Anti-Flavivirus Vaccines: Review of the Present Situation and Perspectives of Subunit Vaccines Produced in Escherichia coli

 ,
, 
 , and
, and 
Abstract
:1. Introduction
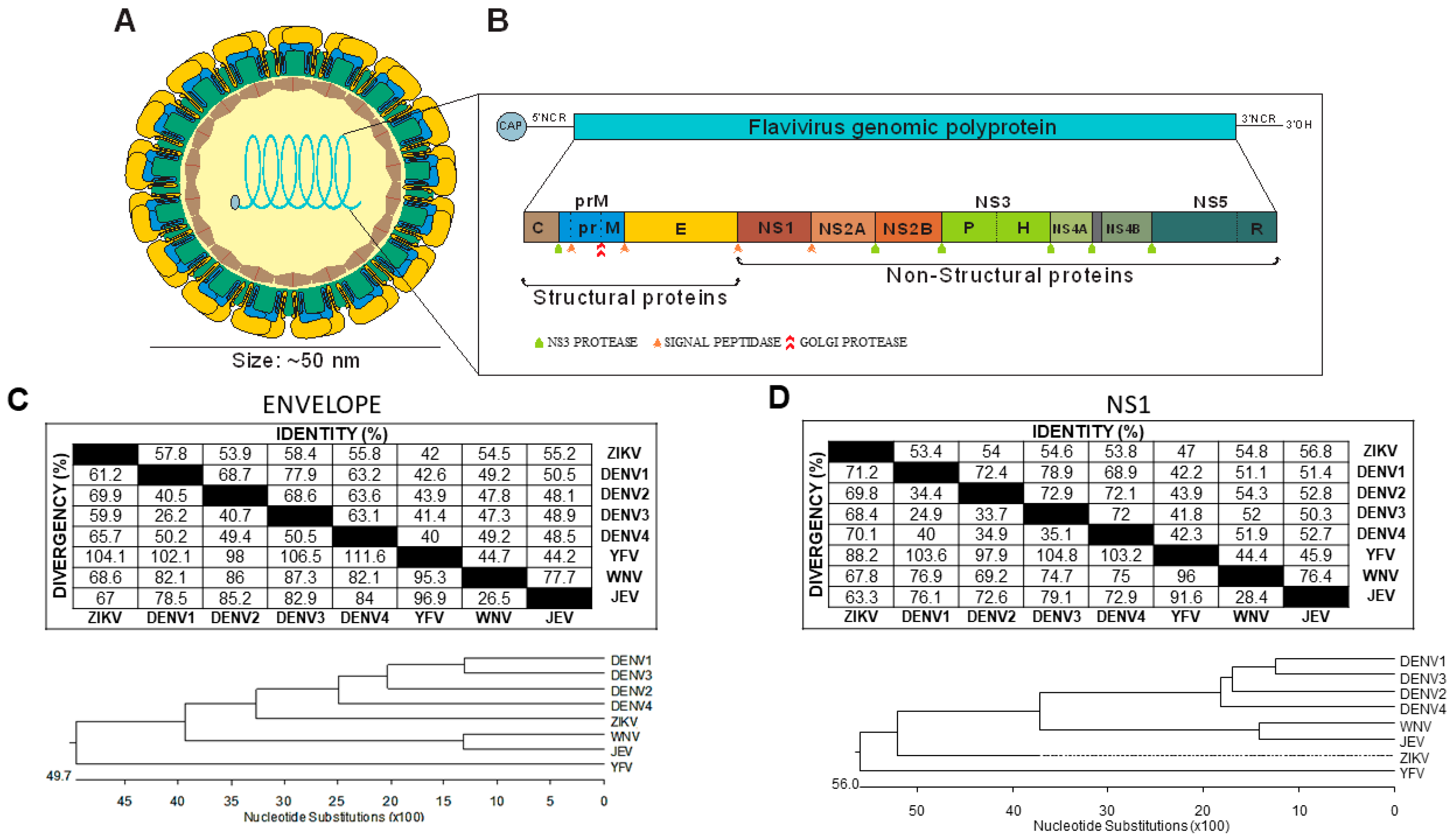
2. Flavivirus Structure and Proteins
3. Yellow Fever Virus
4. Japanese Encephalitis Virus
5. West Nile Virus
6. Dengue Virus
7. Zika Virus
8. Discussion
Funding
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses (ICTV) Virus Taxonomy: 2019 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 21 July 2020).
- Kuhn, R.J.; Zhang, W.; Rossmann, M.G.; Pletnev, S.V.; Corver, J.; Lenches, E.; Jones, C.T.; Mukhopadhyay, S.; Chipman, P.R.; Strauss, E.G.; et al. Structure of dengue virus: Implications for flavivirus organization, maturation, and fusion. Cell 2002, 108, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ge, P.; Yu, X.; Brannan, J.M.; Bi, G.; Zhang, Q.; Schein, S.; Hong Zhou, Z. Cryo-EM structure of the mature dengue virus at 3.5-Å resolution. Nat. Struct. Mol. Biol. 2013, 20, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S. Structure of West Nile Virus. Science 2003, 302, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.D.; Whitehead, S.S.; Pierce, K.K.; Tibery, C.M.; Grier, P.L.; Hynes, N.A.; Larsson, C.J.; Sabundayo, B.P.; Talaat, K.R.; Janiak, A.; et al. The live attenuated dengue vaccine TV003 elicits complete protection against dengue in a human challenge model. Sci. Transl. Med. 2016, 8, 330ra36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, S.H.; Zhu, L.; Nian, Q.G.; Yuan, S.; Gao, Q.; Hu, Z.; Ye, Q.; Li, X.F.; Xie, D.Y.; et al. Near-atomic structure of Japanese encephalitis virus reveals critical determinants of virulence and stability. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Morrone, S.R.; Chew, V.S.Y.; Lim, X.N.; Ng, T.S.; Kostyuchenko, V.A.; Zhang, S.; Wirawan, M.; Chew, P.L.; Lee, J.; Tan, J.L.; et al. High flavivirus structural plasticity demonstrated by a non-spherical morphological variant. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, L. Systematic Analysis of Structure Similarity between Zika Virus and Other Flaviviruses. ACS Infect. Dis. 2019, 5, 1070–1080. [Google Scholar] [CrossRef]
- Watterson, D.; Modhiran, N.; Young, P.R. The many faces of the flavivirus NS1 protein offer a multitude of options for inhibitor design. Antiviral Res. 2016, 130, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Melian, E.B.; Hinzman, E.; Nagasaki, T.; Firth, A.E.; Wills, N.M.; Nouwens, A.S.; Blitvich, B.J.; Leung, J.; Funk, A.; Atkins, J.F.; et al. NS1′ of Flaviviruses in the Japanese Encephalitis Virus Serogroup Is a Product of Ribosomal Frameshifting and Plays a Role in Viral Neuroinvasiveness. J. Virol. 2010, 84, 1641–1647. [Google Scholar] [CrossRef] [Green Version]
- Chambers, T.J.; McCourt, D.W.; Rice, C.M. Yellow fever virus proteins NS2A, NS213, and NS4B: Identification and partial N-terminal amino acid sequence analysis. Virology 1989, 169, 100–109. [Google Scholar] [CrossRef]
- Mackenzie, J.M.; Khromykh, A.A.; Jones, M.K.; Westaway, E.G. Subcellular Localization and Some Biochemical Properties of the Flavivirus Kunjin Nonstructural Proteins NS2A and NS4A. Virology 1998, 245, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chen, H.B.; Wang, X.J.; Huang, H.; Khromykh, A.A. Analysis of Adaptive Mutations in Kunjin Virus Replicon RNA Reveals a Novel Role for the Flavivirus Nonstructural Protein NS2A in Inhibition of Beta Interferon Promoter-Driven Transcription. J. Virol. 2004, 78, 12225–12235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.J.; Wang, X.J.; Mokhonov, V.V.; Shi, P.-Y.; Randall, R.; Khromykh, A.A. Inhibition of Interferon Signaling by the New York 99 Strain and Kunjin Subtype of West Nile Virus Involves Blockage of STAT1 and STAT2 Activation by Nonstructural Proteins. J. Virol. 2005, 79, 1934–1942. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Wang, X.J.; Clark, D.C.; Lobigs, M.; Hall, R.A.; Khromykh, A.A. A Single Amino Acid Substitution in the West Nile Virus Nonstructural Protein NS2A Disables Its Ability To Inhibit Alpha/Beta Interferon Induction and Attenuates Virus Virulence in Mice. J. Virol. 2006, 80, 2396–2404. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Jordan, J.L.; Sanchez-Burgos, G.G.; Laurent-Rolle, M.; Garcia-Sastre, A. Inhibition of interferon signaling by dengue virus. Proc. Natl. Acad. Sci. USA 2003, 100, 14333–14338. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.Y.; Pijlman, G.P.; Kondratieva, N.; Hyde, J.; Mackenzie, J.M.; Khromykh, A.A. Role of Nonstructural Protein NS2A in Flavivirus Assembly. J. Virol. 2008, 82, 4731–4741. [Google Scholar] [CrossRef] [Green Version]
- Kümmerer, B.M.; Rice, C.M. Mutations in the Yellow Fever Virus Nonstructural Protein NS2A Selectively Block Production of Infectious Particles. J. Virol. 2002, 76, 4773–4784. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Chen, H.B.; Khromykh, A.A. Molecular and Functional Analyses of Kunjin Virus Infectious cDNA Clones Demonstrate the Essential Roles for NS2A in Virus Assembly and for a Nonconservative Residue in NS3 in RNA Replication. J. Virol. 2003, 77, 7804–7813. [Google Scholar] [CrossRef] [Green Version]
- Falgout, B.; Pethel, M.; Zhang, Y.M.; Lai, C.J. Both nonstructural proteins NS2B and NS3 are required for the proteolytic processing of dengue virus nonstructural proteins. J. Virol. 1991, 65, 2467–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollati, M.; Alvarez, K.; Assenberg, R.; Baronti, C.; Canard, B.; Cook, S.; Coutard, B.; Decroly, E.; de Lamballerie, X.; Gould, E.A.; et al. Structure and functionality in flavivirus NS-proteins: Perspectives for drug design. Antiviral Res. 2010, 87, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, Y.; Loh, Y.R.; Phoo, W.W.; Hung, A.W.; Kang, C.; Luo, D. Crystal structure of unlinked NS2B-NS3 protease from Zika virus. Science 2016, 354, 1597–1600. [Google Scholar] [CrossRef] [PubMed]
- Roosendaal, J.; Westaway, E.G.; Khromykh, A.; Mackenzie, J.M. Regulated Cleavages at the West Nile Virus NS4A-2K-NS4B Junctions Play a Major Role in Rearranging Cytoplasmic Membranes and Golgi Trafficking of the NS4A Protein. J. Virol. 2006, 80, 4623–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, C.S.H.; Chu, J.J.H. Cellular Vimentin Regulates Construction of Dengue Virus Replication Complexes through Interaction with NS4A Protein. J. Virol. 2014, 88, 1897–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, J.E.; Wudzinska, A.; Datan, E.; Quaglino, D.; Zakeri, Z. Flavivirus NS4A-induced Autophagy Protects Cells against Death and Enhances Virus Replication. J. Biol. Chem. 2011, 286, 22147–22159. [Google Scholar] [CrossRef] [Green Version]
- Shiryaev, S.A.; Chernov, A.V.; Aleshin, A.E.; Shiryaeva, T.N.; Strongin, A.Y. NS4A regulates the ATPase activity of the NS3 helicase: A novel cofactor role of the non-structural protein NS4A from West Nile virus. J. Gen. Virol. 2009, 90, 2081–2085. [Google Scholar] [CrossRef]
- Umareddy, I. Dengue virus NS4B interacts with NS3 and dissociates it from single-stranded RNA. J. Gen. Virol. 2006, 87, 2605–2614. [Google Scholar] [CrossRef]
- Ambrose, R.L.; Mackenzie, J.M. West Nile Virus Differentially Modulates the Unfolded Protein Response To Facilitate Replication and Immune Evasion. J. Virol. 2011, 85, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Jordán, J.L.; Laurent-Rolle, M.; Ashour, J.; Martínez-Sobrido, L.; Ashok, M.; Lipkin, W.I.; García-Sastre, A. Inhibition of Alpha/Beta Interferon Signaling by the NS4B Protein of Flaviviruses. J. Virol. 2005, 79, 8004–8013. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Yi, G.; Du, F.; Chuang, Y.-C.; Vaughan, R.C.; Sankaran, B.; Kao, C.C.; Li, P. Structure and function of the Zika virus full-length NS5 protein. Nat. Commun. 2017, 8, 14762. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Tan, X.-F.; Thurmond, S.; Zhang, Z.-M.; Lin, A.; Hai, R.; Song, J. The structure of Zika virus NS5 reveals a conserved domain conformation. Nat. Commun. 2017, 8, 14763. [Google Scholar] [CrossRef]
- Upadhyay, A.K.; Cyr, M.; Longenecker, K.; Tripathi, R.; Sun, C.; Kempf, D.J. Crystal structure of full-length Zika virus NS5 protein reveals a conformation similar to Japanese encephalitis virus NS5. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-H.; Yip, B.-S.; Huang, L.-M.; Wu, S.-C. Zika virus structural biology and progress in vaccine development. Biotechnol. Adv. 2018, 36, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Volk, D.E.; May, F.J.; Gandham, S.H.A.; Anderson, A.; Von Lindern, J.J.; Beasley, D.W.C.; Barrett, A.D.T.; Gorenstein, D.G. Structure of yellow fever virus envelope protein domain III. Virology 2009, 394, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cane, P.A.; Gould, E.A. Reduction of Yellow Fever Virus Mouse Neurovirulence by Immunization with a Bacterially Synthesized Non-structural Protein (NS1) Fragment. J. Gen. Virol. 1988, 69, 1241–1246. [Google Scholar] [CrossRef]
- Lin, C.W.; Liu, K.T.; Huang, H.D.; Chen, W.J. Protective immunity of E. coli-synthesized NS1 protein of Japanese encephalitis virus. Biotechnol. Lett. 2008, 30, 205–214. [Google Scholar] [CrossRef]
- Amorim, J.H.; Diniz, M.O.; Cariri, F.A.M.O.; Rodrigues, J.F.; Bizerra, R.S.P.; Gonçalves, A.J.S.; de Barcelos Alves, A.M.; de Souza Ferreira, L.C. Protective immunity to DENV2 after immunization with a recombinant NS1 protein using a genetically detoxified heat-labile toxin as an adjuvant. Vaccine 2012, 30, 837–845. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.P.d.S.; Pereira, L.R.; Fabris, D.L.N.; Salvador, F.S.; Santos, R.A.; Zanotto, P.M.d.A.; Romano, C.M.; Amorim, J.H.; Ferreira, L.C.d.S. Production of a Recombinant Dengue Virus 2 NS5 Protein and Potential Use as a Vaccine Antigen. Clin. Vaccine Immunol. 2016, 23, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, R.; Falcón, R.; Izquierdo, A.; García, A.; Alvarez, M.; Pérez, A.B.; Soto, Y.; Muné, M.; da Silva, E.M.; Ortega, O.; et al. Recombinant dengue 2 virus NS3 protein conserves structural antigenic and immunological properties relevant for dengue vaccine design. Virus Genes 2014, 49, 185–195. [Google Scholar] [CrossRef]
- Kao, Y.-S.; Yu, C.-Y.; Huang, H.-J.; Tien, S.-M.; Wang, W.-Y.; Yang, M.; Anderson, R.; Yeh, T.-M.; Lin, Y.-S.; Wan, S.-W. Combination of Modified NS1 and NS3 as a Novel Vaccine Strategy against Dengue Virus Infection. J. Immunol. 2019, 203, 1909–1917. [Google Scholar] [CrossRef]
- Costa, S.M.; Yorio, A.P.; Gonçalves, A.J.S.; Vidale, M.M.; Costa, E.C.B.; Mohana-Borges, R.; Motta, M.A.; Freire, M.S.; Alves, A.M.B. Induction of a Protective Response in Mice by the Dengue Virus NS3 Protein Using DNA Vaccines. PLoS ONE 2011, 6, e25685. [Google Scholar] [CrossRef] [Green Version]
- Monath, T.P. Yellow fever: An update. Lancet Infect. Dis. 2001, 1, 11–20. [Google Scholar] [CrossRef]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef]
- Silva, N.I.O.; Sacchetto, L.; De Rezende, I.M.; Trindade, G.D.S.; Labeaud, A.D.; De Thoisy, B.; Drumond, B.P. Recent sylvatic yellow fever virus transmission in Brazil: The news from an old disease. Virol. J. 2020, 17, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; van der Most, R.G.; Akondy, R.S.; Glidewell, J.T.; Albott, S.; Masopust, D.; Murali-Krishna, K.; Mahar, P.L.; Edupuganti, S.; Lalor, S.; et al. Human Effector and Memory CD8+ T Cell Responses to Smallpox and Yellow Fever Vaccines. Immunity 2008, 28, 710–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministério da Saúde do Brasil Monitoramento dos casos e óbitos de Febre Amarela no Brasil–2017. Available online: http://portalarquivos.saude.gov.br/images/pdf/2017/junho/02/COES-FEBRE-AMARELA---INFORME-43---Atualiza----o-em-31maio2017.pdf (accessed on 21 July 2020).
- Ministério da Saúde do Brasil Monitoramento do Período Sazonal da Febre Amarela Brasil-2017/2018. Available online: http://portalarquivos2.saude.gov.br/images/pdf/2018/outubro/08/Informe-FA.pdf (accessed on 21 July 2020).
- Ministério da Saúde do Brasil Monitoramente de Febre Amarela Brasil 2019. Available online: https://www.saude.gov.br/images/pdf/2019/junho/13/Informe-de-Monitoramento-de-Febre-Amarela-Brasil--n-18.pdf (accessed on 21 July 2020).
- Ferguson, M.; Shin Jinho, J.; Knezevic, I.; Minor, P.; Barrett, A. WHO Working Group on Technical Specifications for Manufacture and Evaluation of Yellow Fever Vaccines. In Proceedings of the Vaccine, Geneva, Switzerland, 13–14 May 2009; Elsevier: Amsterdam, The Netherlands, 2010; Volume 28, pp. 8236–8245. [Google Scholar]
- Barrett, A.D.T. Yellow Fever Vaccine: The Conundrum of 2 Doses, One Dose, or One-Fifth Dose to Induce and Maintain Protective Immunity. J. Infect. Dis. 2020, 221, 1922–1924. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, N.P.; Schroeder, B.A.; Miller, E.R.; Braun, M.M.; Hinckley, A.F.; Marano, N.; Slade, B.A.; Barnett, E.D.; Brunette, G.W.; Horan, K.; et al. Adverse event reports following yellow fever vaccination. Vaccine 2008, 26, 6077–6082. [Google Scholar] [CrossRef] [Green Version]
- Vratskikh, O.; Stiasny, K.; Zlatkovic, J.; Tsouchnikas, G.; Jarmer, J.; Karrer, U.; Roggendorf, M.; Roggendorf, H.; Allwinn, R.; Heinz, F.X. Dissection of Antibody Specificities Induced by Yellow Fever Vaccination. PLoS Pathog. 2013, 9, e1003458. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; Nichols, R.; Archambault, W.T.; Moore, L.; Marchesani, R.; Tian, J.; Shope, R.E.; Thomas, N.; Schrader, R.; Furby, D.; et al. Comparative safety and immunogenicity of two yellow fever 17D vaccines (ARILVAX and YF-VAX) in a phase III multicenter, double-blind clinical trial. Am. J. Trop. Med. Hyg. 2002, 66, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Kohler, S.; Bethke, N.; Böthe, M.; Sommerick, S.; Frentsch, M.; Romagnani, C.; Niedrig, M.; Thiel, A. The early cellular signatures of protective immunity induced by live viral vaccination. Eur. J. Immunol. 2012, 42, 2363–2373. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, D.; Therrien, R.; Kettaf, N.; Angermann, B.R.; Boucher, G.; Filali-Mouhim, A.; Moser, J.M.; Mehta, R.S.; Drake, D.R.; Castro, E.; et al. Yellow fever vaccine induces integrated multilineage and polyfunctional immune responses. J. Exp. Med. 2008, 205, 3119–3131. [Google Scholar] [CrossRef]
- Douam, F.; Ploss, A. Yellow Fever Virus: Knowledge Gaps Impeding the Fight Against an Old Foe. Trends Microbiol. 2018, 26, 913–928. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; Guirakhoo, F.; Nichols, R.; Yoksan, S.; Schrader, R.; Murphy, C.; Blum, P.; Woodward, S.; McCarthy, K.; Mathis, D.; et al. Chimeric Live, Attenuated Vaccine against Japanese Encephalitis (ChimeriVax-JE): Phase 2 Clinical Trials for Safety and Immunogenicity, Effect of Vaccine Dose and Schedule, and Memory Response to Challenge with Inactivated Japanese Encephalitis Antigen. J. Infect. Dis. 2003, 188, 1213–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, B.; Nougarede, N.; Begue, S.; Sanchez, V.; Souag, N.; Carre, M.; Chambonneau, L.; Morrisson, D.N.; Shaw, D.; Qiao, M.; et al. Cell-mediated immunity induced by chimeric tetravalent dengue vaccine in naive or flavivirus-primed subjects. Vaccine 2008, 26, 5712–5721. [Google Scholar] [CrossRef]
- Monath, T.P.; Liu, J.; Kanesa-Thasan, N.; Myers, G.A.; Nichols, R.; Deary, A.; McCarthy, K.; Johnson, C.; Ermak, T.; Shin, S.; et al. A live, attenuated recombinant West Nile virus vaccine. Proc. Natl. Acad. Sci. USA 2006, 103, 6694–6699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giel-Moloney, M.; Goncalvez, A.P.; Catalan, J.; Lecouturier, V.; Girerd-Chambaz, Y.; Diaz, F.; Maldonado-Arocho, F.; Gomila, R.C.; Bernard, M.C.; Oomen, R.; et al. Chimeric yellow fever 17D-Zika virus (ChimeriVax-Zika) as a live-attenuated Zika virus vaccine. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, M.C.; Garratt, R.C.; Caufour, P.S.; Freire, M.S.; Rodrigues, M.M.; Nussenzweig, R.S.; Galler, R. Surface expression of an immunodominant malaria protein B cell epitope by yellow fever virus. J. Mol. Biol. 2002, 315, 873–885. [Google Scholar] [CrossRef]
- Bonaldo, M.C.; Martins, M.A.; Rudersdorf, R.; Mudd, P.A.; Sacha, J.B.; Piaskowski, S.M.; Costa Neves, P.C.; Veloso de Santana, M.G.; Vojnov, L.; Capuano, S.; et al. Recombinant Yellow Fever Vaccine Virus 17D Expressing Simian Immunodeficiency Virus SIVmac239 Gag Induces SIV-Specific CD8+ T-Cell Responses in Rhesus Macaques. J. Virol. 2010, 84, 3699–3706. [Google Scholar] [CrossRef] [Green Version]
- Khromava, A.Y.; Eidex, R.B.; Weld, L.H.; Kohl, K.S.; Bradshaw, R.D.; Chen, R.T.; Cetron, M.S. Yellow fever vaccine: An updated assessment of advanced age as a risk factor for serious adverse events. Vaccine 2005, 23, 3256–3263. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization The Weekly Epidemiological Record (WER). Weely Epidemiol. Rec. 2018, 93, 445–452.
- Porudominsky, R.; Gotuzzo, E.H. Yellow fever vaccine and risk of developing serious adverse events: A systematic review. Rev. Panam. Salud Publica/Pan Am. J. Public Health 2018, 42, 18. [Google Scholar] [CrossRef] [Green Version]
- Marfin, A.A.; Eidex, R.S.B.; Kozarsky, P.E.; Cetron, M.S. Yellow fever and Japanese encephalitis vaccines: Indications and complications. Infect. Dis. Clin. North Am. 2005, 19, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, S.D.A.; Nunes-Araújo, F.R.F.; Pires, W.P.; Silva, F.A.; Costa, H.L. Yellow fever vaccination during pregnancy and spontaneous abortion: A case-control study. Trop. Med. Int. Health 1998, 3, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, K.; Ewan, P.W.; Nasser, S.M. Administration of yellow fever vaccine in patients with egg allergy. Int. Arch. Allergy Immunol. 2013, 161, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Zidane, N.; Dussart, P.; Bremand, L.; Villani, M.E.; Bedouelle, H. Thermodynamic stability of domain III from the envelope protein of flaviviruses and its improvement by molecular design. Protein Eng. Des. Sel. 2013, 26, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.T.; Ma, L.; Burgner, J.W.; Groesch, T.D.; Post, C.B.; Kuhn, R.J. Flavivirus Capsid Is a Dimeric Alpha-Helical Protein. J. Virol. 2003, 77, 7143–7149. [Google Scholar] [CrossRef] [Green Version]
- Warrener, P.; Tamura, J.K.; Collett, M.S. RNA-stimulated NTPase activity associated with yellow fever virus NS3 protein expressed in bacteria. J. Virol. 1993, 67, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Lohr, K.; Knox, J.E.; Phong, W.Y.; Ma, N.L.; Yin, Z.; Sampath, A.; Patel, S.J.; Wang, W.-L.; Chan, W.-L.; Rao, K.R.R.; et al. Yellow fever virus NS3 protease: Peptide-inhibition studies. J. Gen. Virol. 2007, 88, 2223–2227. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Hoover, S.; Falk, S.P.; Weisblum, B.; Vestling, M.; Striker, R. Phosphorylation of yellow fever virus NS5 alters methyltransferase activity. Virology 2008, 380, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Mayuri, S.M.B.; Perera, R.; Kuhn, R.J.; Striker, R. Protein Kinase G Phosphorylates Mosquito-Borne Flavivirus NS5. J. Virol. 2009, 83, 9195–9205. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Kim, W.-B.; Cho, S.-Y.; Oh, E.-J.; Lee, H.; Kang, K.; Lee, Y.; Lee, D.-G. A Simple Method for the Design and Development of Flavivirus NS1 Recombinant Proteins Using an In Silico Approach. Biomed Res. Int. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Despres, P.; Girard, M.; Bouloy, M. Characterization of yellow fever virus proteins E and NS1 expressed in Vero and Spodoptera frugiperda cells. J. Gen. Virol. 1991, 72, 1331–1342. [Google Scholar] [CrossRef] [PubMed]
- Tottey, S.; Shoji, Y.; Jones, R.M.; Chichester, J.A.; Green, B.J.; Musiychuk, K.; Si, H.; Manceva, S.D.; Rhee, A.; Shamloul, M.; et al. Plant-produced subunit vaccine candidates against yellow fever induce virus neutralizing antibodies and confer protection against viral challenge in animal models. Am. J. Trop. Med. Hyg. 2018, 98, 420–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosta, S.F.D.O.; Passos, M.S.; Kato, R.; Salgado, Á.; Xavier, J.; Jaiswal, A.K.; Soares, S.C.; Azevedo, V.; Giovanetti, M.; Tiwari, S.; et al. Multi-epitope based vaccine against yellow fever virus applying immunoinformatics approaches. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P. Japanese Encephalitis—A Plague of the Orient. N. Engl. J. Med. 1988, 319, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Japanese Encephalitis. Available online: https://www.who.int/en/news-room/fact-sheets/detail/japanese-encephalitis (accessed on 21 July 2020).
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, present, and future of Japanese encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef]
- Markoff, L. Points to consider in the development of a surrogate for efficacy of novel Japanese encephalitis virus vaccines. Vaccine 2000, 18, 26–32. [Google Scholar] [CrossRef]
- Erra, E.O.; Askling, H.H.; Yoksan, S.; Rombo, L.; Riutta, J.; Vene, S.; Lindquist, L.; Vapalahti, O.; Kantele, A. Cross-Protective Capacity of Japanese Encephalitis (JE) Vaccines Against Circulating Heterologous JE Virus Genotypes. Clin. Infect. Dis. 2013, 56, 267–270. [Google Scholar] [CrossRef] [Green Version]
- Bonaparte, M.; Dweik, B.; Feroldi, E.; Meric, C.; Bouckenooghe, A.; Hildreth, S.; Hu, B.; Yoksan, S.; Boaz, M. Immune response to live-attenuated Japanese encephalitis vaccine (JE-CV) neutralizes Japanese encephalitis virus isolates from South-East Asia and India. BMC Infect. Dis. 2014, 14, 156. [Google Scholar] [CrossRef] [Green Version]
- Hombach, J.; Barrett, A.D.; Cardosa, M.J.; Deubel, V.; Guzman, M.; Kurane, I.; Roehrig, J.T.; Sabchareon, A.; Kieny, M.P. Review on flavivirus vaccine development: Proceedings of a meeting jointly organised by the World Health Organization and the Thai Ministry of Public Health. In Proceedings of the Vaccine, Bangkok, Thailand, 26–27 April 2004; Elsevier BV: Amsterdam, The Netherlands, 2005; Volume 23, pp. 2689–2695. [Google Scholar]
- Van Gessel, Y.; Klade, C.S.; Putnak, R.; Formica, A.; Krasaesub, S.; Spruth, M.; Cena, B.; Tungtaeng, A.; Gettayacamin, M.; Dewasthaly, S. Correlation of protection against Japanese encephalitis virus and JE vaccine (IXIARO ®) induced neutralizing antibody titers. Vaccine 2011, 29, 5925–5931. [Google Scholar] [CrossRef]
- Leonova, G.N.; Pavlenko, E.V.; Maistrovskaya, O.S.; Chausov, E.V. Protective antibody titer for patients vaccinated against tickborne encephalitis virus. Procedia Vaccinol. 2011, 4, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Igarashi, T.; Haigwood, N.; Sadjadpour, R.; Plishka, R.J.; Buckler-White, A.; Shibata, R.; Martin, M.A. Determination of a Statistically Valid Neutralization Titer in Plasma That Confers Protection against Simian-Human Immunodeficiency Virus Challenge following Passive Transfer of High-Titered Neutralizing Antibodies. J. Virol. 2002, 76, 2123–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willey, R.; Nason, M.C.; Nishimura, Y.; Follmann, D.A.; Martin, M.A. Neutralizing Antibody Titers Conferring Protection to Macaques from a Simian/Human Immunodeficiency Virus Challenge Using the TZM-bl Assay. AIDS Res. Hum. Retroviruses 2010, 26, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Controlled Field Trial for an Evaluation of Effectiveness of Mouse-Brain Japanese Encephalitis Vaccine—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/4325804/ (accessed on 21 July 2020).
- Hoke, C.H.; Nisalak, A.; Sangawhipa, N.; Jatanasen, S.; Laorakapongse, T.; Innis, B.L.; Kotchasenee, S.O.; Gingrich, J.B.; Latendresse, J.; Fukai, K.; et al. Protection against Japanese Encephalitis by Inactivated Vaccines. N. Engl. J. Med. 1988, 319, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Kurane, I.; Takasaki, T. Immunogenicity and protective efficacy of the current inactivated Japanese encephalitis vaccine against different Japanese encephalitis virus strains. Vaccine 2000, 18, 33–35. [Google Scholar] [CrossRef]
- Lyons, A.; Kanesa-thasan, N.; Kuschner, R.A.; Eckels, K.H.; Putnak, R.; Sun, W.; Burge, R.; Towle, A.C.; Wilson, P.; Tauber, E.; et al. A Phase 2 study of a purified, inactivated virus vaccine to prevent Japanese encephalitis. Vaccine 2007, 25, 3445–3453. [Google Scholar] [CrossRef]
- Sugawara, K.; Nishiyama, K.; Ishikawa, Y.; Abe, M.; Sonoda, K.; Komatsu, K.; Horikawa, Y.; Takeda, K.; Honda, T.; Kuzuhara, S.; et al. Development of vero cell-derived inactivated Japanese encephalitis vaccine. Biologicals 2002, 30, 303–314. [Google Scholar] [CrossRef]
- Singh, A.; Mitra, M.; Sampath, G.; Venugopal, P.; Rao, J.V.; Krishnamurthy, B.; Gupta, M.K.; Sri Krishna, S.; Sudhakar, B.; Rao, N.B.; et al. A Japanese Encephalitis Vaccine From India Induces Durable and Cross-protective Immunity Against Temporally and Spatially Wide-ranging Global Field Strains. J. Infect. Dis. 2015, 212, 715–725. [Google Scholar] [CrossRef]
- Yu, Y. Phenotypic and genotypic characteristics of Japanese encephalitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine 2010, 28, 3635–3641. [Google Scholar] [CrossRef]
- Kanesa-thasan, N.; Smucny, J.J.; Hoke, C.H., Jr.; Marks, D.H.; Konishi, E.; Kurane, I.; Tang, D.B.; Vaughn, D.W.; Mason, P.W.; Shope, R.E. Safety and immunogenicity of NYVAC-JEV and ALVAC-JEV attenuated recombinant Japanese encephalitis virus—Poxvirus vaccines in vaccinia-nonimmune and vaccinia-immune humans. Vaccine 2000, 19, 483–491. [Google Scholar] [CrossRef]
- Kim, D.S.; Houillon, G.; Jang, G.C.; Cha, S.-H.; Choi, S.-H.; Lee, J.; Kim, H.M.; Kim, J.H.; Kang, J.H.; Kim, J.-H.; et al. A randomized study of the immunogenicity and safety of Japanese Encephalitis Chimeric Virus Vaccine (JE-CV) in comparison with SA14-14-2 Vaccine in children in the Republic of Korea. Hum. Vaccin. Immunother. 2014, 10, 2656–2663. [Google Scholar] [CrossRef]
- Mason, P.W.; Dalrymple, J.M.; Gentry, M.K.; McCown, J.M.; Hoke, C.H.; Burke, D.S.; Fournier, M.J.; Mason, T.L. Molecular characterization of a neutralizing domain of the Japanese encephalitis virus structural glycoprotein. J. Gen. Virol. 1989, 70, 2037–2049. [Google Scholar] [CrossRef]
- Chia, S.C.; Leung, P.S.C.; Liao, C.P.; Huang, J.H.; Lee, S.T. Fragment of Japanese encephalitis virus envelope protein produced in Escherichia coli protects mice from virus challenge. Microb. Pathog. 2001, 31, 9–19. [Google Scholar] [CrossRef]
- Wu, H.H.; Chen, C.T.; Lin, Y.L.; Lee, S.T. Sub-fragments of the envelope gene are highly protective against the Japanese encephalitis virus lethal infection in DNA priming Protein boosting immunization strategies. Vaccine 2004, 22, 793–800. [Google Scholar] [CrossRef]
- Wu, S.C.; Yu, C.H.; Lin, C.W.; Chu, I.M. The domain III fragment of Japanese encephalitis virus envelope protein: Mouse immunogenicity and liposome adjuvanticity. Vaccine 2003, 21, 2516–2522. [Google Scholar] [CrossRef]
- Chowers, M.Y.; Lang, R.; Nassar, F.; Ben-David, D.; Giladi, M.; Rubinshtein, E.; Itzhaki, A.; Mishal, J.; Siegman-Igra, Y.; Kitzes, R.; et al. Clinical Characteristics of the West Nile Fever Outbreak, Israel, 2000. Emerg. Infect. Dis. 2001, 7, 675–678. [Google Scholar] [CrossRef]
- Tsai, T.F.; Popovici, F.; Cernescu, C.; Campbell, G.L.; Nedelcu, N.I. West Nile encephalitis epidemic in southeastern Romania. Lancet 1998, 352, 767–771. [Google Scholar] [CrossRef]
- Calistri, P. Epidemiology of West Nile in Europe and in the Mediterranean Basin. Open Virol. J. 2010, 4, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H. A Neurotropic Virus Isolated from the Blood of a Native of Uganda 1. Am. J. Trop. Med. Hyg. 1940, s1-20, 471–492. [Google Scholar] [CrossRef]
- Riccardo, F.; Bolici, F.; Fafangel, M.; Jovanovic, V.; Socan, M.; Klepac, P.; Plavsa, D.; Vasic, M.; Bella, A.; Diana, G.; et al. West Nile virus in Europe: After action reviews of preparedness and response to the 2018 transmission season in Italy, Slovenia, Serbia and Greece. Glob. Health 2020, 16, 47. [Google Scholar] [CrossRef]
- Suthar, M.S.; Aguirre, S.; Fernandez-Sesma, A. Innate Immune Sensing of Flaviviruses. PLoS Pathog. 2013, 9, e1003541. [Google Scholar] [CrossRef]
- CDC National Healthcare Safety Network Biovigilance Component Hemovigilance Module Surveillance Protocol. Available online: https://www.cdc.gov/nhsn/pdfs/biovigilance/bv-hv-protocol-current.pdf (accessed on 21 July 2020).
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A review of successful flavivirus vaccines and the problems with those flaviviruses for which vaccines are not yet available. Vaccine 2014, 32, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Zohrabian, A.; Hayes, E.B.; Petersen, L.R. Cost-effectiveness of West Nile virus vaccination. Emerg. Infect. Dis. 2006, 12, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Shankar, M.B.; Staples, J.E.; Meltzer, M.I.; Fischer, M. Cost effectiveness of a targeted age-based West Nile virus vaccination program. Vaccine 2017, 35, 3143–3151. [Google Scholar] [CrossRef] [PubMed]
- West Nile Virus Vaccines | NIH: National Institute of Allergy and Infectious Diseases. Available online: https://www.niaid.nih.gov/diseases-conditions/wnv-vaccines (accessed on 21 July 2020).
- Oliphant, T.; Nybakken, G.E.; Austin, S.K.; Xu, Q.; Bramson, J.; Loeb, M.; Throsby, M.; Fremont, D.H.; Pierson, T.C.; Diamond, M.S. Induction of Epitope-Specific Neutralizing Antibodies against West Nile Virus. J. Virol. 2007, 81, 11828–11839. [Google Scholar] [CrossRef] [Green Version]
- Brandler, S.; Tangy, F. Vaccines in development against West Nile virus. Viruses 2013, 5, 2384–2409. [Google Scholar] [CrossRef] [Green Version]
- Ulbert, S. West Nile virus vaccines–current situation and future directions. Hum. Vaccines Immunother. 2019, 15, 2337–2342. [Google Scholar] [CrossRef] [Green Version]
- Coller, B.-A.; Pai, V.; Weeks-Levy, C.; Ogata, S.A. Recombinant Subunit West Nile Virus Vaccine for Protection of Human Subjects. U.S. Patent 20170165349A1, 1 June 2010. [Google Scholar]
- Lieberman, M.M.; Nerurkar, V.R.; Luo, H.; Cropp, B.; Carrion, R.; De La Garza, M.; Coller, B.A.; Clements, D.; Ogata, S.; Wong, T.; et al. Immunogenicity and protective efficacy of a recombinant subunit West Nile Virus vaccine in rhesus monkeys. Clin. Vaccine Immunol. 2009, 16, 1332–1337. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, S.E.; Karlsson, K.H.; Reimer, J.M.; Corbach-Söhle, S.; Patel, S.; Richner, J.M.; Nowotny, N.; Barzon, L.; Bengtsson, K.L.; Ulbert, S.; et al. Matrix-MTM adjuvanted envelope protein vaccine protects against lethal lineage 1 and 2 West Nile virus infection in mice. Vaccine 2014, 32, 800–808. [Google Scholar] [CrossRef]
- Martina, B.E.; Koraka, P.; van den Doel, P.; van Amerongen, G.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E. Immunization with West Nile virus envelope domain III protects mice against lethal infection with homologous and heterologous virus. Vaccine 2008, 26, 153–157. [Google Scholar] [CrossRef]
- Spohn, G.; Jennings, G.T.; Martina, B.E.; Keller, I.; Beck, M.; Pumpens, P.; Osterhaus, A.D.; Bachmann, M.F. A VLP-based vaccine targeting domain III of the West Nile virus E protein protects from lethal infection in mice. Virol. J. 2010, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- McDonald, W.F.; Huleatt, J.W.; Foellmer, H.G.; Hewitt, D.; Tang, J.; Desai, P.; Price, A.; Jacobs, A.; Takahashi, V.N.; Huang, Y.; et al. A West Nile Virus Recombinant Protein Vaccine That Coactivates Innate and Adaptive Immunity. J. Infect. Dis. 2007, 195, 1607–1617. [Google Scholar] [CrossRef]
- Wang, T.; Anderson, J.F.; Magnarelli, L.A.; Wong, S.J.; Koski, R.A.; Fikrig, E. Immunization of Mice Against West Nile Virus with Recombinant Envelope Protein. J. Immunol. 2001, 167, 5273–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Filette, M.; Soehle, S.; Ulbert, S.; Richner, J.; Diamond, M.S.; Sinigaglia, A.; Barzon, L.; Roels, S.; Lisziewicz, J.; Lorincz, O.; et al. Vaccination of Mice Using the West Nile Virus E-Protein in a DNA Prime-Protein Boost Strategy Stimulates Cell-Mediated Immunity and Protects Mice against a Lethal Challenge. PLoS ONE 2014, 9, e87837. [Google Scholar] [CrossRef] [PubMed]
- Tinker, J.; Yan, J.; Knippel, R.; Panayiotou, P.; Cornell, K. Immunogenicity of a West Nile Virus DIII-Cholera Toxin A2/B Chimera after Intranasal Delivery. Toxins 2014, 6, 1397–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.A.; Haque, M.; Stanfield, B.; Andrews, F.M.; Roy, A.A.; Kousoulas, K.G. A recombinant fusion protein consisting of West Nile virus envelope domain III fused in-frame with equine CD40 ligand induces antiviral immune responses in horses. Vet. Microbiol. 2017, 198, 51–58. [Google Scholar] [CrossRef]
- Chu, J.-H.J.; Chiang, C.-C.S.; Ng, M.-L. Immunization of Flavivirus West Nile Recombinant Envelope Domain III Protein Induced Specific Immune Response and Protection against West Nile Virus Infection. J. Immunol. 2007, 178, 2699–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlatkovic, J.; Stiasny, K.; Heinz, F.X. Immunodominance and Functional Activities of Antibody Responses to Inactivated West Nile Virus and Recombinant Subunit Vaccines in Mice. J. Virol. 2011, 85, 1994–2003. [Google Scholar] [CrossRef] [Green Version]
- Schneeweiss, A.; Chabierski, S.; Salomo, M.; Delaroque, N.; Al-Robaiy, S.; Grunwald, T.; Bürki, K.; Liebert, U.G.; Ulbert, S. A DNA vaccine encoding the E protein of West Nile Virus is protective and can be boosted by recombinant domain DIII. Vaccine 2011, 29, 6352–6357. [Google Scholar] [CrossRef]
- Friedrich, S.; Schmidt, T.; Schierhorn, A.; Lilie, H.; Szczepankiewicz, G.; Bergs, S.; Liebert, U.G.; Golbik, R.P.; Behrens, S.-E. Arginine methylation enhances the RNA chaperone activity of the West Nile virus host factor AUF1 p45. RNA 2016, 22, 1574–1591. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, N.K.; Karothia, D.; Shrivastava, A.; Banger, S.; Kumar, J.S. Enhanced production and immunological characterization of recombinant West Nile virus envelope domain III protein. N. Biotechnol. 2018, 46, 7–13. [Google Scholar] [CrossRef]
- Ledizet, M.; Kar, K.; Foellmer, H.G.; Wang, T.; Bushmich, S.L.; Anderson, J.F.; Fikrig, E.; Koski, R.A. A recombinant envelope protein vaccine against West Nile virus. Vaccine 2005, 23, 3915–3924. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.W.; Nguyen, H.-Y.; Hanna, S.L.; Sánchez, M.D.; Doms, R.W.; Pierson, T.C. West Nile Virus Discriminates between DC-SIGN and DC-SIGNR for Cellular Attachment and Infection. J. Virol. 2006, 80, 1290–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfert, M.A.; Boons, G.J. Adaptive immune activation: Glycosylation does matter. Nat. Chem. Biol. 2013, 9, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Buenbrazo, N.; Kell, L.; Rahmani, S.; Sim, L.; Withers, S.G.; DeFrees, S.; Wakarchuk, W. A Bacterial Expression Platform for Production of Therapeutic Proteins Containing Human-like O-Linked Glycans. Cell Chem. Biol. 2019, 26, 203–212.e5. [Google Scholar] [CrossRef] [PubMed]
- Kanno, A.I.; de Cerqueira Leite, L.C.; Pereira, L.R.; de Jesus, M.J.R.; Andreata-Santos, R.; dos Santos Alves, R.P.; Durigon, E.L.; de Souza Ferreira, L.C.; Gonçalves, V.M. Optimization and scale-up production of Zika virus ΔNS1 in Escherichia coli: Application of Response Surface Methodology. AMB Express 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef]
- Hadinegoro, S.R.S. The revised WHO dengue case classification: Does the system need to be modified? Paediatr. Int. Child Health 2012, 32, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Luedtke, A.; Langevin, E.; Zhu, M.; Bonaparte, M.; Machabert, T.; Savarino, S.; Zambrano, B.; Moureau, A.; Khromava, A.; et al. Effect of Dengue Serostatus on Dengue Vaccine Safety and Efficacy. N. Engl. J. Med. 2018, 379, 327–340. [Google Scholar] [CrossRef]
- Vaughn, D.W. Invited Commentary: Dengue Lessons from Cuba. Am. J. Epidemiol. 2000, 152, 800–803. [Google Scholar] [CrossRef]
- Guzmán, M.G.; Kourí, G.; Valdés, L.; Bravo, J.; Vázquez, S.; Halstead, S.B. Enhanced severity of secondary dengue-2 infections: Death rates in 1981 and 1997 Cuban outbreaks. Rev. Panam. Salud Publica/Pan Am. J. Public Health 2002, 11, 223–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Dengue vaccines: WHO position September 2018. Wkly. Epidemiol. Rec. 2018, 93, 457–476.
- Nhan, N.T.; Phuong, C.X.T.; Kneen, R.; Wills, B.; Van My, N.; Phuong, N.T.Q.; Van Thien, C.; Nga, N.T.T.; Simpson, J.A.; Solomon, T.; et al. Acute Management of Dengue Shock Syndrome: A Randomized Double-Blind Comparison of 4 Intravenous Fluid Regimens in the First Hour. Clin. Infect. Dis. 2001, 32, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Villar, L.; Dayan, G.H.; Arredondo-García, J.L.; Rivera, D.M.; Cunha, R.; Deseda, C.; Reynales, H.; Costa, M.S.; Morales-Ramírez, J.O.; Carrasquilla, G.; et al. Efficacy of a Tetravalent Dengue Vaccine in Children in Latin America. N. Engl. J. Med. 2015, 372, 113–123. [Google Scholar] [CrossRef]
- Redoni, M.; Yacoub, S.; Rivino, L.; Giacobbe, D.R.; Luzzati, R.; Di Bella, S. Dengue: Status of current and under-development vaccines. Rev. Med. Virol. 2020, 30, e2101. [Google Scholar] [CrossRef] [PubMed]
- WHO Dengue vaccine: WHO position paper, September 2018—Recommendations. Vaccine 2019, 37, 4848–4849. [CrossRef]
- Pinheiro-Michelsen, J.R.; Souza, R.D.S.O.; Santana, I.V.R.; da Silva, P.D.S.; Mendez, E.C.; Luiz, W.B.; Amorim, J.H. Anti-dengue Vaccines: From Development to Clinical Trials. Front. Immunol. 2020, 11, 1252. [Google Scholar] [CrossRef]
- Sáez-Llorens, X.; Tricou, V.; Yu, D.; Rivera, L.; Jimeno, J.; Villarreal, A.C.; Dato, E.; Mazara, S.; Vargas, M.; Brose, M.; et al. Immunogenicity and safety of one versus two doses of tetravalent dengue vaccine in healthy children aged 2–17 years in Asia and Latin America: 18-month interim data from a phase 2, randomised, placebo-controlled study. Lancet Infect. Dis. 2018, 18, 162–170. [Google Scholar] [CrossRef]
- Sáez-Llorens, X.; Tricou, V.; Yu, D.; Rivera, L.; Tuboi, S.; Garbes, P.; Borkowski, A.; Wallace, D. Safety and immunogenicity of one versus two doses of Takeda’s tetravalent dengue vaccine in children in Asia and Latin America: Interim results from a phase 2, randomised, placebo-controlled study. Lancet Infect. Dis. 2017, 17, 615–625. [Google Scholar] [CrossRef]
- Rupp, R.; Luckasen, G.J.; Kirstein, J.L.; Osorio, J.E.; Santangelo, J.D.; Raanan, M.; Smith, M.K.; Wallace, D.; Gordon, G.S.; Stinchcomb, D.T. Safety and immunogenicity of different doses and schedules of a live attenuated tetravalent dengue vaccine (TDV) in healthy adults: A Phase 1b randomized study. Vaccine 2015, 33, 6351–6359. [Google Scholar] [CrossRef] [Green Version]
- Kirkpatrick, B.D.; Durbin, A.P.; Pierce, K.K.; Carmolli, M.P.; Tibery, C.M.; Grier, P.L.; Hynes, N.; Diehl, S.A.; Elwood, D.; Jarvis, A.P.; et al. Robust and Balanced Immune Responses to All 4 Dengue Virus Serotypes Following Administration of a Single Dose of a Live Attenuated Tetravalent Dengue Vaccine to Healthy, Flavivirus-Naive Adults. J. Infect. Dis. 2015, 212, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P.; Kirkpatrick, B.D.; Pierce, K.K.; Elwood, D.; Larsson, C.J.; Lindow, J.C.; Tibery, C.; Sabundayo, B.P.; Shaffer, D.; Talaat, K.R.; et al. A Single Dose of Any of Four Different Live Attenuated Tetravalent Dengue Vaccines Is Safe and Immunogenic in Flavivirus-naive Adults: A Randomized, Double-blind Clinical Trial. J. Infect. Dis. 2013, 207, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Biswal, S.; Borja-Tabora, C.; Martinez Vargas, L.; Velásquez, H.; Theresa Alera, M.; Sierra, V.; Johana Rodriguez-Arenales, E.; Yu, D.; Wickramasinghe, V.P.; Duarte Moreira, E.; et al. Efficacy of a tetravalent dengue vaccine in healthy children aged 4–16 years: A randomised, placebo-controlled, phase 3 trial. Lancet 2020, 395, 1423–1433. [Google Scholar] [CrossRef]
- Murphy, B.R.; Whitehead, S.S. Immune Response to Dengue Virus and Prospects for a Vaccine. Annu. Rev. Immunol. 2011, 29, 587–619. [Google Scholar] [CrossRef] [PubMed]
- Balsitis, S.J.; Williams, K.L.; Lachica, R.; Flores, D.; Kyle, J.L.; Mehlhop, E.; Johnson, S.; Diamond, M.S.; Beatty, P.R.; Harris, E. Lethal Antibody Enhancement of Dengue Disease in Mice Is Prevented by Fc Modification. PLoS Pathog. 2010, 6, e1000790. [Google Scholar] [CrossRef] [Green Version]
- Waggoner, J.J.; Katzelnick, L.C.; Burger-Calderon, R.; Gallini, J.; Moore, R.H.; Kuan, G.; Balmaseda, A.; Pinsky, B.A.; Harris, E. Antibody-Dependent Enhancement of Severe Disease Is Mediated by Serum Viral Load in Pediatric Dengue Virus Infections. J. Infect. Dis. 2020, 221, 1846–1854. [Google Scholar] [CrossRef] [PubMed]
- Capeding, M.R.; Tran, N.H.; Hadinegoro, S.R.S.; Ismail, H.I.H.M.; Chotpitayasunondh, T.; Chua, M.N.; Luong, C.Q.; Rusmil, K.; Wirawan, D.N.; Nallusamy, R.; et al. Clinical efficacy and safety of a novel tetravalent dengue vaccine in healthy children in Asia: A phase 3, randomised, observer-masked, placebo-controlled trial. Lancet 2014, 384, 1358–1365. [Google Scholar] [CrossRef]
- Weiskopf, D.; Cerpas, C.; Angelo, M.A.; Bangs, D.J.; Sidney, J.; Paul, S.; Peters, B.; Sanches, F.P.; Silvera, C.G.T.; Costa, P.R.; et al. Human CD8 + T-Cell Responses Against the 4 Dengue Virus Serotypes Are Associated With Distinct Patterns of Protein Targets. J. Infect. Dis. 2015, 212, 1743–1751. [Google Scholar] [CrossRef] [Green Version]
- Elong Ngono, A.; Chen, H.-W.; Tang, W.W.; Joo, Y.; King, K.; Weiskopf, D.; Sidney, J.; Sette, A.; Shresta, S. Protective Role of Cross-Reactive CD8 T Cells Against Dengue Virus Infection. EBioMedicine 2016, 13, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Amorim, J.H.; dos Santos Alves, R.P.; Bizerra, R.; Araújo Pereira, S.; Ramos Pereira, L.; Nascimento Fabris, D.L.; Santos, R.A.; Romano, C.M.; de Souza Ferreira, L.C. Antibodies are not required to a protective immune response against dengue virus elicited in a mouse encephalitis model. Virology 2016, 487, 41–49. [Google Scholar] [CrossRef]
- Weiskopf, D.; Angelo, M.A.; Bangs, D.J.; Sidney, J.; Paul, S.; Peters, B.; de Silva, A.D.; Lindow, J.C.; Diehl, S.A.; Whitehead, S.; et al. The Human CD8 + T Cell Responses Induced by a Live Attenuated Tetravalent Dengue Vaccine Are Directed against Highly Conserved Epitopes. J. Virol. 2015, 89, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Grifoni, A.; Sette, A.; Weiskopf, D. Human T Cell Response to Dengue Virus Infection. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mladinich, K.M.; Piaskowski, S.M.; Rudersdorf, R.; Eernisse, C.M.; Weisgrau, K.L.; Martins, M.A.; Furlott, J.R.; Partidos, C.D.; Brewoo, J.N.; Osorio, J.E.; et al. Dengue virus-specific CD4+ and CD8+ T lymphocytes target NS1, NS3 and NS5 in infected Indian rhesus macaques. Immunogenetics 2012, 64, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Manoff, S.B.; Sausser, M.; Falk Russell, A.; Martin, J.; Radley, D.; Hyatt, D.; Roberts, C.C.; Lickliter, J.; Krishnarajah, J.; Bett, A.; et al. Immunogenicity and safety of an investigational tetravalent recombinant subunit vaccine for dengue: Results of a Phase I randomized clinical trial in flavivirus-naïve adults. Hum. Vaccin. Immunother. 2019, 15, 2195–2204. [Google Scholar] [CrossRef]
- Clements, D.E.; Coller, B.-A.G.; Lieberman, M.M.; Ogata, S.; Wang, G.; Harada, K.E.; Putnak, J.R.; Ivy, J.M.; McDonell, M.; Bignami, G.S.; et al. Development of a recombinant tetravalent dengue virus vaccine: Immunogenicity and efficacy studies in mice and monkeys. Vaccine 2010, 28, 2705–2715. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, D.; Meschino, S.; Guan, L.; Clements, D.E.; ter Meulen, J.H.; Casimiro, D.R.; Coller, B.-A.G.; Bett, A.J. Preclinical development of a dengue tetravalent recombinant subunit vaccine: Immunogenicity and protective efficacy in nonhuman primates. Vaccine 2015, 33, 4105–4116. [Google Scholar] [CrossRef]
- Zulueta, A.; Hermida, L.; Lazo, L.; Valdés, I.; Rodríguez, R.; López, C.; Silva, R.; Rosario, D.; Martín, J.; Guzmán, M.G.; et al. The fusion site of envelope fragments from each serotype of Dengue virus in the P64k protein, influence some parameters of the resulting chimeric constructs. Biochem. Biophys. Res. Commun. 2003, 308, 619–626. [Google Scholar] [CrossRef]
- Hermida, L.; Bernardo, L.; Martin, J.; Alvarez, M.; Prado, I.; Lopez, C.; Sierra, B.; Martinez, R.; Rodriguez, R.; Zulueta, A. A recombinant fusion protein containing the domain III of the dengue-2 envelope protein is immunogenic and protective in nonhuman primates. Vaccine 2006, 24, 3165–3171. [Google Scholar] [CrossRef]
- Bernardo, L.; Hermida, L.; Martin, J.; Alvarez, M.; Prado, I.; López, C.; Martínez, R.; Rodríguez-Roche, R.; Zulueta, A.; Lazo, L.; et al. Anamnestic antibody response after viral challenge in monkeys immunized with dengue 2 recombinant fusion proteins. Arch. Virol. 2008, 153, 849–854. [Google Scholar] [CrossRef]
- Batista, I.C.A.; Quinan, B.R.; Alves, É.A.R.; Jangola, S.T.G.; Oliveira, E.S.; Colombarolli, S.G.; Ferreira, J.G.G.; de Oliveira Rocha, E.S.; Kroon, E.G.; de Assis, R.R.; et al. Design and production of dengue virus chimeric proteins useful for developing tetravalent vaccines. Vaccine 2020, 38, 2005–2015. [Google Scholar] [CrossRef]
- Ganguly, A.; Malabadi, R.B.; Das, D.; Suresh, M.R.; Sunwoo, H.H. Enhanced Prokaryotic Expression of Dengue Virus Envelope Protein. J. Pharm. Pharm. Sci. 2013, 16, 609. [Google Scholar] [CrossRef] [PubMed]
- Simmons, M.; Hayes, C.G.; Wu, S.J.; Nelson, W.M. Evaluation of the protective efficacy of a recombinant dengue envelope B domain fusion protein against dengue 2 virus infection in mice. Am. J. Trop. Med. Hyg. 1998, 58, 655–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, C.G.; Simmons, M.; Kochel, T.; Raviprakash, K.; Murphy, G.S. Characterization of antibody responses to combinations of a dengue-2 DNA and dengue-2 recombinant subunit vaccine. Am. J. Trop. Med. Hyg. 2001, 65, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Simmons, M.; Porter, K.R.; Hayes, C.G.; Vaughn, D.W.; Putnak, R. Characterization of Antibody Responses to Combinations of a Dengue Virus Type 2 DNA Vaccine and Two Dengue Virus Type 2 Protein Vaccines in Rhesus Macaques. J. Virol. 2006, 80, 9577–9585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, C.G.; Simmons, M.; Murphy, G.S. Short report: Antibody responses of mice immunized with a tetravalent dengue recombinant protein subunit vaccine. Am. J. Trop. Med. Hyg. 2001, 65, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Suzarte, E.; Marcos, E.; Gil, L.; Valdés, I.; Lazo, L.; Ramos, Y.; Pérez, Y.; Falcón, V.; Romero, Y.; Guzmán, M.G.; et al. Generation and characterization of potential dengue vaccine candidates based on domain III of the envelope protein and the capsid protein of the four serotypes of dengue virus. Arch. Virol. 2014, 159, 1629–1640. [Google Scholar] [CrossRef]
- Gil, L.; Lazo, L.; Valdés, I.; Suzarte, E.; Yen, P.; Ramírez, R.; Álvarez, M.; Dung, L.T.; Cobas, K.; Marcos, E.; et al. The tetravalent formulation of domain III-capsid proteins recalls memory B- and T-cell responses induced in monkeys by an experimental dengue virus infection. Clin. Transl. Immunol. 2017, 6, e148. [Google Scholar] [CrossRef]
- Suzarte, E.; Gil, L.; Valdés, I.; Marcos, E.; Lazo, L.; Izquierdo, A.; García, A.; López, L.; Álvarez, M.; Pérez, Y.; et al. A novel tetravalent formulation combining the four aggregated domain III-capsid proteins from dengue viruses induces a functional immune response in mice and monkeys. Int. Immunol. 2015, 27, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Gil, L.; Marcos, E.; Izquierdo, A.; Lazo, L.; Valdés, I.; Ambala, P.; Ochola, L.; Hitler, R.; Suzarte, E.; Álvarez, M.; et al. The protein DIIIC-2, aggregated with a specific oligodeoxynucleotide and adjuvanted in alum, protects mice and monkeys against DENV-2. Immunol. Cell Biol. 2015, 93, 57–66. [Google Scholar] [CrossRef]
- Zuest, R.; Valdes, I.; Skibinski, D.; Lin, Y.; Toh, Y.X.; Chan, K.; Hermida, L.; Connolly, J.; Guillen, G.; Fink, K. Tetravalent dengue DIIIC protein together with alum and ODN elicits a Th1 response and neutralizing antibodies in mice. Vaccine 2015, 33, 1474–1482. [Google Scholar] [CrossRef]
- Valdés, I.; Izquierdo, A.; Cobas, K.; Thao, P.; Anh Duc, H.; Duc Loc, H.; Dung, L.T.; Lazo, L.; Suzarte, E.; Pérez, Y.; et al. A heterologous prime-boost strategy for immunization against Dengue virus combining the Tetra DIIIC subunit vaccine candidate with the TV005 live-attenuated tetravalent vaccine. J. Gen. Virol. 2019, 100, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Valdés, I.; Bernardo, L.; Gil, L.; Pavón, A.; Lazo, L.; López, C.; Romero, Y.; Menendez, I.; Falcón, V.; Betancourt, L.; et al. A novel fusion protein domain III-capsid from dengue-2, in a highly aggregated form, induces a functional immune response and protection in mice. Virology 2009, 394, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-H.; Yang, S.-P.; Tsai, M.-J.; Lin, G.-C.; Wu, H.-C.; Wu, S.-C. Dengue and Zika Virus Domain III-Flagellin Fusion and Glycan-Masking E Antigen for Prime-Boost Immunization. Theranostics 2019, 9, 4811–4826. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yu, M.; Jiang, T.; Deng, Y.; Qin, C.; Qin, E. Induction of Tetravalent Protective Immunity Against Four Dengue Serotypes by the Tandem Domain III of the Envelope Protein. DNA Cell Biol. 2007, 26, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Frei, J.C.; Wirchnianski, A.S.; Govero, J.; Vergnolle, O.; Dowd, K.A.; Pierson, T.C.; Kielian, M.; Girvin, M.E.; Diamond, M.S.; Lai, J.R. Engineered Dengue Virus Domain III Proteins Elicit Cross-Neutralizing Antibody Responses in Mice. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Maeda, D.L.N.F.; Batista, M.T.; Pereira, L.R.; de Jesus Cintra, M.; Amorim, J.H.; Mathias-Santos, C.; Pereira, S.A.; Boscardin, S.B.; Silva, S.D.R.; Faquim-Mauro, E.L.; et al. Adjuvant-Mediated Epitope Specificity and Enhanced Neutralizing Activity of Antibodies Targeting Dengue Virus Envelope Protein. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, J.; Chen, W.; Hu, Z.; Zhu, J.; Fang, X.; Yuan, W.; Li, M.; Hu, X.; Tan, Y.; et al. Eliciting cross-neutralizing antibodies in mice challenged with a dengue virus envelope domain III expressed in Escherichia coli. Can. J. Microbiol. 2012, 58, 369–380. [Google Scholar] [CrossRef]
- Chiang, C.-Y.; Pan, C.-H.; Hsieh, C.-H.; Tsai, J.-P.; Chen, M.-Y.; Liu, H.-H.; Liu, S.-J.; Chong, P.; Leng, C.-H.; Chen, H.-W. Lipidated Dengue-2 Envelope Protein Domain III Independently Stimulates Long-Lasting Neutralizing Antibodies and Reduces the Risk of Antibody-Dependent Enhancement. PLoS Negl. Trop. Dis. 2013, 7, e2432. [Google Scholar] [CrossRef]
- Chiang, C.-Y.; Huang, M.-H.; Pan, C.-H.; Hsieh, C.-H.; Chen, M.-Y.; Liu, H.-H.; Tsai, J.-P.; Liu, S.-J.; Chong, P.; Leng, C.-H.; et al. Induction of robust immunity by the emulsification of recombinant lipidated dengue-1 envelope protein domain III. Microbes Infect. 2013, 15, 719–728. [Google Scholar] [CrossRef]
- Chiang, C.-Y.; Hsieh, C.-H.; Chen, M.-Y.; Tsai, J.-P.; Liu, H.-H.; Liu, S.-J.; Chong, P.; Leng, C.-H.; Chen, H.-W. Recombinant lipidated dengue-4 envelope protein domain III elicits protective immunity. Vaccine 2014, 32, 1346–1353. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.-Y.; Liu, S.-J.; Hsieh, C.-H.; Chen, M.-Y.; Tsai, J.-P.; Liu, H.-H.; Chen, I.-H.; Chong, P.; Leng, C.-H.; Chen, H.-W. Recombinant lipidated dengue-3 envelope protein domain III stimulates broad immune responses in mice. Vaccine 2016, 34, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.-Y.; Huang, M.-H.; Hsieh, C.-H.; Chen, M.-Y.; Liu, H.-H.; Tsai, J.-P.; Li, Y.-S.; Chang, C.-Y.; Liu, S.-J.; Chong, P.; et al. Dengue-1 Envelope Protein Domain III along with PELC and CpG Oligodeoxynucleotides Synergistically Enhances Immune Responses. PLoS Negl. Trop. Dis. 2012, 6, e1645. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Jiang, T.; Zhou, X.-Z.; Deng, Y.-Q.; Li, X.-F.; Chen, S.-P.; Zhu, S.-Y.; Zhou, X.; Qin, E.-D.; Qin, C.-F. Induction of Neutralizing Antibodies against Four Serotypes of Dengue Viruses by MixBiEDIII, a Tetravalent Dengue Vaccine. PLoS ONE 2014, 9, e86573. [Google Scholar] [CrossRef] [PubMed]
- ZHANG, Z.-S.; WENG, Y.-W.; HUANG, H.-L.; ZHANG, J.-M.; YAN, Y.-S. Neutralizing antibodies respond to a bivalent dengue DNA vaccine or/and a recombinant bivalent antigen. Mol. Med. Rep. 2015, 11, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-S.; Yan, Y.-S.; Weng, Y.-W.; Huang, H.-L.; Li, S.-Q.; He, S.; Zhang, J.-M. High-level expression of recombinant dengue virus type 2 envelope domain III protein and induction of neutralizing antibodies in BALB/C mice. J. Virol. Methods 2007, 143, 125–131. [Google Scholar] [CrossRef]
- Fahimi, H.; Allahyari, H.; Hassan, Z.M.; Sadeghizadeh, M. Dengue virus type-3 envelope protein domain III; expression and immunogenicity. Iran. J. Basic Med. Sci. 2014, 17, 836–843. [Google Scholar]
- Leng, C.-H.; Liu, S.-J.; Tsai, J.-P.; Li, Y.-S.; Chen, M.-Y.; Liu, H.-H.; Lien, S.-P.; Yueh, A.; Hsiao, K.-N.; Lai, L.-W.; et al. A novel dengue vaccine candidate that induces cross-neutralizing antibodies and memory immunity. Microbes Infect. 2009, 11, 288–295. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Shrivastava, A. Evaluation of antibody response against recombinant domain III proteins of dengue virus type 1 and 2. AIMS Microbiol. 2017, 3, 248–266. [Google Scholar] [CrossRef]
- Afroz, S.; Shama; Battu, S.; Matin, S.; Solouki, S.; Elmore, J.P.; Minhas, G.; Huang, W.; August, A.; Khan, N. Amino acid starvation enhances vaccine efficacy by augmenting neutralizing antibody production. Sci. Signal. 2019, 12, eaav4717. [Google Scholar] [CrossRef]
- Arora, U.; Tyagi, P.; Swaminathan, S.; Khanna, N. Chimeric Hepatitis B core antigen virus-like particles displaying the envelope domain III of dengue virus type 2. J. Nanobio. 2012, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Rathore, A.S.; Sarker, A.; Gupta, R.D. Production and immunogenicity of Fubc subunit protein redesigned from DENV envelope protein. Appl. Microbiol. Biotechnol. 2020, 104, 4333–4344. [Google Scholar] [CrossRef] [PubMed]
- Amorim, J.H.; Porchia, B.F.M.M.; Balan, A.; Cavalcante, R.C.M.; da Costa, S.M.; de Barcelos Alves, A.M.; de Souza Ferreira, L.C. Refolded dengue virus type 2 NS1 protein expressed in Escherichia coli preserves structural and immunological properties of the native protein. J. Virol. Methods 2010, 167, 186–192. [Google Scholar] [CrossRef]
- Rouvinski, A.; Guardado-Calvo, P.; Barba-Spaeth, G.; Duquerroy, S.; Vaney, M.-C.; Kikuti, C.M.; Navarro Sanchez, M.E.; Dejnirattisai, W.; Wongwiwat, W.; Haouz, A.; et al. Recognition determinants of broadly neutralizing human antibodies against dengue viruses. Nature 2015, 520, 109–113. [Google Scholar] [CrossRef]
- Li, L.; Meng, W.; Horton, M.; DiStefano, D.R.; Thoryk, E.A.; Pfaff, J.M.; Wang, Q.; Salazar, G.T.; Barnes, T.; Doranz, B.J.; et al. Potent neutralizing antibodies elicited by dengue vaccine in rhesus macaque target diverse epitopes. PLOS Pathog. 2019, 15, e1007716. [Google Scholar] [CrossRef]
- Rodriguez-Barraquer, I.; Costa, F.; Nascimento, E.J.M.; Júnior, N.N.; Castanha, P.M.S.; Sacramento, G.A.; Cruz, J.; Carvalho, M.; De Olivera, D.; Hagan, J.E.; et al. Impact of preexisting dengue immunity on Zika virus emergence in a dengue endemic region. Science 2019, 363, 607–610. [Google Scholar] [CrossRef] [Green Version]
- Einspieler, C.; Utsch, F.; Brasil, P.; Panvequio Aizawa, C.Y.; Peyton, C.; Hydee Hasue, R.; Françoso Genovesi, F.; Damasceno, L.; Moreira, M.E.; Adachi, K.; et al. Association of Infants Exposed to Prenatal Zika Virus Infection With Their Clinical, Neurologic, and Developmental Status Evaluated via the General Movement Assessment Tool. JAMA Netw. Open 2019, 2, e187235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulkey, S.B.; Arroyave-Wessel, M.; Peyton, C.; Bulas, D.I.; Fourzali, Y.; Jiang, J.; Russo, S.; McCarter, R.; Msall, M.E.; Du Plessis, A.J.; et al. Neurodevelopmental Abnormalities in Children with in Utero Zika Virus Exposure Without Congenital Zika Syndrome. JAMA Pediatr. 2020, 174, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Poland, G.A.; Ovsyannikova, I.G.; Kennedy, R.B. Zika Vaccine Development: Current Status. Mayo Clin. Proc. 2019, 94, 2572–2586. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Fernandez, E.; Dowd, K.A.; Speer, S.D.; Platt, D.J.; Gorman, M.J.; Govero, J.; Nelson, C.A.; Pierson, T.C.; Diamond, M.S.; et al. Structural Basis of Zika Virus-Specific Antibody Protection. Cell 2016, 166, 1016–1027. [Google Scholar] [CrossRef] [Green Version]
- Larocca, R.A.; Abbink, P.; Peron, J.P.S.; Zanotto, P.M.D.A.; Iampietro, M.J.; Badamchi-Zadeh, A.; Boyd, M.; Ng’ang’a, D.; Kirilova, M.; Nityanandam, R.; et al. Vaccine protection against Zika virus from Brazil. Nature 2016, 536, 474–478. [Google Scholar] [CrossRef]
- Weiskopf, D.; Angelo, M.A.; de Azeredo, E.L.; Sidney, J.; Greenbaum, J.A.; Fernando, A.N.; Broadwater, A.; Kolla, R.V.; De Silva, A.D.; de Silva, A.M.; et al. Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+ T cells. Proc. Natl. Acad. Sci. USA 2013, 110, E2046–E2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, C.; Delgado, F.; Simon-Lorière, E.; Sakuntabhai, A. Immune Responses to Dengue and Zika Viruses—Guidance for T Cell Vaccine Development. Int. J. Environ. Res. Public Health 2018, 15, 385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elong Ngono, A.; Vizcarra, E.A.; Tang, W.W.; Sheets, N.; Joo, Y.; Kim, K.; Gorman, M.J.; Diamond, M.S.; Shresta, S. Mapping and Role of the CD8+ T Cell Response During Primary Zika Virus Infection in Mice. Cell Host Microbe 2017, 21, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Gaudinski, M.R.; Houser, K.V.; Morabito, K.M.; Hu, Z.; Yamshchikov, G.; Rothwell, R.S.; Berkowitz, N.; Mendoza, F.; Saunders, J.G.; Novik, L.; et al. Safety, tolerability, and immunogenicity of two Zika virus DNA vaccine candidates in healthy adults: Randomised, open-label, phase 1 clinical trials. Lancet 2018, 391, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Han, J.F.; Qiu, Y.; Yu, J.Y.; Wang, H.J.; Deng, Y.Q.; Li, X.F.; Zhao, H.; Sun, H.X.; Qin, C.F. Immunization with truncated envelope protein of Zika virus induces protective immune response in mice. Sci. Rep. 2017, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Li, C.; Afridi, S.K.; Zu, S.; Xu, J.W.; Quanquin, N.; Yang, H.; Cheng, G.; Xu, Z. E90 subunit vaccine protects mice from Zika virus infection and microcephaly. Acta Neuropathol. Commun. 2018, 6, 77. [Google Scholar] [CrossRef]
- Liang, H.; Yang, R.; Liu, Z.; Li, M.; Liu, H.; Jin, X. Recombinant Zika virus envelope protein elicited protective immunity against Zika virus in immunocompetent mice. PLoS ONE 2018, 13, e0194860. [Google Scholar] [CrossRef] [Green Version]
- Amaral, M.P.; de Souza Apostolico, J.; Tomita, N.; Coirada, F.C.; Lunardelli, V.A.S.; Fernandes, E.R.; Souza, H.F.S.; Astray, R.M.; Boscardin, S.B.; Rosa, D.S. Homologous prime-boost with Zika virus envelope protein and poly (I:C) induces robust specific humoral and cellular immune responses. Vaccine 2020, 38, 3653–3664. [Google Scholar] [CrossRef]
- Yang, M.; Dent, M.; Lai, H.; Sun, H.; Chen, Q. Immunization of Zika virus envelope protein domain III induces specific and neutralizing immune responses against Zika virus. Vaccine 2017, 35, 4287–4294. [Google Scholar] [CrossRef]
- Garske, T.; Van Kerkhove, M.D.; Yactayo, S.; Ronveaux, O.; Lewis, R.F.; Staples, J.E.; Perea, W.; Ferguson, N.M. Yellow Fever in Africa: Estimating the Burden of Disease and Impact of Mass Vaccination from Outbreak and Serological Data. PLoS Med. 2014, 11, e1001638. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.-T.; Li, Q.-F.; Ma, C.; Zhao, Z.-X.; He, L.-F.; Tang, T.-T.; Yu, W.; Owiti, P. Reduction patterns of Japanese encephalitis incidence following vaccine introduction into long-term expanded program on immunization in Yunnan Province, China. Infect. Dis. Poverty 2019, 8, 102. [Google Scholar] [CrossRef]
- Detection and Investigation of Serious Adverse Events Following Yellow Fever Vaccination. Available online: https://apps.who.int/iris/bitstream/handle/10665/70251/WHO_HSE_GAR_ERI_2010.2_eng.pdf;jsessionid=00CE1944BF3BB748F23372FB80C8AA48?sequence=1 (accessed on 21 July 2020).
- Bae, H.; Domingo, C.; Tenorio, A.; de Ory, F.; Muñoz, J.; Weber, P.; Teuwen, D.E.; Niedrig, M. Immune Response during Adverse Events after 17D-Derived Yellow Fever Vaccination in Europe. J. Infect. Dis. 2008, 197, 1577–1584. [Google Scholar] [CrossRef]
- Sabetian, S.; Nezafat, N.; Dorosti, H.; Zarei, M.; Ghasemi, Y. Exploring dengue proteome to design an effective epitope-based vaccine against dengue virus. J. Biomol. Struct. Dyn. 2019, 37, 2546–2563. [Google Scholar] [CrossRef]
- Murphy, D.; Reche, P.; Flower, D.R. Selection-based design of in silico dengue epitope ensemble vaccines. Chem. Biol. Drug Des. 2019, 93, 21–28. [Google Scholar] [CrossRef]
- Ali, M.; Pandey, R.K.; Khatoon, N.; Narula, A.; Mishra, A.; Prajapati, V.K. Exploring dengue genome to construct a multi-epitope based subunit vaccine by utilizing immunoinformatics approach to battle against dengue infection. Sci. Rep. 2017, 7, 9232. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Recombinant Protein–Based Dengue Vaccines. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Tripathi, N.K. Production and Purification of Recombinant Proteins from Escherichia coli. ChemBioEng Rev. 2016, 3, 116–133. [Google Scholar] [CrossRef]
- Tripathi, N.K.; Shrivastava, A. Recent Developments in Bioprocessing of Recombinant Proteins: Expression Hosts and Process Development. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Pierson, T.C.; Fremont, D.H.; Kuhn, R.J.; Diamond, M.S. Structural Insights into the Mechanisms of Antibody-Mediated Neutralization of Flavivirus Infection: Implications for Vaccine Development. Cell Host Microbe 2008, 4, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Slon Campos, J.L.; Mongkolsapaya, J.; Screaton, G.R. The immune response against flaviviruses. Nat. Immunol. 2018, 19, 1189–1198. [Google Scholar] [CrossRef]
- Mamat, U.; Wilke, K.; Bramhill, D.; Schromm, A.B.; Lindner, B.; Kohl, T.A.; Corchero, J.L.; Villaverde, A.; Schaffer, L.; Head, S.R.; et al. Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Fact. 2015, 14, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilding, K.M.; Hunt, J.P.; Wilkerson, J.W.; Funk, P.J.; Swensen, R.L.; Carver, W.C.; Christian, M.L.; Bundy, B.C. Endotoxin-Free, E. coli- Based Cell-Free Protein Synthesis: Pre-Expression Endotoxin Removal Approaches for on-Demand Cancer Therapeutic Production. Biotechnol. J. 2019, 14, 1800271. [Google Scholar] [CrossRef] [PubMed]
- Chura-Chambi, R.M.; da Silva, C.M.R.; Pereira, L.R.; Bartolini, P.; Ferreira, L.C.D.S.; Morganti, L. Protein refolding based on high hydrostatic pressure and alkaline pH: Application on a recombinant dengue virus NS1 protein. PLoS ONE 2019, 14, e0211162. [Google Scholar] [CrossRef] [PubMed]
- Rosa da Silva, C.M.; Chura-Chambi, R.M.; Ramos Pereira, L.; Cordeiro, Y.; de Souza Ferreira, L.C.; Morganti, L. Association of high pressure and alkaline condition for solubilization of inclusion bodies and refolding of the NS1 protein from zika virus. BMC Biotechnol. 2018, 18, 78. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-B.; Xu, J. Better Adjuvants for Better Vaccines: Progress in Adjuvant Delivery Systems, Modifications, and Adjuvant–Antigen Codelivery. Vaccines 2020, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Bidet, K.; Ho, V.; Chu, C.W.; Naim, A.N.M.; Thazin, K.; Chan, K.R.; Low, J.G.H.; Choy, M.M.; Wong, L.H.; Florez de Sessions, P.; et al. Mimicking immune signatures of flavivirus infection with targeted adjuvants improves dengue subunit vaccine immunogenicity. npj Vaccines 2019, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.L.; Soema, P.C.; Slütter, B.; Ossendorp, F.; Jiskoot, W. PLGA particulate delivery systems for subunit vaccines: Linking particle properties to immunogenicity. Hum. Vaccin. Immunother. 2016, 12, 1056–1069. [Google Scholar] [CrossRef]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Wang, Y.-T.; Valentine, K.M.; dos Santos Alves, R.P.; Xu, Z.; Regla-Nava, J.A.; Ngono, A.E.; Young, M.P.; Ferreira, L.C.S.; Shresta, S. CD4+ T Cells Cross-Reactive with Dengue and Zika Viruses Protect against Zika Virus Infection. Cell Rep. 2020, 31, 107566. [Google Scholar] [CrossRef]
- Flipse, J.; Smit, J.M. The Complexity of a Dengue Vaccine: A Review of the Human Antibody Response. PLoS Negl. Trop. Dis. 2015, 9, e0003749. [Google Scholar] [CrossRef] [Green Version]
- Bodenheimer, T. Uneasy Alliance—Clinical Investigators and the Pharmaceutical Industry. N. Engl. J. Med. 2000, 342, 1539–1544. [Google Scholar] [CrossRef]
- Chopra, S.S. Industry Funding of Clinical Trials: Benefit or Bias? JAMA J. Am. Med. Assoc. 2003, 290, 113–114. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Antigen | System | Purpose | Soluble | Ref. |
|---|---|---|---|---|
| EDIII | E. coli, pET-15b vector | Structural characterization | No | [34] |
| EDIII | E. coli, pET-20b vector, pelB signal sequence | Thermodynamic stability and molecular design | No | [69] |
| C protein | E. coli strain BL21(DE3) RIL, pET-30a vector, lacking 20 C-terminal amino acids | Structural characterization | Yes | [70] |
| NS2A, NS2B and NS4B | E. coli strain C600, trpE fusion proteins | Identification of the cleavage sites | No | [11] |
| NS3 | E. coli strain W3110, cro full-length NS3 fusion and N-terminal truncated NS3 | Enzymatic activity characterization | No | [71] |
| NS2B/NS3 | E. coli strain M15, pQE30 vector, N-terminal His-tag, hydrophilic core sequence of NS2B linked to NS3 via nonapeptide | Enzymatic activity characterization | Yes | [72] |
| NS5 | HEK293T cells, pIRES/GFP bicistronic mammalian expression vector, His-tagged protein | Phosphorylation characterization | Yes | [73,74] |
| NS1 | E. coli strain Lemo21 (DE3), pBT7-N-His vector | Diagnostics | No | [75] |
| E and NS1 proteins | Vero and Spodoptera frugiperda cells | Glycosylation characterization | Yes | [76] |
| NS1 | E. coli strain BMH17-18, pUR vector, β-galactosidase fusion | Mice immunization: ↑survival after i.c. challenge | No | [35] |
| E protein | Transgenic plant (Nicotiana benthamiana), N-terminal His-tag | Mice immunization: ↑ neutralizing Ab; ↑ IgG avidity; protection after i.c. challenge Monkey immunization: ↓ Viremia; ↑ neutralizing Ab titers; ↑ IFNy | Yes | [77] |
| Target | Strategy | Immune Response | Protection | Refolding | Ref. |
|---|---|---|---|---|---|
| E protein | Fragments of E protein capable of reacting with neutralizing mAbs | ↓Neutralizing Abs | No significant protection | Yes | [99] |
| E protein | Two fragments of E protein Ea and Eb | Ea: ↓Neutralizing Abs Eb: ↑ Neutralizing Abs | Ea–No significant protection Eb–Partial protection | Ea–Yes Eb–No | [100] |
| EDIII | Immunization with Freund’s adjuvant or different charged liposomes in mice | Freund’s adjuvant: ↑ Neutralizing Abs Cationic liposomes: ↑ Neutralizing Abs | Freund’s adjuvant: 60% of protection Cationic liposomes: 80% of protection | No | [102] |
| EDIII | Immunization with Freund’s complete adjuvant Boost with Immunization with Freund’s incomplete adjuvant | ↑ Neutralizing Abs | 62.5% of protection | No | [36] |
| NS1 | Immunization with Freund’s complete adjuvant Boost with Immunization with Freund’s incomplete | ↓Neutralizing Abs | 87.5% of protection | No | [36] |
| Target | Strategy | Immune Response | Protection | Refolding | Ref. |
|---|---|---|---|---|---|
| E protein | Full length E and truncated (80% N-terminal) | Mice: ↑IgG titers. Antibodies recognize WNV-infected cells | Mice: 100% survival. Serum passive transfer 80% survival | No | [123] |
| E antigen in a DNA prime and protein boost | Mice: ↑IgG and neutralizing Abs titers. ↑ CD8 + IFNg+ T cells | Mice: 100% survival. ↓ viremia | Yes | [124] | |
| E ectodomain | E ectodomain plus Matrix-M (saponin) adjuvant | Mice: ↑IgG and neutralizing Ab titers | Mice: 100% survival | Yes | [119] |
| EDIII | EDIII fused with cholera toxin | Mice: ↑IgG, IgM and IgA titers. Complement-mediated killing of serum from immunized mice | ND | No | [125] |
| EDIII | EDIII genetically fused with flagelin (TLR5 agonist) | Mice: ↑IgG and neutralizing Ab titers | Mice: 100% survival | Yes | [122] |
| EDIII | EDIII fused with CD40 ligand | Horses: ↑IgG and neutralizing Ab titers | ND | No | [126] |
| EDIII | EDIII conjugated with VLP of bacteriophage AP205 plus alum | Mice: ↑IgG long-lasting response (>1 year) and neutralizing Ab titers | Mice: 100% survival | Yes | [121] |
| EDIII | EDIII plus CpG | Mice: ↑IgG titers. Abs recognize WNV-infected cells. ↑T-cell proliferation. ↑cytokines (splenocytes) | Mice: ↑survival of mice receiving WNV premixed with antiserum from immunized mice | No | [127] |
| EDIII | Comparison EDIII and soluble full-length E | Mice: partially neutralizing Abs. Different immunogens induce different potencies of neutralizing Abs | ND | No | [128] |
| EDIII | EDIII plus CpG and boosted with oil | Mice: ↑IgG and neutralizing Ab titers | Mice: 80% survival | Yes | [120] |
| EDIII | EDIII boost after E ectodomain DNA prime | Mice: ↑ IgG and neutralizing Ab titers. Moderate IFN-γ | Mice: 100% survival | Yes | [129] |
| EDIII | EDIII plus KFE8 peptide hydrogel | Mice: ↑ IgG and neutralizing Ab titers | Mice: 60% survival | Yes | [130] |
| EDIII | EDIII plus Freund’s adjuvant | Mice: ↑IgG and neutralizing Ab titers | ND | Yes | [131] |
| Target | Strategy | Immune Response | Protection | Refolding | Ref. |
|---|---|---|---|---|---|
| E protein | Fusion with P64k protein from N. meningitidis | Mice and monkeys: ↑ IgG and neutralizing Ab titers after 4 doses | Mice: 50% survival (DENV2 i.c.) Monkeys: reduction of viremia after challenge (DENV2) | No | [168,169,170] |
| E protein | Chimeric protein with 11 peptides from DENV1-4 E protein | Mice: ↑ IgG titers; ↑ CD8 and CD4 T cells; splenocytes (↑IFNy, IL-2,IL-4 and IL-17) | ND | Yes | [171] |
| E protein | DENV2 E protein | Mice: ↑ IgG titers | ND | Yes | [172] |
| E protein | Fragment of DENV1-4 E proteins fused with maltose binding protein (MBP) | Mice and monkey: ↑ IgG and neutralizing Ab titers | Mice: 80% of survival (i.c. challenge); Monkeys: no protection | No | [173,174,175,176] |
| EDIII and C proteins | EDIII fused with C proteins from DENV1-4 | Monkeys and mice: ↑ IgG and neutralizing Ab titers, ↑ B cells antigen-specific; ↑ IFNy (splenocytes or PBMC); ↑ CD8 e CD8 T cells (IFNy) | Mice: ↑ survival (DENV1-4) Monkeys: ↓ viremia and ↑ survival against DENV1-4 | Yes | [177,178,179,180,181,182,183] |
| EDIII protein | DENV1-4 EDIII protein fused to fliC (S. typhimurium) | Mice: ↑ IgG and neutralizing Ab titers (heterologous prime-boost with LATV vaccine); ↓ADE effect | Mice: ↑ survival (DENV1-4) | No | [184] |
| EDIII protein | DENV1-4 EDIII in tandem (B1234 protein) | Mice: ↑ IgG and neutralizing Ab titers | Mice: ↑ survival (DENV1-4) | Yes | [185] |
| EDIII protein | DENV2 EDIII fused with pIII coat protein | Mice: ↑ IgG and neutralizing Ab titers | Mice: no protection; ↑ ADE | Yes | [186] |
| EDIII protein | DENV2 EDIII with different adjuvants (LT1, LTB and Alum) | Mice: ↑ IgG and neutralizing Ab titers | ND | Yes | [187] |
| EDIII protein | DENV2 EDIII | Mice: ↑ IgG and neutralizing Ab titers | Mice: ↑ survival (DENV2) | Yes | [188] |
| EDIII protein | DENV1-4 EDIII fused with lipid signal peptide of the lipoprotein Ag473 | Mice: ↑ IgG and neutralizing Ab titers; ↑ IgG avidity; ↑ neutralization (PRNT); ↓ ADE in vitro | Mice: ↓ viremia | Yes | [189,190,191,192] |
| EDIII protein | DENV1 EDIII with different adjuvants (PELC and CpG) | Mice: ↑ IgG and neutralizing Ab titers; ↑ IFNy (ELISPOT) | ND | No | [193] |
| EDIII protein | DENV1-2 EDIII or DENV3-4 EDIII in tandem | Mice: ↑ IgG and neutralizing Ab titers | Mice: ↑ survival (DENV1-4) | Yes | [194,195] |
| EDIII protein | DENV2 EDIII with Freund’s adjuvant | Mice: ↑ IgG and neutralizing Ab titers | Mice: ↑ survival (DENV2) | Yes | [196] |
| EDIII protein | DENV3 EDIII consensus | Mice: ↑ IgG titers; ↑ proliferation of splenocytes; ↑ IFNy and IL-4 (splenocytes) | ND | No | [197] |
| EDIII protein | DENV1-4 EDIII consensus | Mice: ↑ IgG and neutralizing (DENV1-4) Ab titers | ND | No | [198] |
| EDIII protein | DENV (1,2) EDIII protein | Mice: ↑ IgG titers | ND | Yes | [199] |
| EDIII protein | DENV1-4 EDIII | Mice: ↑ IgG and neutralizing Ab titers; ↑ CD4 and CD8 T cells (producing IFNy and IL-2); ↑ IFNy, IL-2, IL-12p40 (splenocytes) | ND | Yes | [200] |
| EDIII protein | DENV2 EDIII in Chimeric VLP (HBcAg-EDIII-2) | Mice: ↑ IgG and neutralizing Ab titers | ND | Yes | [201] |
| EDII protein | Fubc protein (peptides from fusion and bc loop regions of EDII fused by link sequence) | Mice: ↑ IgG titers | ND | Yes | [202] |
| NS1 protein | DENV2 NS1 with LTG33D, Alum or Freund’s adjuvant | Mice: ↑ IgG titers | Mice: ↑ survival (DENV2) | Yes | [37,203] |
| NS5 protein | DENV2 NS5 | Mice: ↑ IgG titers; ↑ IFNy and TNFα (splenocytes) | Mice: ↑ survival (DENV2) | No | [38] |
| NS3 protein | DENV2 NS3 protein | Mice: ↑ IgG titers; ↑ IFNy (splenocytes) | ND | Yes | [39] |
| NS3/NS1 | DJ NS1 chimera (DENV2 and JEV) and NS3 | Mice: ↑ IgG titers;↑ CD4 and CD8 T cells; ↑ CTL responses against NS3 | Mice: ↓ viremia; ↓ soluble NS1 levels; ↓ mouse tail bleeding time, and vascular leakage at skin injection sites | Yes | [40] |
| Clinical Trials | Status | Inactivated Virus | Attenuated Virus | DNA-Based | RNA-Based | Viral Vector-Based | Total |
|---|---|---|---|---|---|---|---|
| Early phase I or phase I | Ongoing | NCT03008122 NCT03343626 | - | - | NCT04064905 | NCT04033068 NCT04440774 NCT04015648 | 6 |
| Completed | NCT02937233 NCT02952833 NCT02963909 NCT03425149 | NCT03611946 | NCT02840487 NCT02996461 | NCT03014089 | NCT02996890 | 9 | |
| Phase II | Completed | - | - | NCT03110770 | - | - | 1 |
| Target | Strategy | Immune Response | Protection | Refolding | Ref. |
|---|---|---|---|---|---|
| E protein | 90% of the N-terminal whole E protein + alum adjuvant | Mice: ↑IgG and neutralizing Ab titers | Neonatal mice: full protection Pregnant mice: fetus and offspring protection from ZIKV-induced microcephaly | Yes | [216] |
| E protein | 80% of the N-terminal whole E protein + alum adjuvant | Mice: ↑IgG and neutralizing Ab titers; ↑ ZIKV specific T cells; IFNy | Mice: ↑survival | Yes | [218] |
| E protein | Consensus E protein lacking the whole stem and trans membrane regions + poly (I:C) or CpG ODN adjuvants | Mice: ↑IgG titer; ↑ ZIKV specific T cells; IFNy | ND | Yes | [219] |
| EDIII protein | Full EDIII sequence + TMG or alum adjuvants | Mice: ↑IgG and neutralizing Ab titers; ↑IFNy, IL-4 and IL-6 | ND | Yes | [220] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araujo, S.C.; Pereira, L.R.; Alves, R.P.S.; Andreata-Santos, R.; Kanno, A.I.; Ferreira, L.C.S.; Gonçalves, V.M. Anti-Flavivirus Vaccines: Review of the Present Situation and Perspectives of Subunit Vaccines Produced in Escherichia coli. Vaccines 2020, 8, 492. https://doi.org/10.3390/vaccines8030492
Araujo SC, Pereira LR, Alves RPS, Andreata-Santos R, Kanno AI, Ferreira LCS, Gonçalves VM. Anti-Flavivirus Vaccines: Review of the Present Situation and Perspectives of Subunit Vaccines Produced in Escherichia coli. Vaccines. 2020; 8(3):492. https://doi.org/10.3390/vaccines8030492
Chicago/Turabian StyleAraujo, Sergio C., Lennon R. Pereira, Rubens P. S. Alves, Robert Andreata-Santos, Alex I. Kanno, Luis Carlos S. Ferreira, and Viviane M. Gonçalves. 2020. "Anti-Flavivirus Vaccines: Review of the Present Situation and Perspectives of Subunit Vaccines Produced in Escherichia coli" Vaccines 8, no. 3: 492. https://doi.org/10.3390/vaccines8030492
APA StyleAraujo, S. C., Pereira, L. R., Alves, R. P. S., Andreata-Santos, R., Kanno, A. I., Ferreira, L. C. S., & Gonçalves, V. M. (2020). Anti-Flavivirus Vaccines: Review of the Present Situation and Perspectives of Subunit Vaccines Produced in Escherichia coli. Vaccines, 8(3), 492. https://doi.org/10.3390/vaccines8030492