
Entamoeba histolytica: Proteomics Bioinformatics Reveal Predictive Functions and Protein–Protein Interactions of Differentially Abundant Membrane and Cytosolic Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. PERSEUS Software Platform Analysis
2.2. Determination of Transmembrane and Signal Peptide Domains
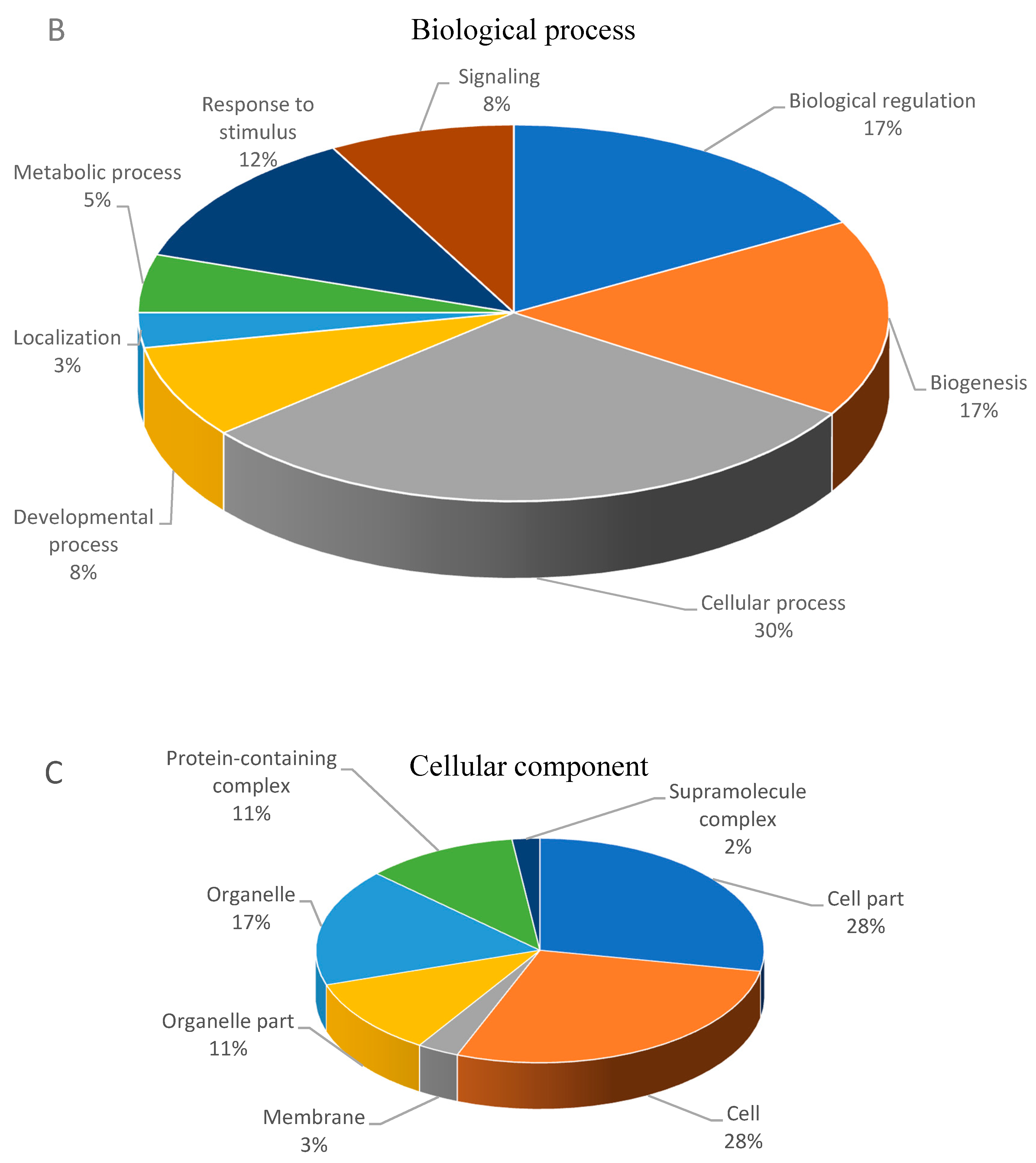
2.3. Gene Ontology
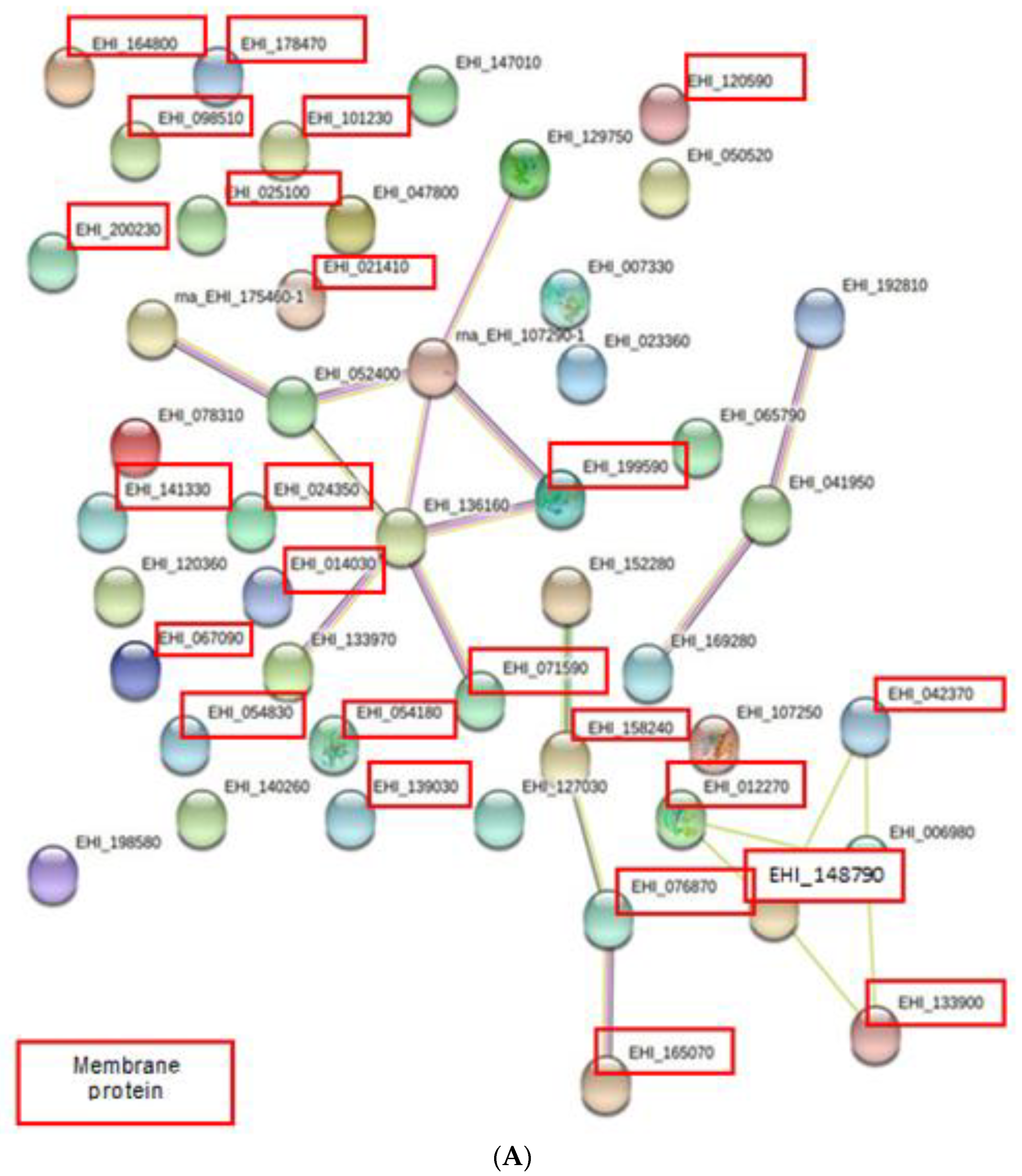
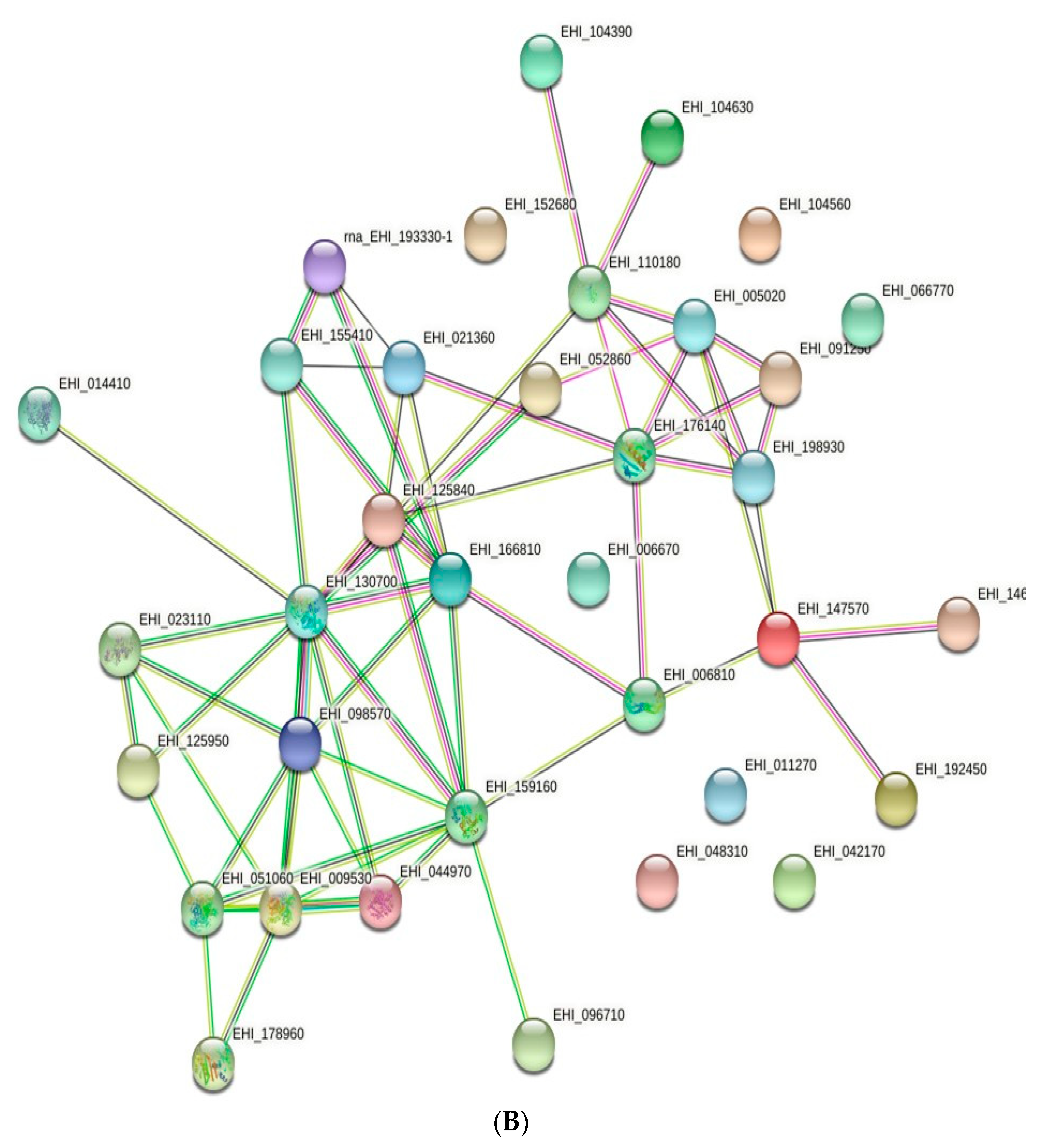
2.4. Protein–Protein Interaction Network Prediction
3. Results
3.1. Differential Abundant Proteins
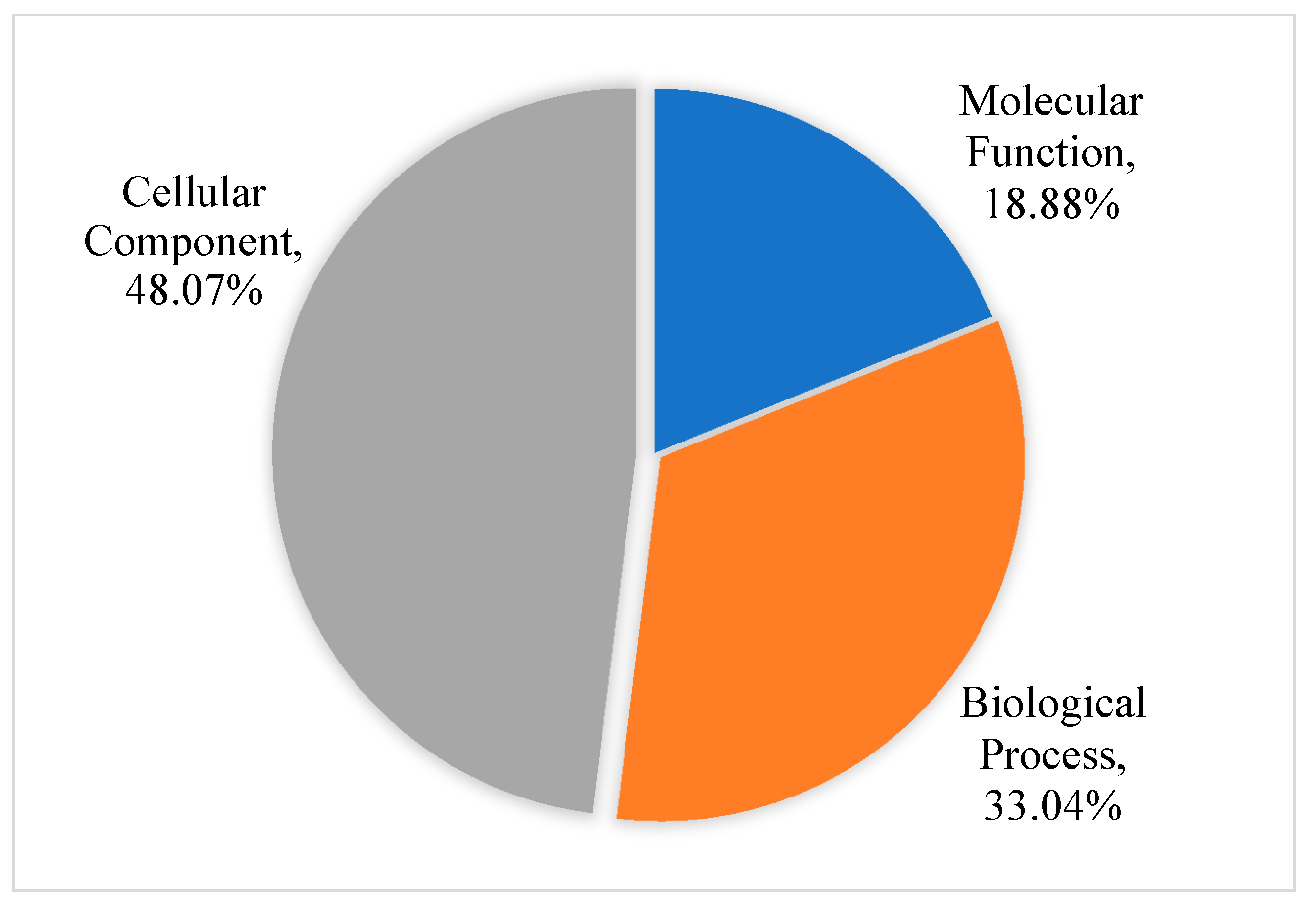
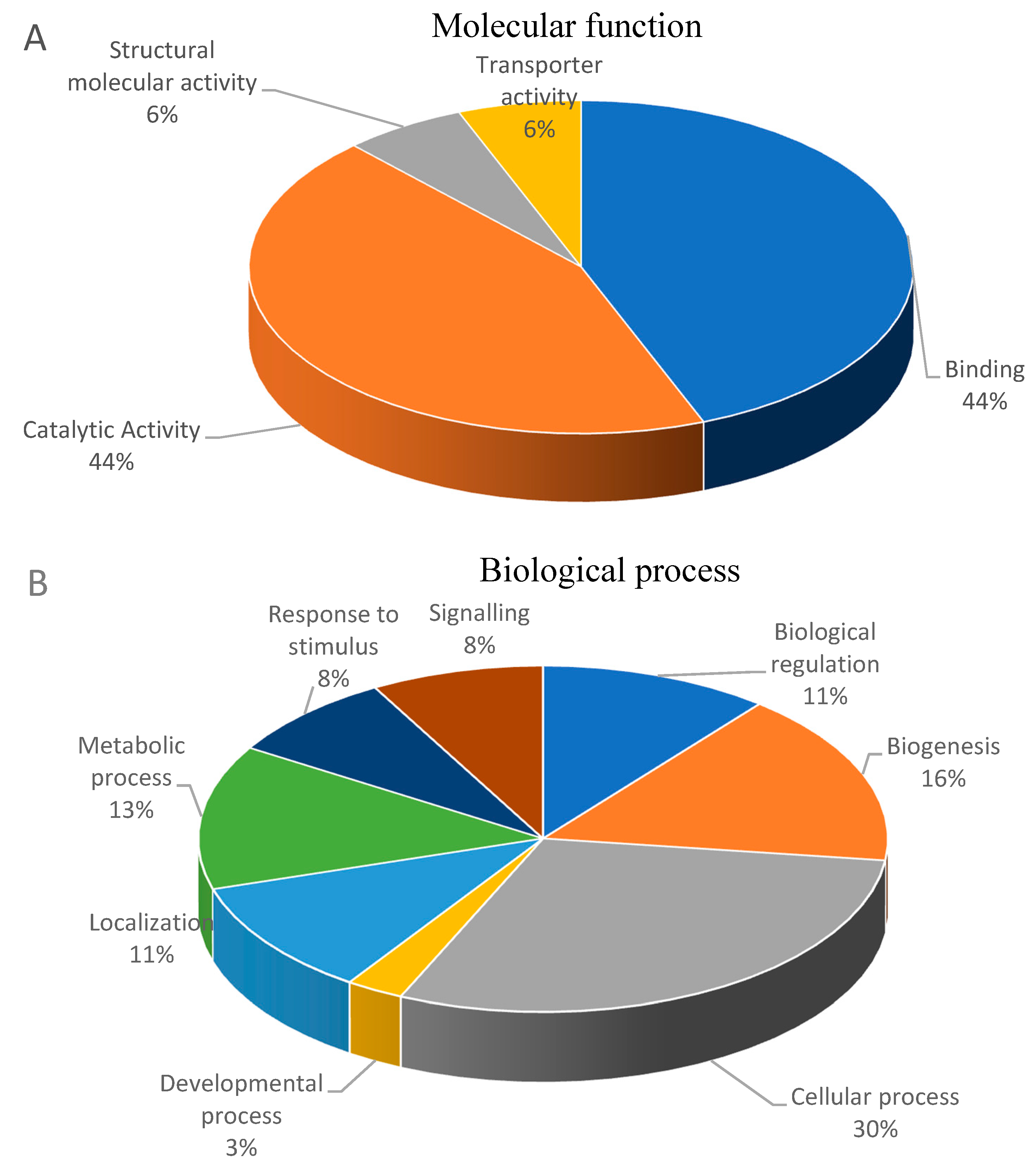
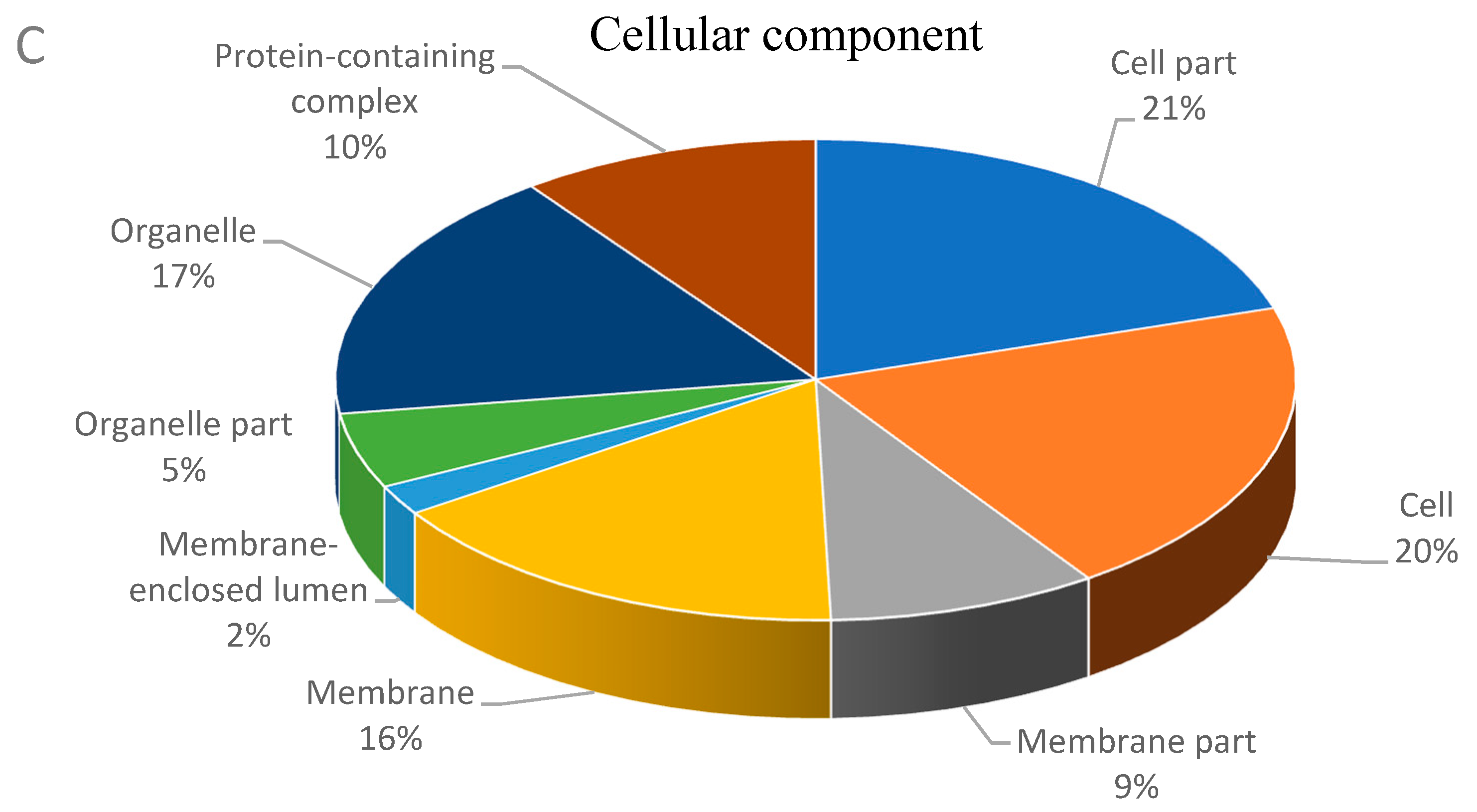
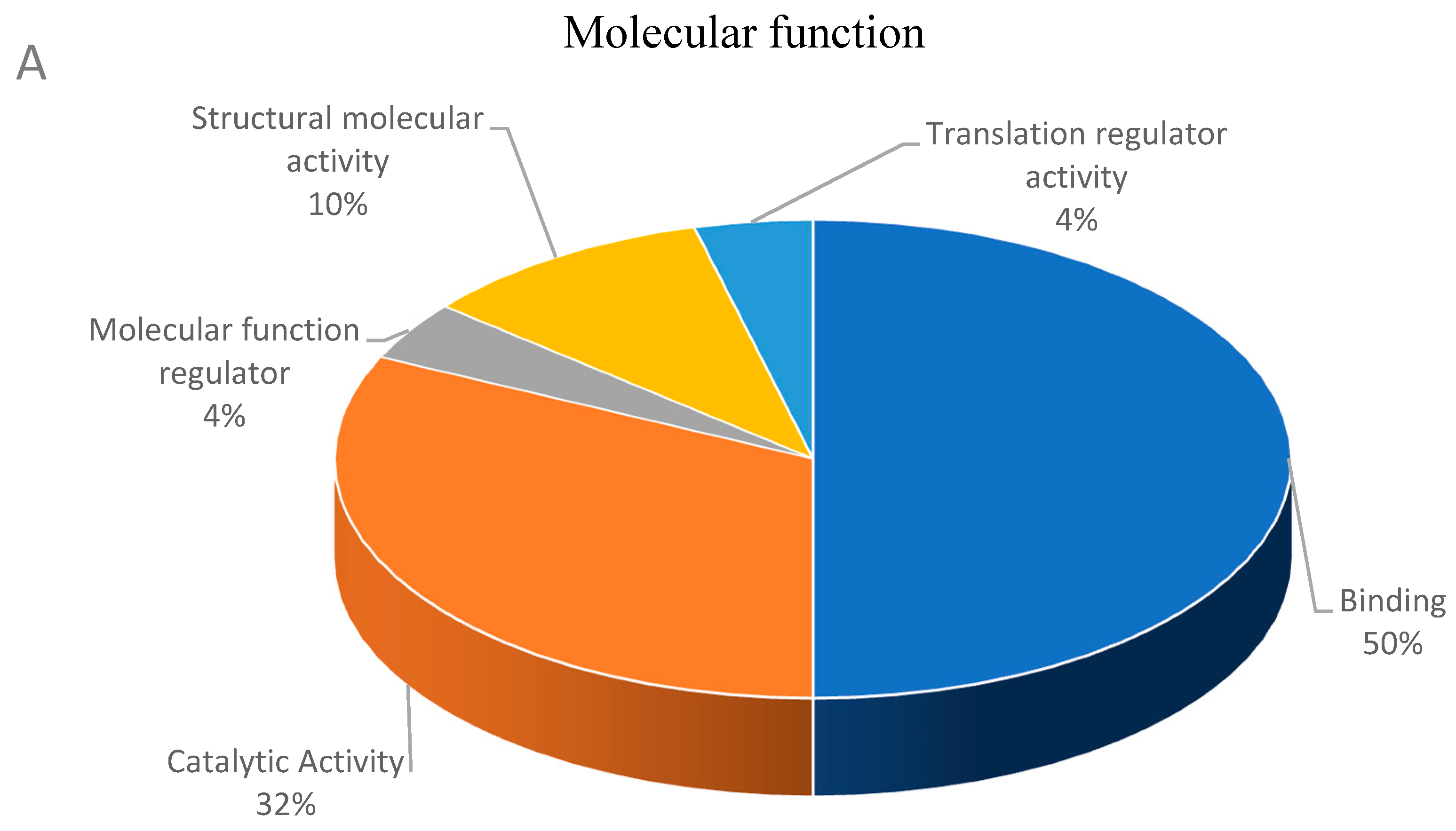
3.2. Functional Classification of Differential Abundant Proteins.
3.3. Protein–Protein Interaction Network Prediction.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biller, L.; Matthiesen, J.; Kühne, V.; Lotter, H.; Handal, G.; Nozaki, T.; Saito-Nakano, Y.; Schümann, M.; Roeder, T.; Tannich, E.; et al. The Cell Surface Proteome of Entamoeba histolytica. Mol. Cell. Proteom. 2014, 13, 132–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laura, M.; Upinder, S. Entamoeba histolytica: A snapshot of current research and methods for genetic analysis. Curr. Opin. Microbiol. 2012, 15, 469–475. [Google Scholar]
- Ibne Karim, M.A.; Rashidul, H.; Abdullah, S.; Mamun, K.; Nicholas, E.S.; Sean, A.G.; Gerard, A.C.; William, A.P., Jr. Proteomic analysis of the cyst stage of Entamoeba histolytica. PLoS Negl. Trop. Dis. 2012, 6, e1643. [Google Scholar]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsirigos, K. Bioinformatics Methods for Topology Prediction of Membrane Proteins. Ph.D. Thesis, Department of Biochemistry and Biophysics, Stockholm University, Stockholm, Sweden, 2017. [Google Scholar]
- Zhang, Q.C.; Petrey, D.; Garzón, J.I.; Deng, L.; Honig, B. PrePPI: A structure-informed database of protein–protein interactions. Nucleic Acids Res. 2012, 41, D828–D833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujang, J.; Sani, A.A.A.; Lim, B.H.; Noordin, R.; Othman, N. Analysis of Entamoeba histolytica Membrane Proteome Using Three Extraction Methods. Proteomics 2018, 18, e1700397. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, R.C.; Singh, U. Identification of Differentially Expressed Genes in Virulent and Nonvirulent Entamoeba Species: Potential Implications for Amebic Pathogenesis. Infect. Immun. 2006, 74, 340–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, Y.L.; Olivos-García, A.; Lim, T.K.; Noordin, R.; Lin, Q.; Othman, N. Entamoeba histolytica: Quantitative Proteomics Analysis Reveals Putative Virulence-Associated Differentially Abundant Membrane Proteins. Am. J. Trop. Med. Hyg. 2018, 99, 1518–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soid-Raggi, L.G.; Torres-Márquez, M.E.; Meza, I. Entamoeba histolytica: Identification of functional G(s) and G(i) proteins as possible signal transduction elements in the interaction of trophozoites with fibronectin. Exp. Parasitol. 1998, 90, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Ximénez, C.; González, E.; Nieves, M.; Magaña, U.; Morán, P.; Gudiño-Zayas, M. Differential expression of pathogenic genes of E. histolytica versus E. dispar in a model of infection using human liver tissue explants. PLoS ONE 2017, 12, e0181962. [Google Scholar]
- Schlosser, S.; Leitschm, D.; Duchêne, M. Entamoeba histolytica: Identification of thioredoxin-targeted proteins and analysis of serine acetyltransferase-1 as a prototype example. Biochem. J. 2013, 451, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Isenberg, G.; Aebi, U.; Pollard, T.D. An action-binding protein from Acanthamoeba regulates actin filament polymerization and interactions. Nature 1980, 288, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Salgado, M.; Villagómez-Castro, J.C.; Rocha-Rodríguez, R.; Sabanero-Lopez, M.; Ramos, M.A.; Alagón, A.; López-Romero, E.; Sánchez-López, R. Entamoeba histolytica: Biochemical and molecular insights into the activities within microsomal fractions. Exp. Parasitol. 2005, 110, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, E.; Rico, G.; Mendoza, G.; Ramos, F.; Garcıa, G.; Moran, P.; Valadez, A.; Melendro, E.I.; Ximenez, C. Calreticu-lin-like molecule in trophozoites of Entamoeba histolytica HM1: IMSS (Swissprot: Accession P83003). Am. J. Trop. Med. Hyg. 2002, 67, 636–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction Method | Increased | Decreased |
|---|---|---|
| Fc > 2-Fold; q < 0.05 | Fc > 2-Fold; q < 0.05 | |
| Conventional Method | 19 | 23 |
| ProteoExtract Kit | 4 | 2 |
| ProteoPrep Kit | 31 | 20 |
| TotalL | 54 | 45 |
| TM and/or Signal Peptide | Fold Change | Mapped ID | Gene Name |
|---|---|---|---|
| - | 2.0141 | EHI_078310 | Uncharacterized protein |
| - | 2.3102 | EHI_129750 | Rho family GTPase |
| - | 2.0333 | EHI_198580 | Uncharacterized protein |
| Yes | 2.4443 | EHI_165070 | Estradiol 17-beta-dehydrogenase putative |
| - | 2.6115 | EHI_023360 | Alpha-amylase family protein |
| - | 3.6739 | EHI_065790 | Rab family GTPase (Fragment) |
| - | 2.3510 | EHI_152280 | Serine palmitoyltransferase putative |
| Yes | 2.6371 | EHI_199590 | 70 kDa heat shock protein putative |
| - | 2.2277 | EHI_120360 | Grainin putative |
| - | 2.1083 | EHI_006980 | Gal galnac lectin subunit igl1 |
| - | 2.5152 | EHI_102170 | Elongation factor 1-alpha |
| Yes | 2.0661 | EHI_024350 | Uncharacterized protein |
| - | 2.0284 | EHI_107290 | Actin |
| - | 2.6989 | EHI_047800 | Uncharacterized protein |
| Yes | 2.5246 | EHI_076870 | 3-oxo 5 alpha-steroid 4-dehydrogenase domain-containing |
| Yes | 2.9713 | EHI_054830 | Calcium-transporting ATPase |
| - | 4.4102 | EHI_050520 | Uncharacterized protein |
| - | 2.8741 | EHI_169280 | EhRab7E protein |
| Yes | 3.3411 | EHI_014030 | NAD(P) transhydrogenase subunit alpha, putative |
| Yes | 2.6606 | EHI_025100 | G protein-coupled receptor 1 |
| Yes | 4.1994 | EHI_012270 | 170 kDa surface lectin |
| Yes | 2.3614 | EHI_141330 | Uncharacterized protein |
| Yes | 2.5185 | EHI_120590 | Uncharacterized protein |
| - | 2.0883 | EHI_041950 | Vacuolar protein sorting 35, putative |
| Yes | 2.8047 | EHI_164800 | Cysteine protease binding protein family 1 |
| - | 3.411 | EHI_136160 | Calreticulin putative |
| Yes | 3.1331 | EHI_098510 | Uncharacterized protein |
| - | 4.4288 | EHI_192810 | small GTPase Rab7A |
| - | 2.9434 | EHI_147010 | Long-chain-fatty-acid—CoA ligase, putative |
| Yes | 2.3719 | EHI_071590 | Protein disulfide isomerase putative |
| - | 2.2623 | EHI_133970 | Thioredoxin putative |
| Yes | 2.8980 | EHI_158240 | 3-ketoacyl-CoA synthase |
| Yes | 2.2738 | EHI_139030 | EhSyntaxin 1A (Fragment) |
| - | 2.3774 | EHI_107250 | small GTPase Rab11B |
| - | 3.3584 | EHI_140260 | Copine putative |
| Yes | 2.2996 | EHI_054180 | Protein SEY1 homolog 2 |
| Yes | 3.1250 | EHI_200230 | Cell surface protease gp63 putative |
| - | 2.3851 | EHI_127030 | Uncharacterized protein |
| - | 2.4601 | EHI_007330 | Beta-hexosaminidase |
| Yes | 2.0039 | EHI_067090 | Uncharacterized protein |
| Yes | 2.5342 | EHI_178470 | Cysteine protease binding protein family 6 |
| Yes | 3.1223 | EHI_042370 | Galactose-specific adhesin 170 kDa subunit, putative |
| Yes | 2.7603 | EHI_021410 | EhSyntaxin B |
| Yes | 3.8750 | EHI_133900 | Galactose-inhibitable lectin 170 kDa subunit putative |
| Yes | 3.4012 | EHI_148790 | Gal galNac lectin light subunit |
| - | 2.7452 | EHI_175460 | 60S acidic ribosomal protein P0 |
| Yes | 2.4905 | EHI_101230 | P-glycoprotein 6 |
| - | 2.5152 | EH_052400 | Elongation factor-1-alpha |
| Mapped ID | Fold Change | Gene Name/Symbol | PANTHER (Family/Subfamily) |
|---|---|---|---|
| EHI_130700 | 2.953 | Enolase putative | Enolase (Pthr11902:Sf1) |
| EHI_052860 | 2.9703 | Heat shock protein 70 putative | Ribosome-Associated Molecular Chaperone Ssb1-Related (Pthr19375:Sf395) |
| EHI_104390 | 2.6725 | Actin binding protein putative | - |
| EHI_042170 | 3.2691 | Aminoacyl-histidine dipeptidase putative | Cytosol Non-Specific Dipeptidase (Pthr43501:Sf1) |
| EHI_048310 | 2.4029 | EhSec24C | Secretory 24cd, Isoform C (Pthr13803:Sf38) |
| EHI_023110 | 3.1633 | NADP-dependent Alcohol dehydrogenase | Dehydrogenase 1, Putative-Related (Pthr42813:Sf4) |
| EHI_125840 | 3.0822 | Peptidyl-prolyl cis-trans isomerase | Peptidyl-Prolyl Cis-Trans Isomerase D-Related (Pthr11071:Sf380) |
| EHI_166810 | 3.7550/ 2.6014 | Elongation factor 2 | Elongation Factor 2 (Pthr42908:Sf10) |
| EHI_098570 | 3.1517 | Fructose 1 6-bisphosphate aldolase putative | D-Tagatose-1,6-Bisphosphate Aldolase Subunit Gaty-Related (Pthr30304:Sf0) |
| EHI_155410 | 2.8691 | 40S ribosomal protein S11, putative | 40s Ribosomal Protein S11 (Pthr10744:Sf9) |
| EHI_104630 | 2.5185 | Filamin 2 putative | Zgc:100997 (Pthr19961:Sf58) |
| EHI_006670 | 2.5009 | Uncharacterized protein | Expressed Protein (Pthr19308:Sf14) |
| EHI_198930 | 3.7526 | Actin putative | Actin-Related Protein 3b (Pthr11937:Sf31) |
| EHI_021360 | 2.5234 | Uncharacterized protein | Translationally Controlled Tumor Protein (Pthr11991:Sf0) |
| EHI_192450 | 2.0262 | Rho family GTPase | Ras-Related Protein Rac1-Related (Pthr24072:Sf281) |
| EHI_146180 | 2.3813 | Rho family GTPase | Ras-Related Protein Rac1-Related (Pthr24072:Sf281) |
| EHI_104560 | 2.8300 | Cortexillin putative | Cortexillin-2 (Pthr23167:Sf67) |
| EHI_176140 | 2.7438/ 2.4697 | Profilin | Profilin (Pthr11604:Sf0) |
| EHI_174180 | 2.1882 | Skp1 family protein | S-Phase Kinase-Associated Protein 1 (Pthr11165:Sf24) |
| EHI_165350 | 2.6236 | Malate dehydrogenase putative | Hydroxycarboxylate Dehydrogenase B-Related (Pthr11091:Sf0) |
| EHI_159160 | 2.4921 | Superoxide dismutase | Superoxide Dismutase [Fe] 2, Chloroplastic (Pthr42769:Sf3) |
| EHI_011270 | 2.2522 | Uncharacterized protein | Si:Ch211-282j17.12-Related (Pthr18884:Sf83) |
| EHI_178960 | 2.7608/ 2.5979 | Acetyl-CoA synthetase, putative | Acetate--Coa Ligase [Adp-Forming] (Pthr43334:Sf1) |
| EHI_152680 | 2.1693 | EH-domain containing protein putative | At21416p (Pthr11216:Sf31) |
| EHI_147570 | 2.1748/ 2.7106 | Rho GDP exchange inhibitor, putative | Ld16419p (Pthr10980:Sf3) |
| EHI_193330 | 2.2123 | 60S ribosomal protein L23 putative | 60s Ribosomal Protein L23 (Pthr11761:Sf8) |
| EHI_110180 | 2.4310 | Myosin heavy chain | Myosin Heavy Chain, Non-Muscle (Pthr45615:Sf40) |
| EHI_051060 | 3.7812 | Pyruvate ferredoxin oxidoreductase | Pyruvate-Flavodoxin Oxidoreductase-Related (Pthr32154:Sf0) |
| EHI_125950 | 3.1278 | Alcohol dehydrogenase putative | Alcohol Dehydrogenase Yqhd (Pthr43633:Sf1) |
| EHI_006810 | 2.7083 | 14-3-3 protein 3 | 14-3-3 Protein Zeta (Pthr18860:Sf106) |
| EHI_152650 | 2.5095 | Type A flavoprotein, putative | Diflavin Flavoprotein A 2-Related (Pthr32145:Sf11) |
| EHI_044970 | 3.9670 | Malic enzyme | Nadp-Dependent Malic Enzyme (Pthr43237:Sf4) |
| EHI_091250 | 2.8148 | Arp2/3 complex 34 kDa subunit | Actin-Related Protein 2/3 Complex Subunit 2 (Pthr12058:Sf0) |
| EHI_005020 | 2.5429 | F-actin-capping protein subunit beta | F-Actin-Capping Protein Subunit Beta (Pthr10619:Sf0) |
| EHI_009530 | 2.136 | Pyruvate, phosphate dikinase | Pyruvate, Phosphate Dikinase 1, Chloroplastic (Pthr22931:Sf9) |
| Color | Evidence |
|---|---|
| Red | Presence of fusion |
| Green | Neighborhood |
| Blue | Cooccurrence |
| Purple | Experimental |
| Yellow | Text mining |
| Light blue | Database |
| Black | Coexpression |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azmi, N.; Othman, N. Entamoeba histolytica: Proteomics Bioinformatics Reveal Predictive Functions and Protein–Protein Interactions of Differentially Abundant Membrane and Cytosolic Proteins. Membranes 2021, 11, 376. https://doi.org/10.3390/membranes11060376
Azmi N, Othman N. Entamoeba histolytica: Proteomics Bioinformatics Reveal Predictive Functions and Protein–Protein Interactions of Differentially Abundant Membrane and Cytosolic Proteins. Membranes. 2021; 11(6):376. https://doi.org/10.3390/membranes11060376
Chicago/Turabian StyleAzmi, Norhidayah, and Nurulhasanah Othman. 2021. "Entamoeba histolytica: Proteomics Bioinformatics Reveal Predictive Functions and Protein–Protein Interactions of Differentially Abundant Membrane and Cytosolic Proteins" Membranes 11, no. 6: 376. https://doi.org/10.3390/membranes11060376
APA StyleAzmi, N., & Othman, N. (2021). Entamoeba histolytica: Proteomics Bioinformatics Reveal Predictive Functions and Protein–Protein Interactions of Differentially Abundant Membrane and Cytosolic Proteins. Membranes, 11(6), 376. https://doi.org/10.3390/membranes11060376