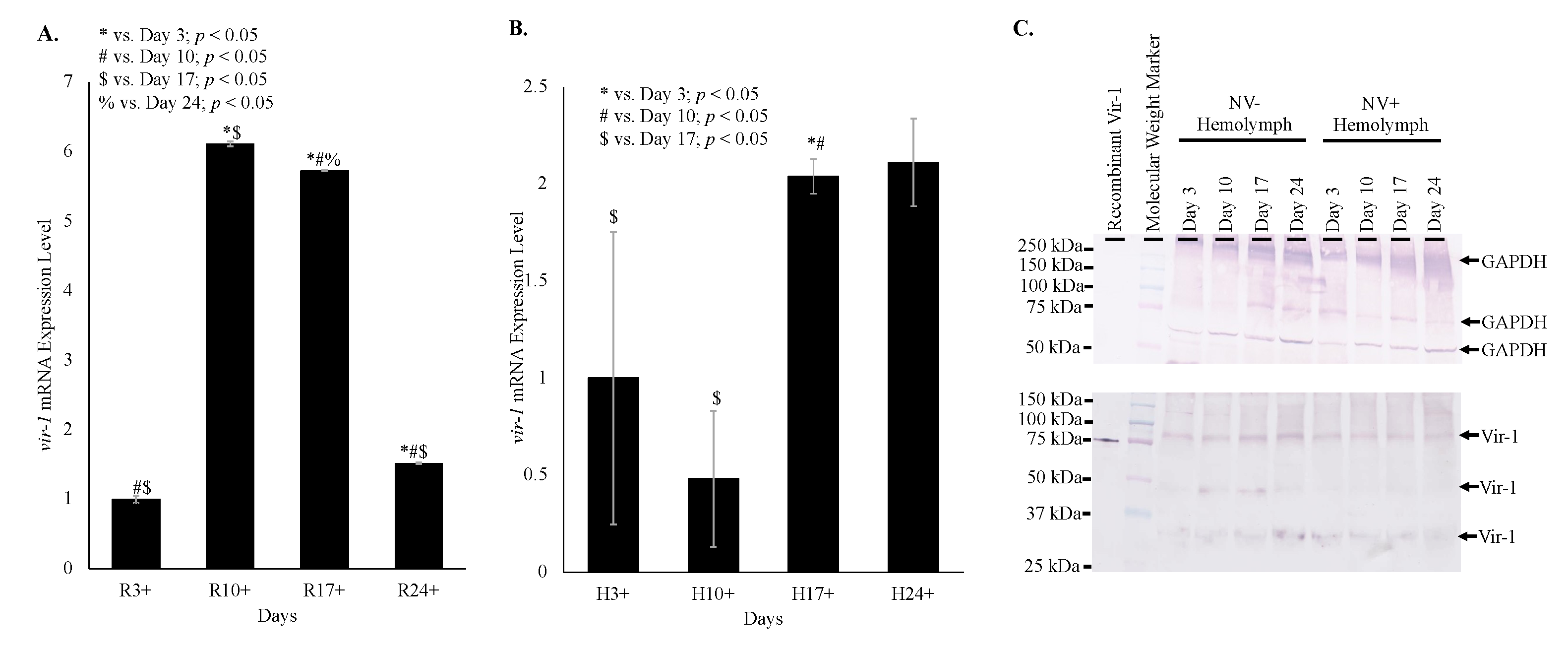
3.6. Vir-1 Expression Is Increased in Nora Virus-Infected D. melanogaster
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hergannan, J.; Rechart, J.-V. Drosophila immunity. Trends Cell. Biol. 1997, 7, 309–316. [Google Scholar] [CrossRef]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome. Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valanne, S.; Wang, J.-H.; Rämet, M. The Drosophila Toll signaling pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Huffmann, J.A.; Imler, J.-L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- McPhee, C.K.; Baehrecke, E.H. Autophagy in Drosophila melanogaster. Biochim. Biophys. Acta Mol. Cell. Res. 2009, 1793, 1452–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, C.; Mueller, S.; Goto, A.; Barbier, V.; Paro, S.; Bonnay, F.; Dostert, C.; Troxler, L.; Hetru, C.; Meignin, C.; et al. Broad RNA interference-mediated antiviral immunity and virus-specific inducible responses in Drosophila. J. Immunol. 2013, 190, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Paradkar, P.N.; Duchemin, J.B.; Voysey, R.; Walker, P.J. Dicer-2-dependent activation of Culex Vago occurs via the TRAF-Rel2 signaling pathway. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antiviral Immunity in the Fruit Fly, Drosophila melanogaster. Available online: https://www.intechopen.com/books/drosophila-melanogaster-model-for-recent-advances-in-genetics-and-therapeutics/antiviral-immunity-in-the-fruit-fly-drosophila-melanogaster (accessed on 26 August 2020).
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffman, J.A.; Imler, J.-L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in Drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Correspondence, K.N.J.; Johnson, K.N. Induction of host defence responses by Drosophila C virus. J. Gen. Virol. 2008, 89, 1497–1501. [Google Scholar] [CrossRef]
- Salehipour-shirazi, G.; Ferguson, L.V.; Sinclair, B.J. Does cold activate the Drosophila melanogaster immune system? J. Insect. Physiol. 2017, 96, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Habayeb, M.S.; Ekengren, S.K.; Hultmark, D. Nora virus, a persistent virus in Drosophila, defines a new picorna-like virus family. J. Gen. Virol. 2006, 87, 3045–3051. [Google Scholar] [CrossRef] [PubMed]
- Habayeb, M.S.; Cantera, R.; Casanova, G.; Ekström, J.O.; Albright, S.; Hultmark, D. The Drosophila Nora virus is an enteric virus, transmitted via feces. J. Invertebr. Pathol. 2009, 101, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Ekstrom, J.-O.; Habayeb, M.S.; Srivastava, V.; Kieselbach, T.; Wingsle, G.; Hultmark, D. Drosophila Nora virus capsid proteins differe from those of other picorna-like viruses. Virus Res. 2011, 160, 51–58. [Google Scholar] [CrossRef]
- Rogers, A.; Towery, L.; McCown, A.; Carlson, K.A. Impaired geotaxis as a novel phenotype of Nora virus infection of Drosophila melanogaster. Scientifica 2020, in press. [Google Scholar] [CrossRef]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating immune cells mediate a systemic RNAi-based adaptive antiviral response in Drosophila. Cell 2017, 169, 314–325.e13. [Google Scholar] [CrossRef] [Green Version]
- Cordes, E.J.; Licking-Murray, K.D.; Carlson, K.A. Differential gene expression related to Nora virus infection of Drosophila melanogaster. Virus Res. 2013, 175, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, W.; Page, A.M.; Carlson, D.J.; Ericson, B.L.; Cserhati, M.F.; Guda, C.; Carlson, K.A. Analysis of immune-related genes during Nora virus infection of Drosophila melanogaster using next generation sequencing. AIMS Microbiol. 2018, 4, 123–139. [Google Scholar] [CrossRef]
- Ericson, B.L.; Carlson, D.J.; Carlson, K.A. Characterization of Nora virus structural proteins via Western blot analysis. Scientifica 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vlisidouj, I.; Wood, W. Drosophila blood cells and their role in immune responses. FEBS J. 2015, 282, 1368–1382. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed]
- Granados, R.R.; Lawler, K.A. In vivo pathway of Autographa californica baculovirus invasion and infection. Virology 1981, 108, 297–308. [Google Scholar] [CrossRef]
- Fujiyuki, T.; Takeuchi, H.; Ono, M.; Ohka, S.; Sasaki, T.; Nomoto, A.; Kubo, T. Novel insect picorna-like virus identified in the brains of aggressive worker honeybees. J. Virol. 2004, 78, 1093–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reetoo, K.N.; Osman, S.A.; Illavia, S.J.; Cameron-Wilson, C.L.; Bantavala, J.E.; Muir, P. Quantitative analysis of viral RNA kinetics in coxsackievirus B3-induced murine myocarditis: Biphasic pattern of clearance following acute infection, with persistence of residual viral RNA throughout and beyond the inflammatory phase of disease. J. Gen. Virol. 2000, 81, 2755–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mierlo, J.T.; Overheul, G.J.; Obadia, B.; Van Cleef, K.W.R.; Webster, C.L. Novel Drosophila viruses encode host-specific suppressors of RNAi. PLoS Pathog. 2014, 10, 1004256. [Google Scholar] [CrossRef] [Green Version]
- Habayeb, M.S.; Ekström, J.O.; Hultmark, D. Nora virus persistent infections are not affected by the RNAi machinery. PLoS ONE 2009, 4, 4–8. [Google Scholar] [CrossRef]
- Agaisse, H.; Petersen, U.M.; Boutros, M.; Mathey-Prevot, B.; Perrimon, N. Signaling role of hemocytes in Drosophila JAK/STAT-dependent response to septic injury. Dev. Cell 2003, 5, 441–450. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macke, A.; Lopez, W.; Carlson, D.J.; Carlson, K.A. Nora Virus VP4b and ORF1 Circulate in Hemolymph of Infected D. melanogaster with Coordinate Expression of Vago and Vir-1. Vaccines 2020, 8, 491. https://doi.org/10.3390/vaccines8030491
Macke A, Lopez W, Carlson DJ, Carlson KA. Nora Virus VP4b and ORF1 Circulate in Hemolymph of Infected D. melanogaster with Coordinate Expression of Vago and Vir-1. Vaccines. 2020; 8(3):491. https://doi.org/10.3390/vaccines8030491
Chicago/Turabian StyleMacke, Amanda, Wilfredo Lopez, Darby J. Carlson, and Kimberly A. Carlson. 2020. "Nora Virus VP4b and ORF1 Circulate in Hemolymph of Infected D. melanogaster with Coordinate Expression of Vago and Vir-1" Vaccines 8, no. 3: 491. https://doi.org/10.3390/vaccines8030491
APA StyleMacke, A., Lopez, W., Carlson, D. J., & Carlson, K. A. (2020). Nora Virus VP4b and ORF1 Circulate in Hemolymph of Infected D. melanogaster with Coordinate Expression of Vago and Vir-1. Vaccines, 8(3), 491. https://doi.org/10.3390/vaccines8030491