
The Natural Oligoribonucleotides Functionalized by D-Mannitol Affected Interactions of Hemagglutinin with Glycan Receptor Indicating Anti-Influenza Activity

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
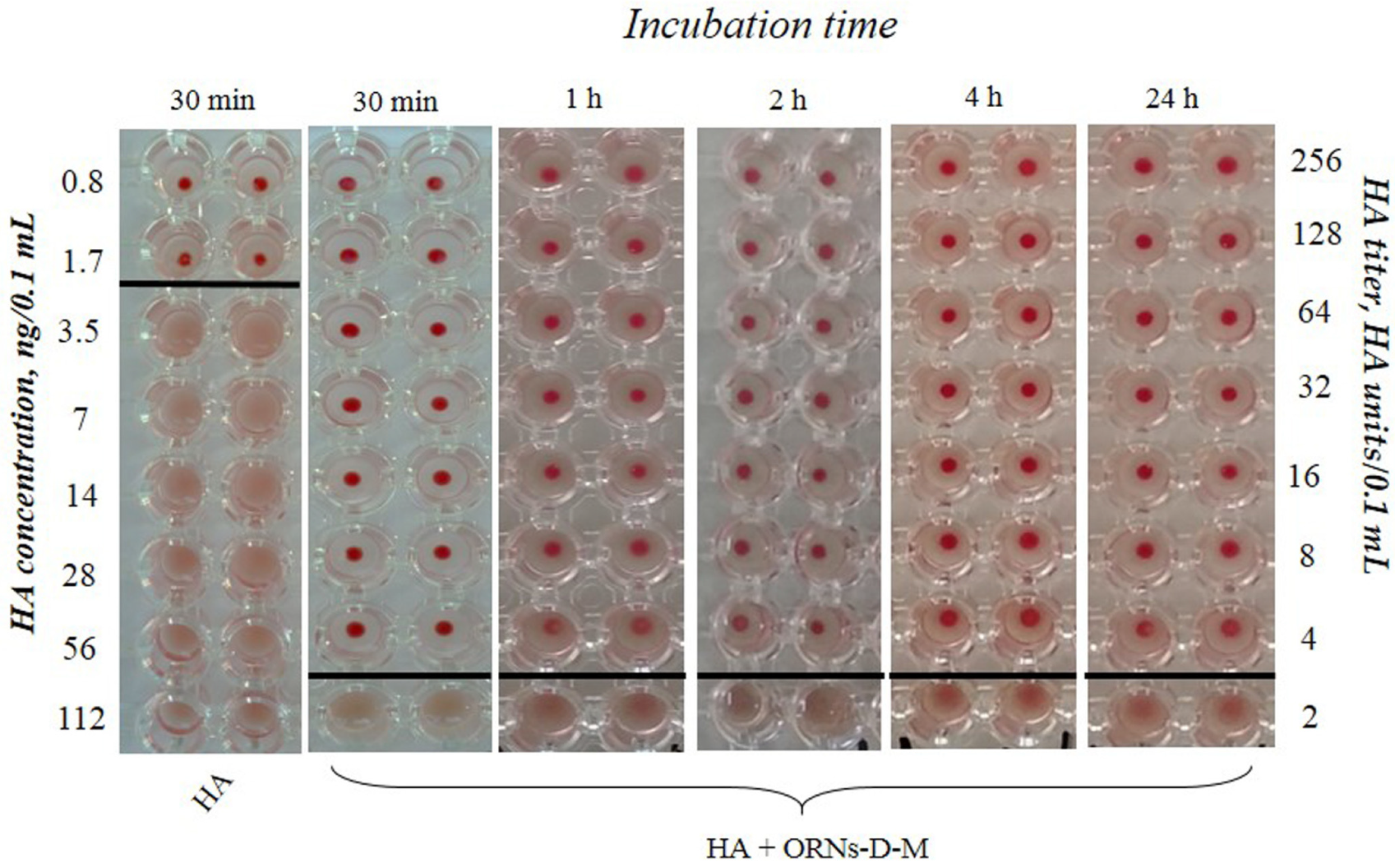
2.2. Hemagglutination Analysis
2.3. Fluorescence Measurements
2.4. Docking of the ORNs and ORNs-D-M to Influenza Hemagglutinin
3. Results and Discussion
3.1. Analysis of the Interaction of ORNs and ORNs-D-M with HA
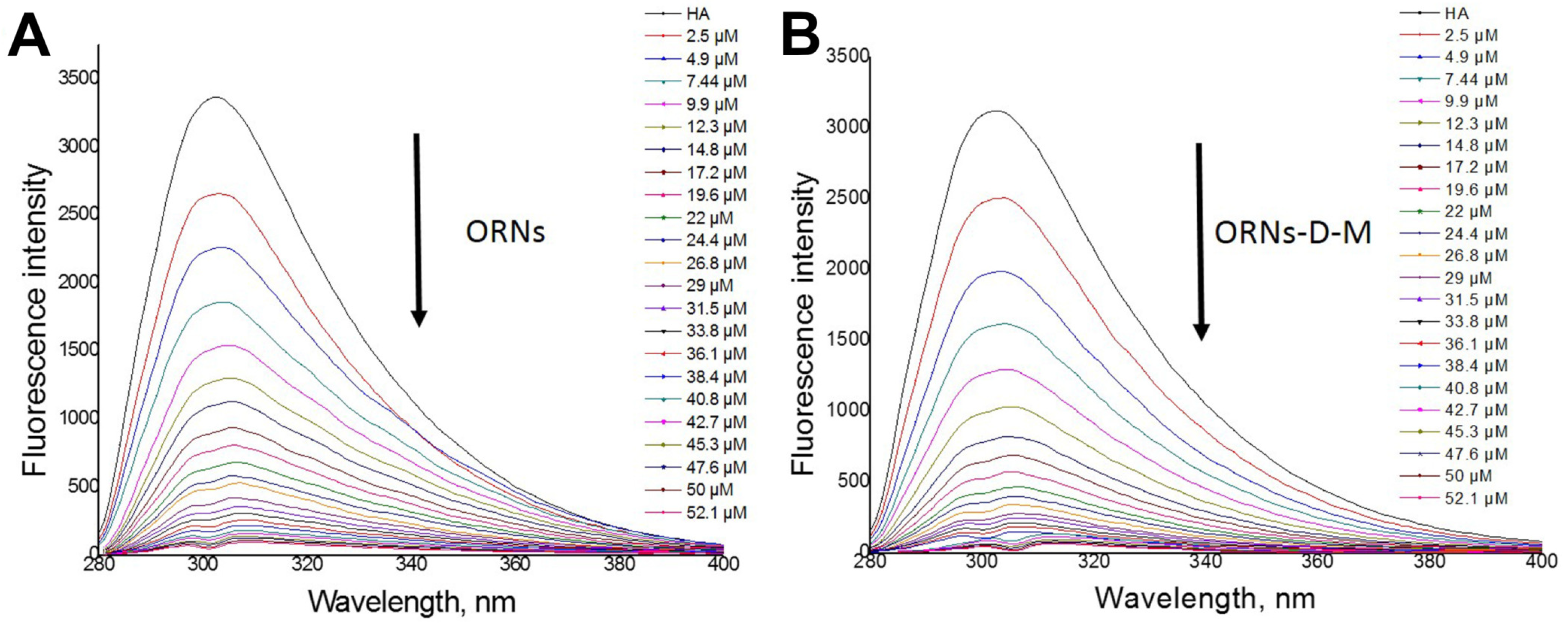
3.2. HA Fluorescence Quenching by the ORNs and ORNs-D-M
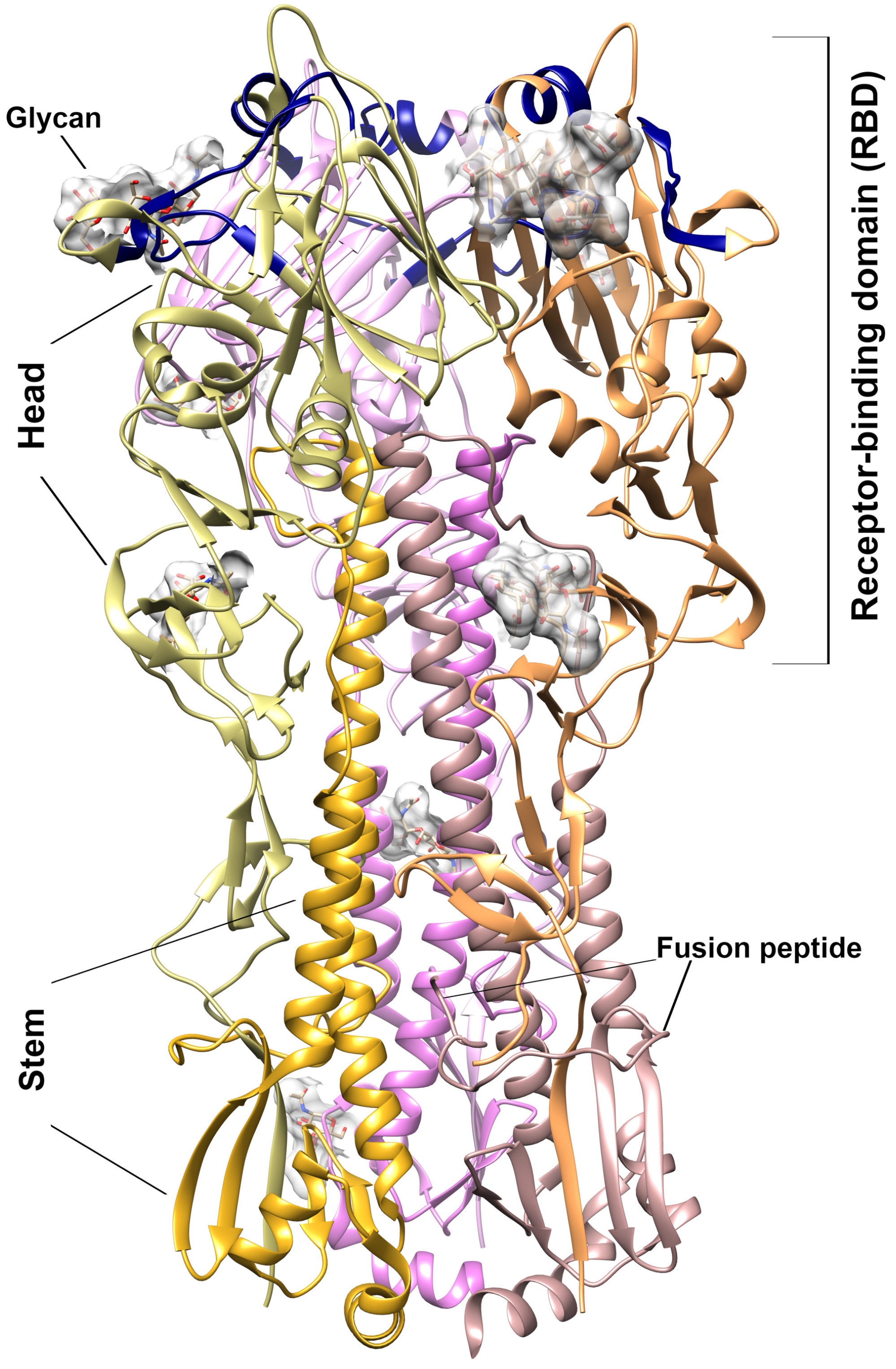
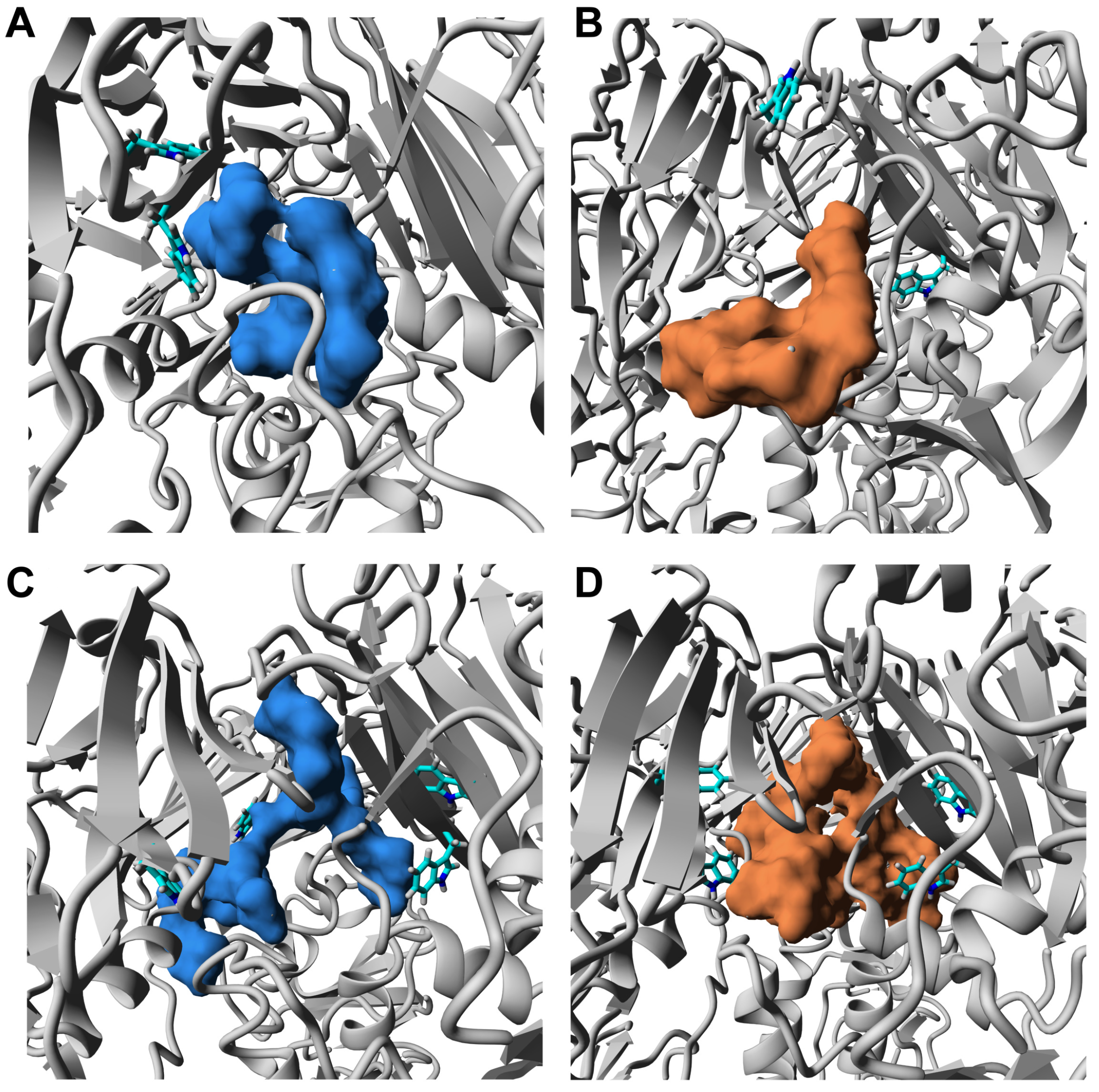
3.3. ORNs and ORNs-D-M Bind to the HA Protein Mostly in RBD Subdomain
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| HA | hemagluttinin protein |
| HA3 | H3 HA strain of the influenza hemagluttinin from A/Hong Kong/1/1968 (H3N2) |
| RBD | receptor-binding domain |
| ORNs | oligoribonucleotides |
| A3-ORNs | ligand comprized sequence of three oligoribonucleotides |
| A6-ORNs | ligand comprized sequence of six oligoribonucleotides |
| ORNs-D-M | oligoribonucleotides functionalized with D-mannitol |
| A3-ORNs-D-M | ligand comprized sequence of three oligoribonucleotides functionalized |
| with D-mannitol | |
| A6-ORNs-D-M | ligand comprized sequence of six oligoribonucleotides functionalized |
| with D-mannitol |
References
- Salzberg, S. The contents of the syringe. Nature 2008, 454, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Davlin, S.L.; Blanton, L.; Kniss, K.; Mustaquim, D.; Smith, S.; Kramer, N.; Cohen, J.; Cummings, C.N.; Garg, S.; Flannery, B.; et al. Influenza activity–United States, 2015–16 season and composition of the 2016–17 influenza vaccine. Morb. Mortal. Wkly. Rep. 2016, 65, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Hensley, S.E. Challenges of selecting seasonal influenza vaccine strains for humans with diverse pre-exposure histories. Curr. Opin. Virol. 2014, 8, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Moscona, A. Global transmission of oseltamivir-resistant influenza. N. Engl. J. Med. 2009, 360, 953–956. [Google Scholar] [CrossRef] [Green Version]
- Sheu, T.G.; Fry, A.M.; Garten, R.J.; Deyde, V.M.; Shwe, T.; Bullion, L.; Peebles, P.J.; Li, Y.; Klimov, A.I.; Gubareva, L.V. Dual resistance to adamantanes and oseltamivir among seasonal influenza A (H1N1) viruses: 2008–2010. J. Infect. Dis. 2011, 203, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bright, R.A.; Shay, D.K.; Shu, B.; Cox, N.J.; Klimov, A.I. Adamantane resistance among influenza A viruses isolated early during the 2005–2006 influenza season in the United States. JAMA 2006, 295, 891–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobasa, D.; Takada, A.; Shinya, K.; Hatta, M.; Halfmann, P.; Theriault, S.; Suzuki, H.; Nishimura, H.; Mitamura, K.; Sugaya, N.; et al. Enhanced virulence of influenza A viruses with the haemagglutinin of the 1918 pandemic virus. Nature 2004, 431, 703–707. [Google Scholar] [CrossRef]
- de Graaf, M.; Fouchier, R.A. Role of receptor binding specificity in influenza A virus transmission and pathogenesis. EMBO J. 2014, 33, 823–841. [Google Scholar] [CrossRef] [Green Version]
- Worch, R. Structural biology of the influenza virus fusion peptide. Acta Biochim. Pol. 2014, 61. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.J.; Kerry, P.S.; Stevens, D.J.; Steinhauer, D.A.; Martin, S.R.; Gamblin, S.J.; Skehel, J.J. Structure of influenza hemagglutinin in complex with an inhibitor of membrane fusion. Proc. Natl. Acad. Sci. USA 2008, 105, 17736–17741. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Zhang, X.; Liu, S. Novel hemagglutinin-based influenza virus inhibitors. J. Thorac. Dis. 2013, 5, S149. [Google Scholar]
- Tkachuk, Z. Multiantivirus Compound, Composition and Method for Treatment of Virus Diseases. U.S. Patent 8,420,617, 16 April 2013. [Google Scholar]
- Melnichuk, N.; Zarubaev, V.; Iosyk, I.; Andreychyn, M.; Semernikova, L.; Tkachuk, Z. Pre-clinical and clinical efficiency of complexes of oligoribonucleotides with D-Mannitol against respiratory viruses. Pharmaceutics 2018, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Tkachuk, Z.; Rybalko, S.; LD, Z.; Starostyla, D. Antiinfluenzal activity of drug Nuclex. Rep. Natl. Acad. Sci. Ukr. 2010, 3, 191–196. [Google Scholar]
- Melnichuk, N.; Kashuba, V.; Rybalko, S.; Tkachuk, Z. Complexes of oligoribonucleotides with D-Mannitol modulate the innate immune response to influenza a virus H1N1 (A/FM/1/47) in vivo. Pharmaceuticals 2018, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnichuk, N.; Semernikova, L.; Tkachuk, Z. Complexes of Oligoribonucleotides with D-mannitol inhibit hemagglutinin–glycan interaction and suppress influenza A virus H1N1 (A/FM/1/47) infectivity in vitro. Pharmaceuticals 2017, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Melnichuk, N.; Tkachuk, Z. Inhibition of hemagglutinin-glycan interaction by complexes of oligoribonucleotides with D-mannitol. Rep. Natl. Acad. Sci. Ukr. 2018, 3, 92–99. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakowicz, J.R. Introduction to fluorescence. In Principles of Fluorescence Spectroscopy; Springer: Berlin/Heidelberg, Germany, 1999; pp. 1–23. [Google Scholar]
- Favicchio, R.; Dragan, A.I.; Kneale, G.G.; Read, C.M. Fluorescence spectroscopy and anisotropy in the analysis of DNA-protein interactions. In DNA–Protein Interactions; Springer: Berlin/Heidelberg, Germany, 2009; pp. 589–611. [Google Scholar]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the precision of comparative models with YASARA NOVA—A self-parameterizing force field. Proteins Struct. Funct. Bioinform. 2002, 47, 393–402. [Google Scholar] [CrossRef]
- Kadam, R.U.; Wilson, I.A. Structural basis of influenza virus fusion inhibition by the antiviral drug Arbidol. Proc. Natl. Acad. Sci. USA 2017, 114, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, A.; Wong, Y.H.; Korsholm, S.S.; Bahring, N.H.B.; Bobone, S.; Tayyab, S.; Van De Weert, M.; Stella, L. On the purported “backbone fluorescence” in protein three-dimensional fluorescence spectra. RSC Adv. 2016, 6, 112870–112876. [Google Scholar] [CrossRef] [Green Version]
- Wani, T.A.; Bakheit, A.H.; Ansari, M.N.; Al-Majed, A.R.A.; Al-Qahtani, B.M.; Zargar, S. Spectroscopic and molecular modeling studies of binding interaction between bovine serum albumin and roflumilast. Drug Des. Dev. Ther. 2018, 12, 2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Eddy, N.R.; Noel, J.K.; Whitford, P.C.; Wang, Q.; Ma, J.; Onuchic, J.N. Order and disorder control the functional rearrangement of influenza hemagglutinin. Proc. Natl. Acad. Sci. USA 2014, 111, 12049–12054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bangaru, S.; Lang, S.; Schotsaert, M.; Vanderven, H.A.; Zhu, X.; Kose, N.; Bombardi, R.; Finn, J.A.; Kent, S.J.; Gilchuk, P.; et al. A site of vulnerability on the influenza virus hemagglutinin head domain trimer interface. Cell 2019, 177, 1136–1152. [Google Scholar] [CrossRef]
- Watanabe, A.; McCarthy, K.R.; Kuraoka, M.; Schmidt, A.G.; Adachi, Y.; Onodera, T.; Tonouchi, K.; Caradonna, T.M.; Bajic, G.; Song, S.; et al. Antibodies to a conserved influenza head interface epitope protect by an IgG subtype-dependent mechanism. Cell 2019, 177, 1124–1135. [Google Scholar] [CrossRef] [PubMed]
- Sawicka, R.; Siedlecki, P.; Kalenik, B.; Radomski, J.P.; Sączyńska, V.; Porębska, A.; Szewczyk, B.; Sirko, A.; Góra-Sochacka, A. Characterization of mAb6-9-1 monoclonal antibody against hemagglutinin of avian influenza virus H5N1 and its engineered derivative, single-chain variable fragment antibody. Acta Biochim. Pol. 2017, 64, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, R.A. The influenza virus RNA segments and their encoded proteins. In Genetics of Influenza Viruses; Springer: Berlin/Heidelberg, Germany, 1983; pp. 21–69. [Google Scholar]
- Wilson, I.A.; Skehel, J.J.; Wiley, D. Structure of the haemagglutinin membrane glycoprotein of influenza virus at 3 Å resolution. Nature 1981, 289, 366–373. [Google Scholar] [CrossRef]
- de Vries, R.P.; de Vries, E.; Bosch, B.J.; de Groot, R.J.; Rottier, P.J.; de Haan, C.A. The influenza A virus hemagglutinin glycosylation state affects receptor-binding specificity. Virology 2010, 403, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Sztain, T.; Ahn, S.H.; Bogetti, A.T.; Casalino, L.; Goldsmith, J.A.; Seitz, E.; McCool, R.S.; Kearns, F.L.; Acosta-Reyes, F.; Maji, S.; et al. A glycan gate controls opening of the SARS-CoV-2 spike protein. Nat. Chem. 2021, 1–6. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tkachuk, Z.; Melnichuk, N.; Nikolaiev, R.O.; Szutkowski, K.; Zhukov, I. The Natural Oligoribonucleotides Functionalized by D-Mannitol Affected Interactions of Hemagglutinin with Glycan Receptor Indicating Anti-Influenza Activity. Membranes 2021, 11, 757. https://doi.org/10.3390/membranes11100757
Tkachuk Z, Melnichuk N, Nikolaiev RO, Szutkowski K, Zhukov I. The Natural Oligoribonucleotides Functionalized by D-Mannitol Affected Interactions of Hemagglutinin with Glycan Receptor Indicating Anti-Influenza Activity. Membranes. 2021; 11(10):757. https://doi.org/10.3390/membranes11100757
Chicago/Turabian StyleTkachuk, Zenoviy, Nataliia Melnichuk, Roman O. Nikolaiev, Kosma Szutkowski, and Igor Zhukov. 2021. "The Natural Oligoribonucleotides Functionalized by D-Mannitol Affected Interactions of Hemagglutinin with Glycan Receptor Indicating Anti-Influenza Activity" Membranes 11, no. 10: 757. https://doi.org/10.3390/membranes11100757
APA StyleTkachuk, Z., Melnichuk, N., Nikolaiev, R. O., Szutkowski, K., & Zhukov, I. (2021). The Natural Oligoribonucleotides Functionalized by D-Mannitol Affected Interactions of Hemagglutinin with Glycan Receptor Indicating Anti-Influenza Activity. Membranes, 11(10), 757. https://doi.org/10.3390/membranes11100757