
Ultrafiltration of Black Soldier Fly (Hermetia illucens) and Mealworm (Tenebrio molitor) Protein Concentrates to Enhance Emulsifying and Foaming Properties

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Defatting of Mealworm (TM) Powder
2.3. Black Soldier Fly (BSF) and Mealworm (TM) Protein Concentrates
2.4. Membranes
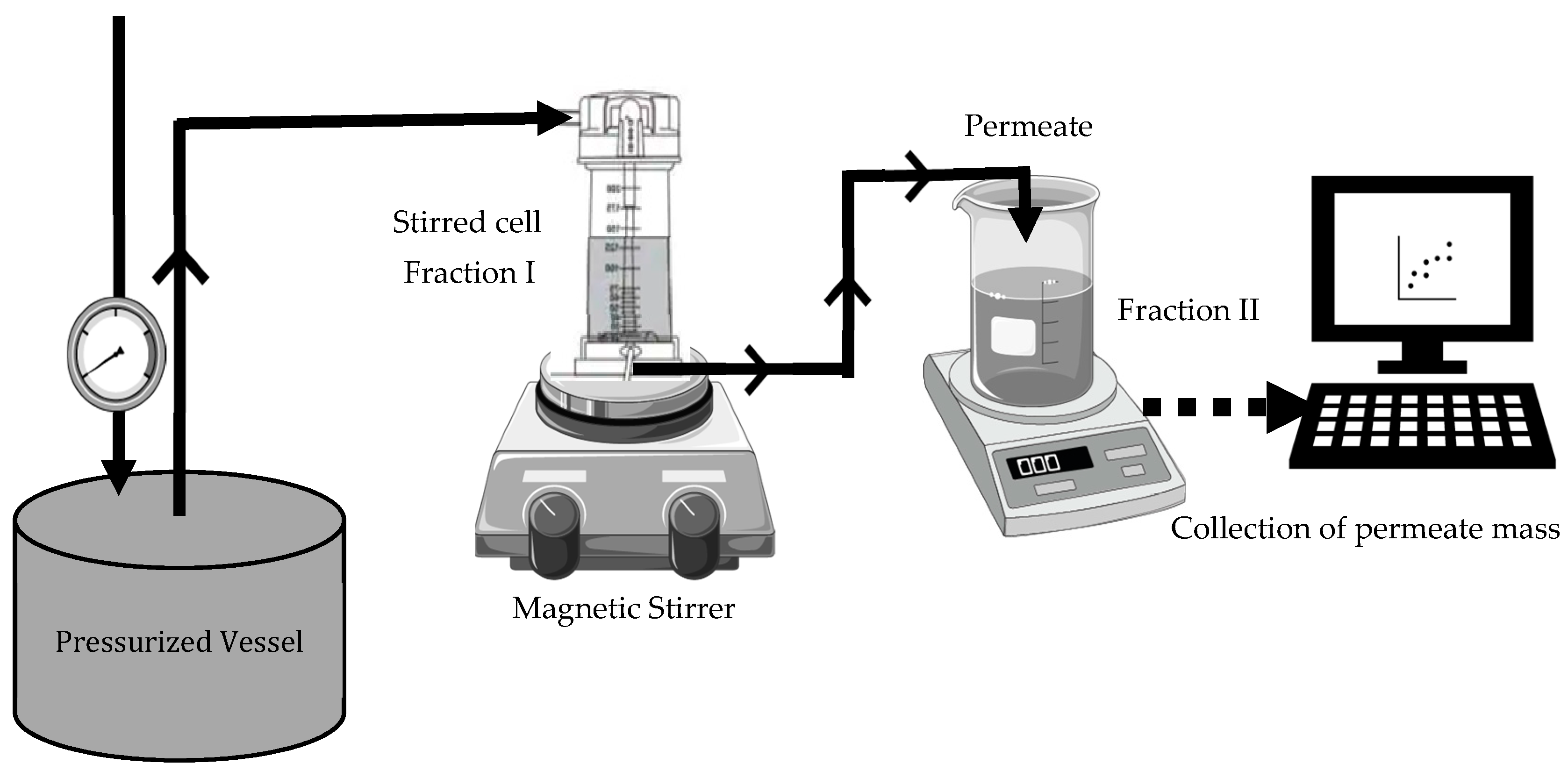
2.5. Ultrafiltration of Black Soldier Fly (BSF) and Mealworm (TM) Protein Concentrates
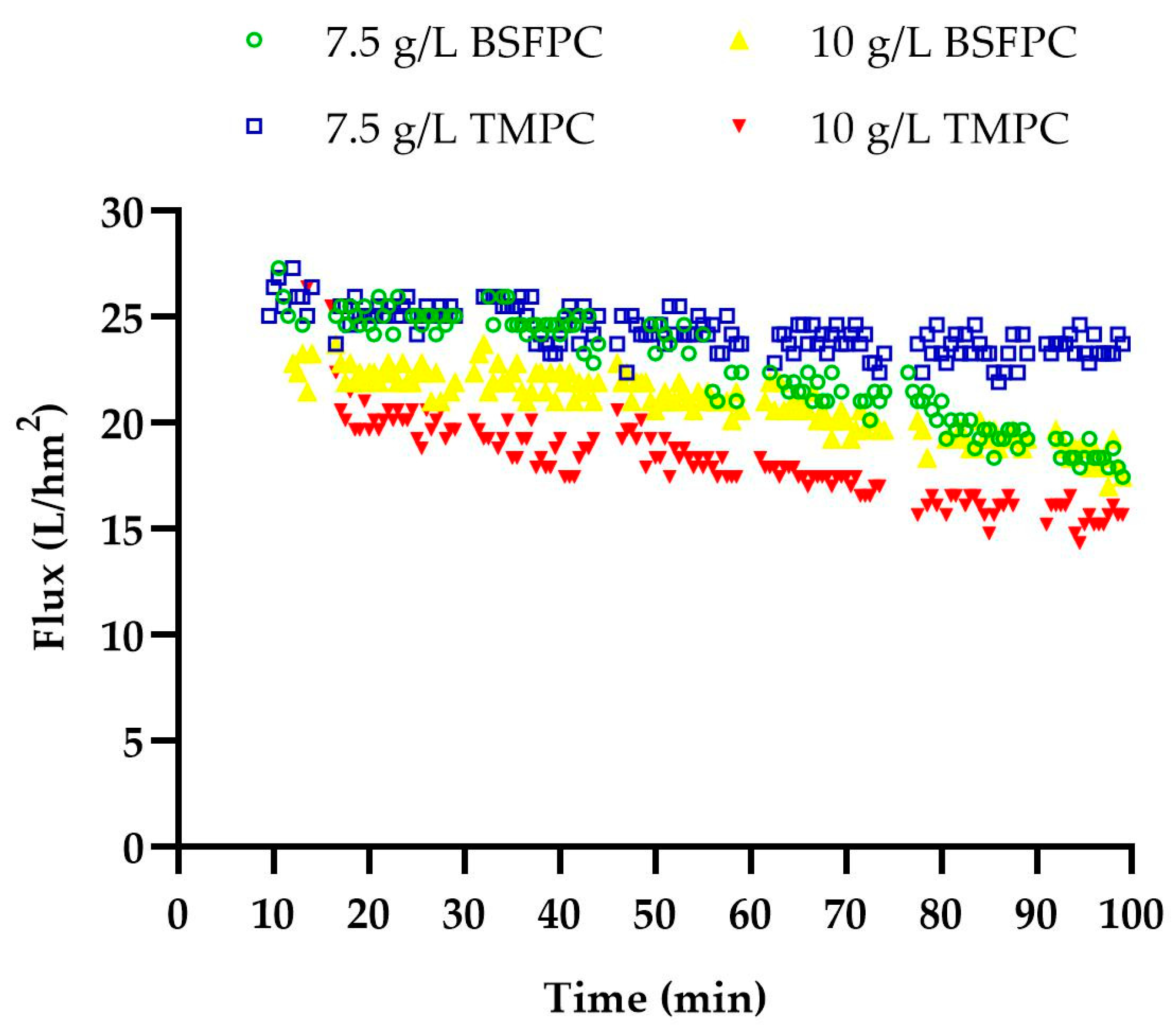
Permeate Flux
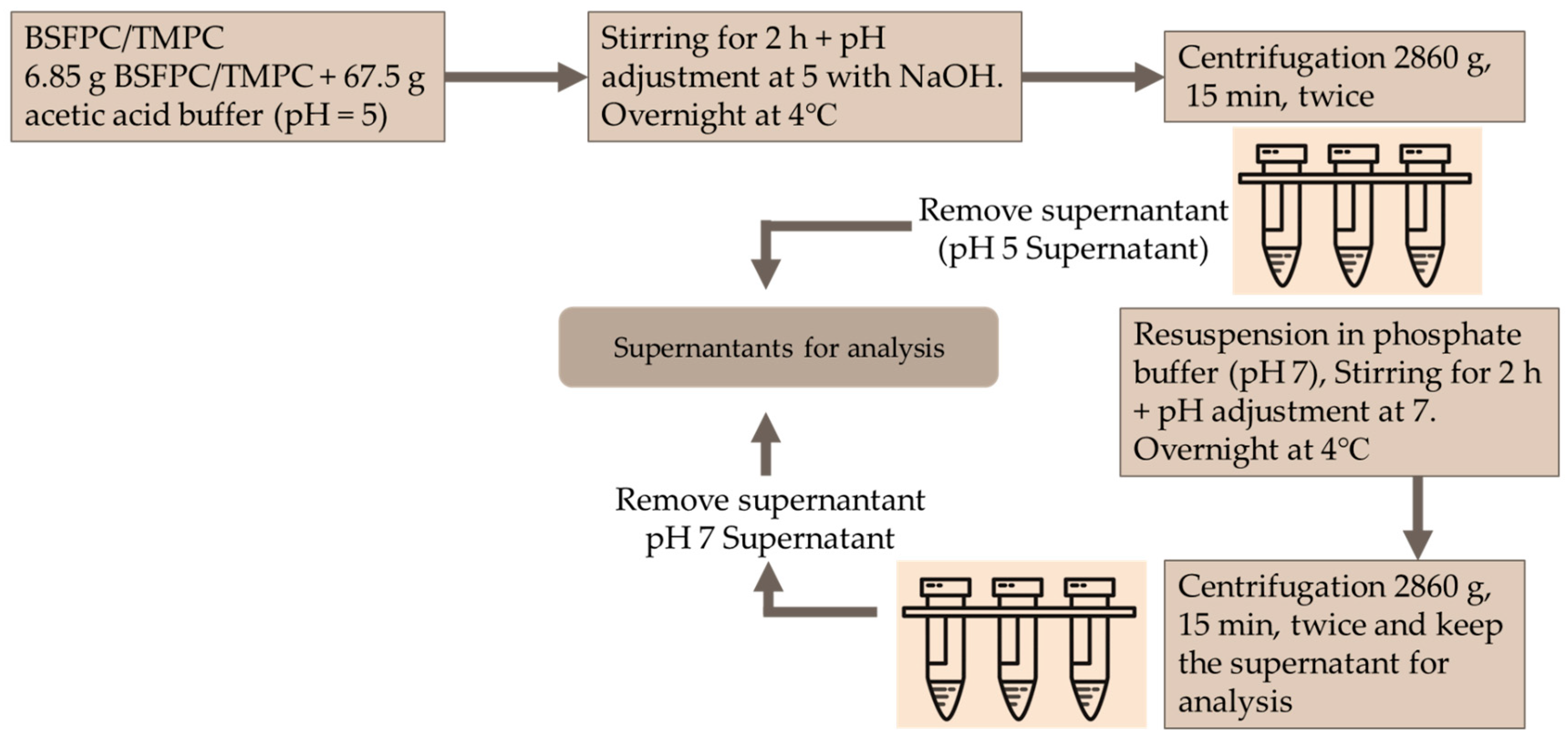
2.6. Black Soldier Fly Protein Concentrate and Mealworm Protein Concentrate Isolation by Acidic Solubilization
2.7. Characterization of the Black Soldier Fly and Mealworm Protein Fractions
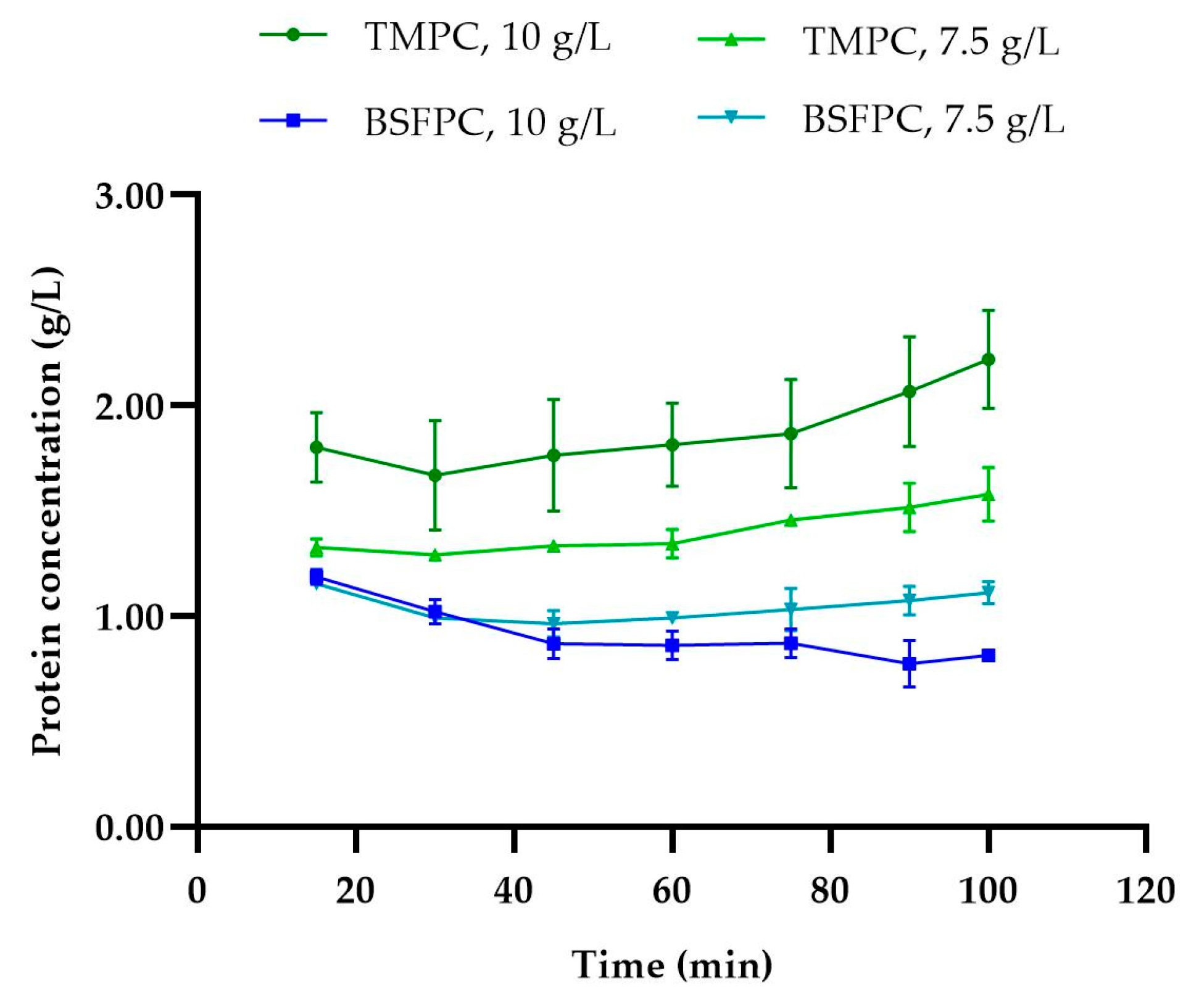
2.7.1. Protein Content
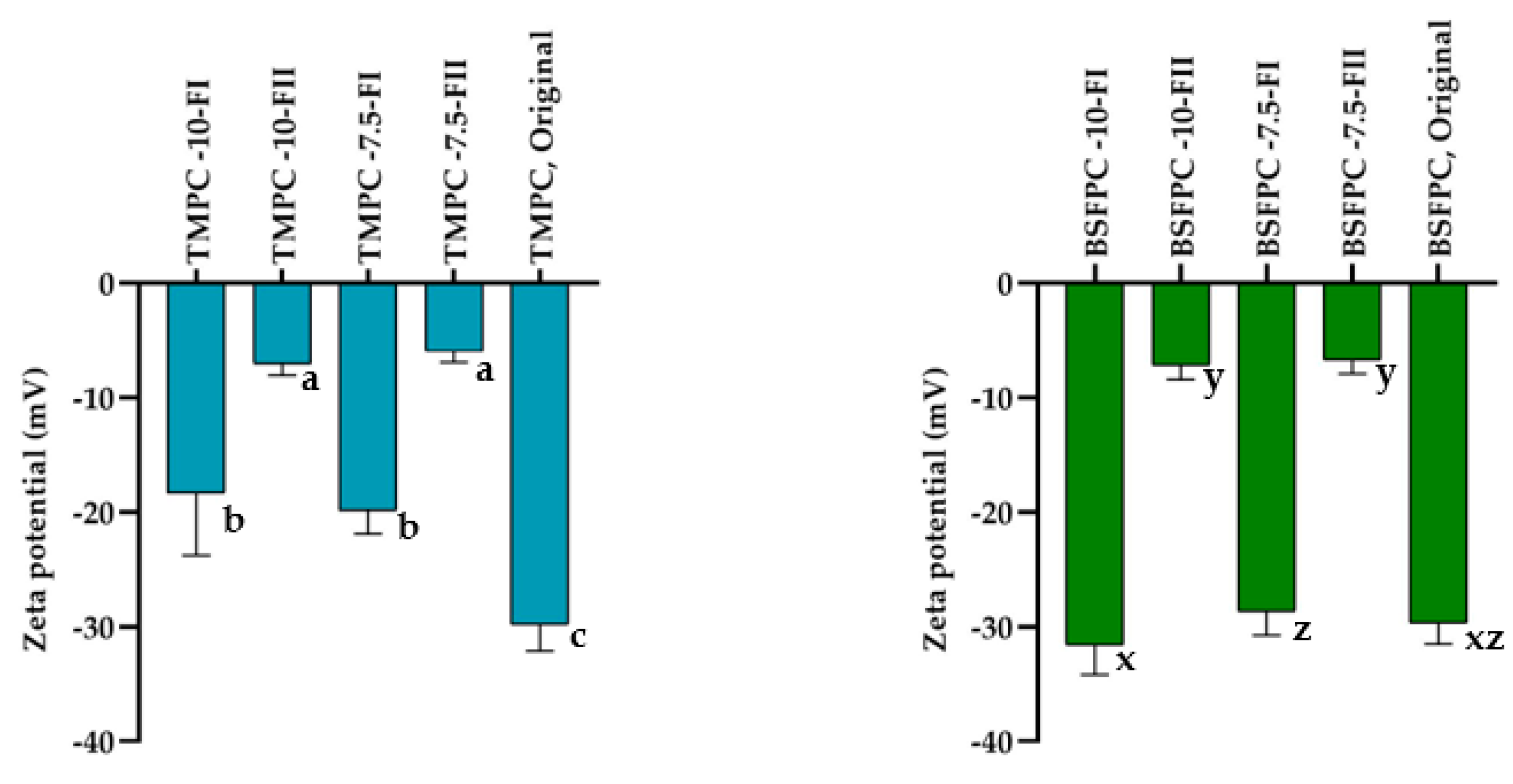
2.7.2. Surface Charge (ζ Potential)
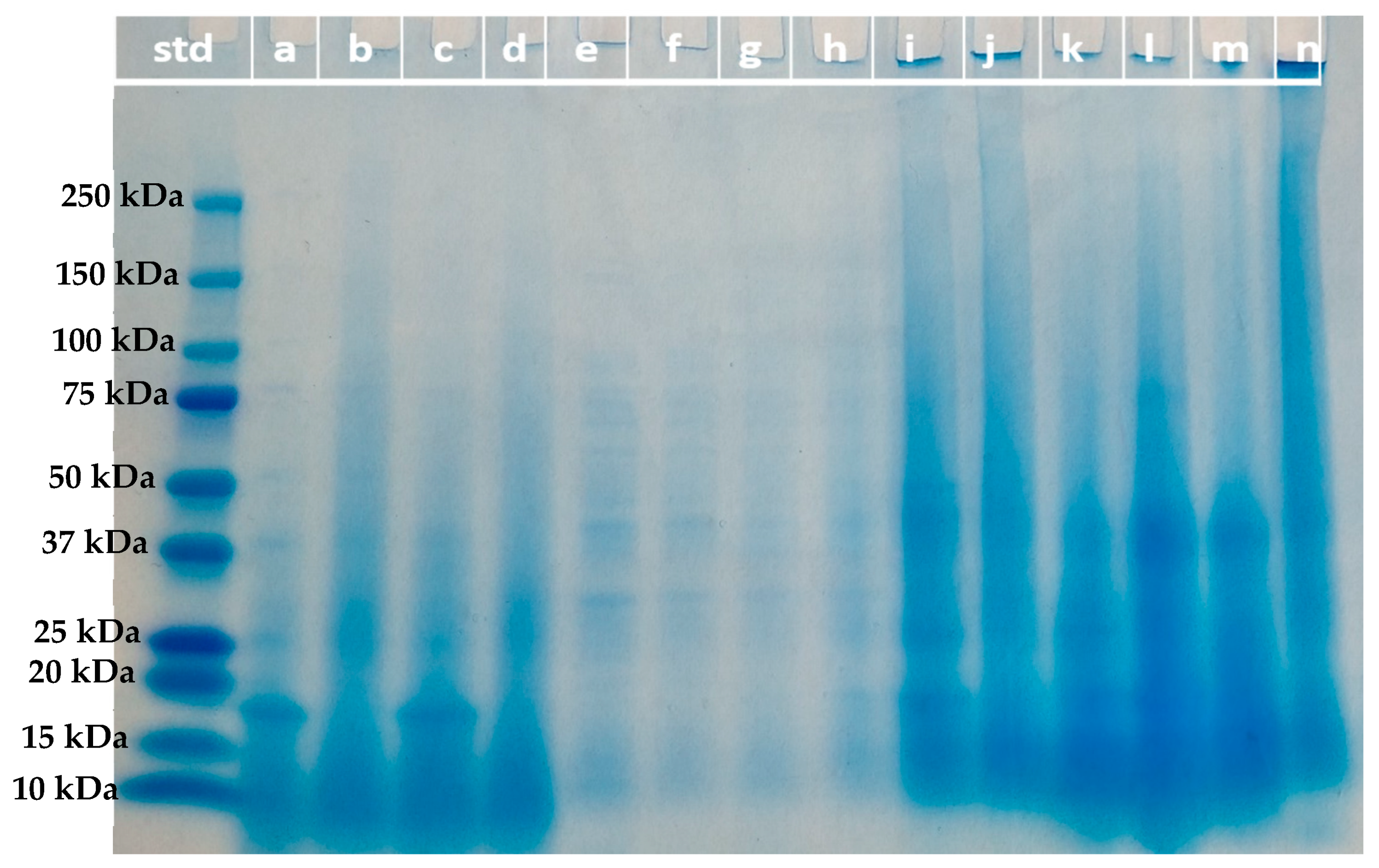
2.7.3. Molecular Weight Distribution of Protein Fractions (SDS-PAGE)
2.8. Techno-Functional Properties of the Black Soldier Fly and Mealworm Protein Fractions
2.8.1. Foaming Capacity and Foam Stability
2.8.2. Emulsifying Activity (EA)
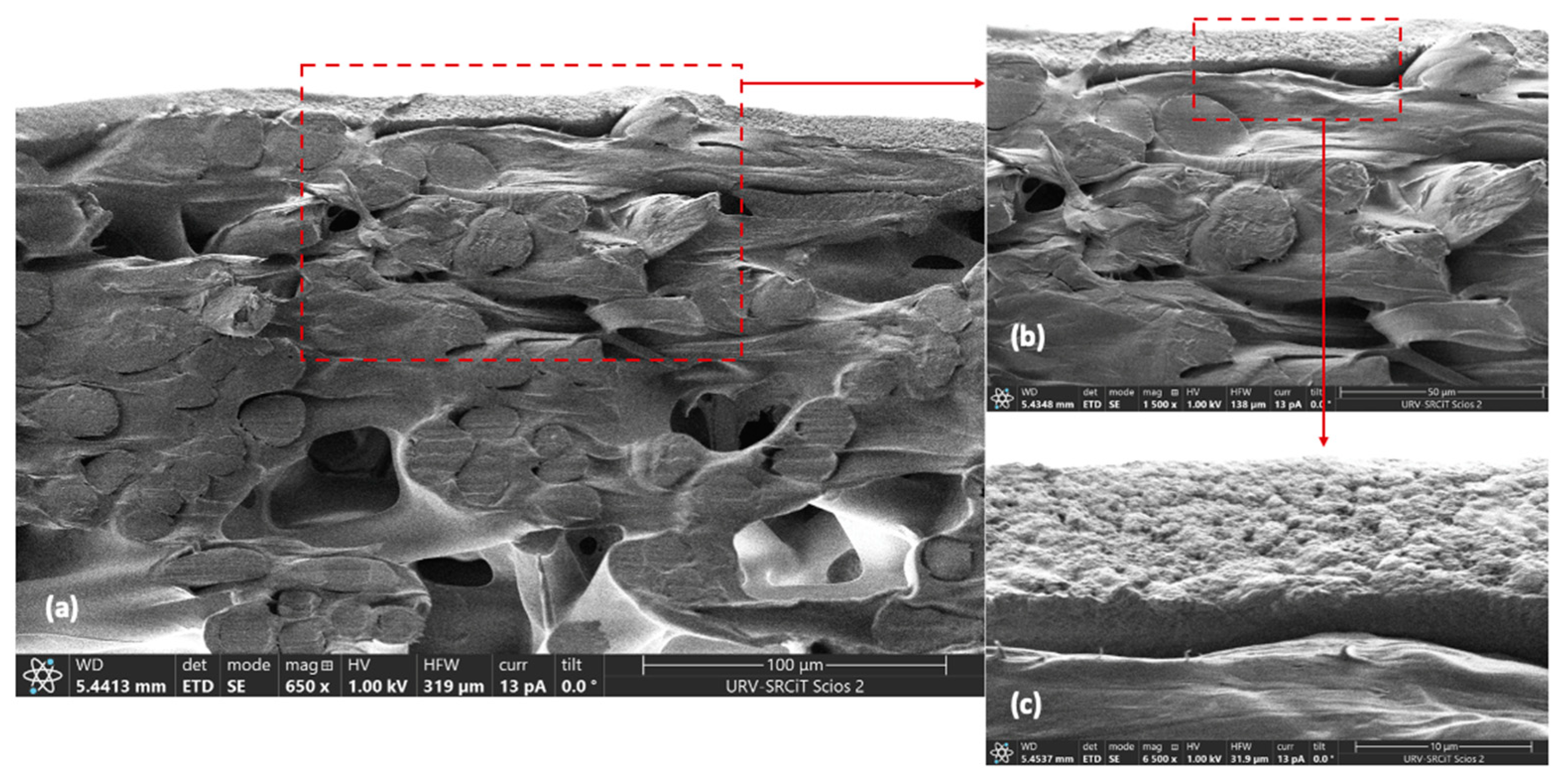
2.9. UF Membrane Characterization
2.10. Statistical Analysis
3. Results and Discussion
3.1. Fractionation of BSFPC and TMPC by Ultrafiltration: Effect of the initial Protein Concentration on Permeate Fluxes, Protein Transmission, and Membrane Performance Recovery
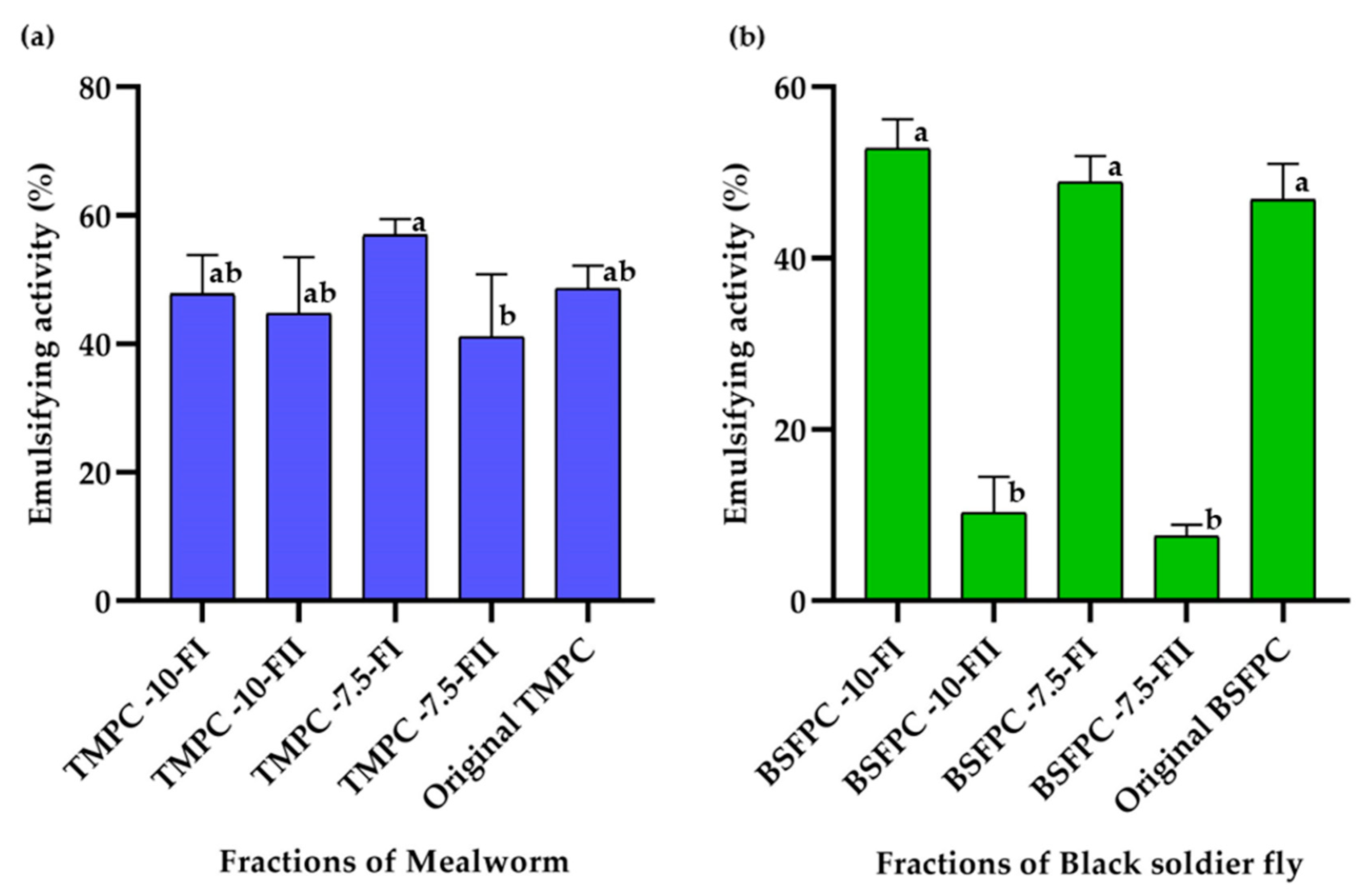
3.2. Emulsifying Activity of the BSFPC and TMPC Fractions
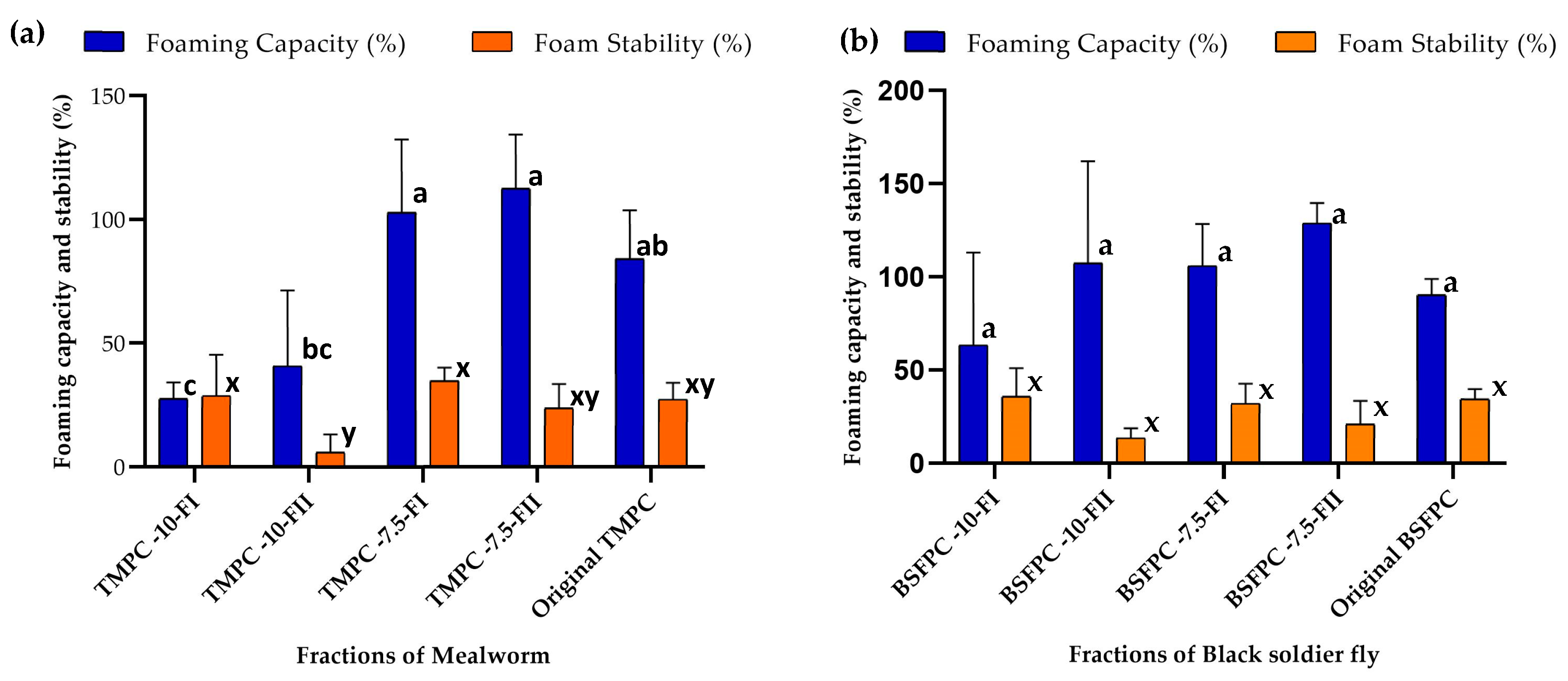
3.3. Foaming Properties of the BSFPC and TMPC Fractions
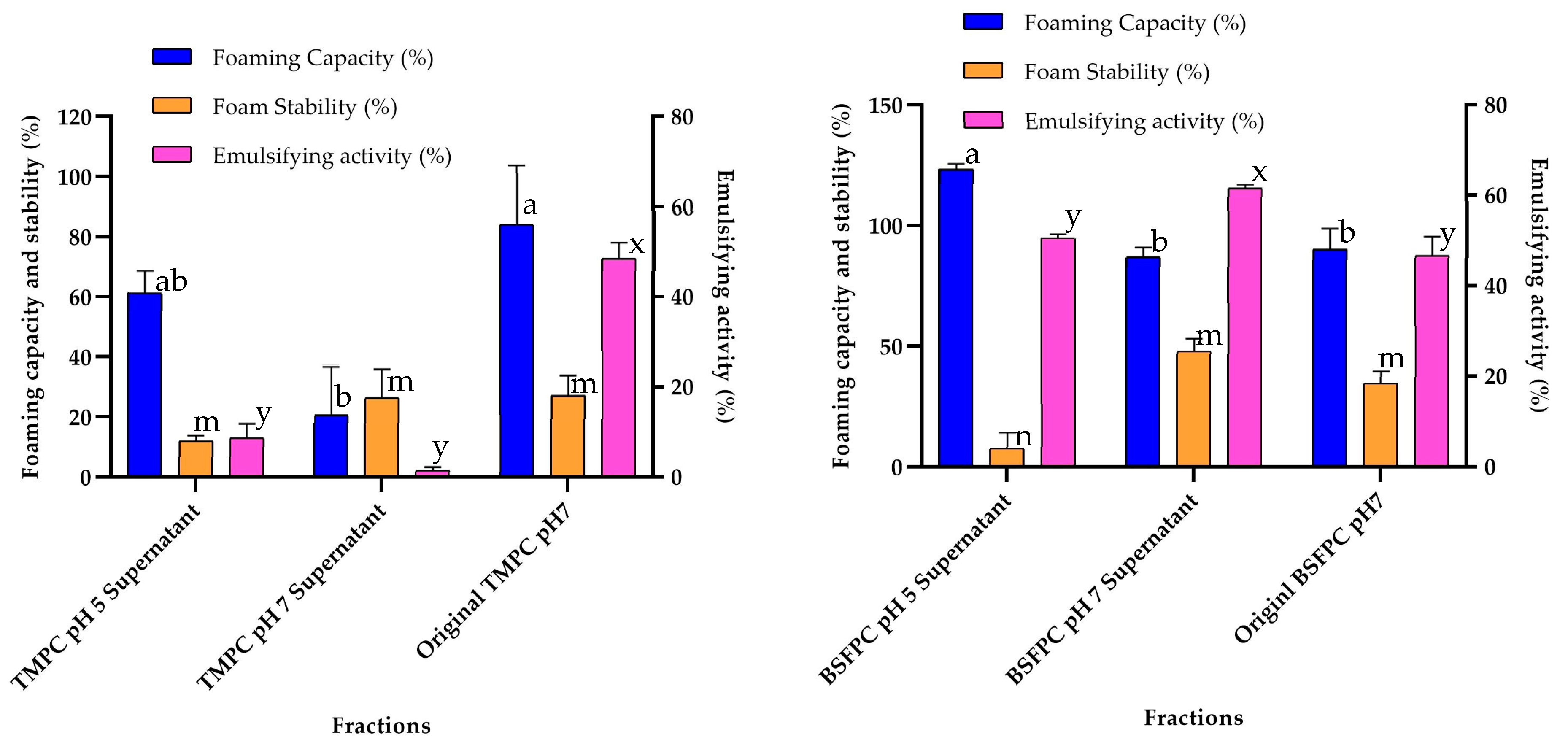
3.4. Effect of Acidic Solubilization Fractionation on Emulsifying and Foaming Properties
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Purschke, B.; Tanzmeister, H.; Meinlschmidt, P.; Baumgartner, S.; Lauter, K.; Jäger, H. Recovery of soluble proteins from migratory locust (Locusta migratoria) and characterization of their compositional and techno-functional properties. Food Res. Int. 2018, 106, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Elorduy, J. Energy supplied by edible insects from Mexico and their nutritional and ecological importance. Ecol. Food Nutr. 2008, 47, 280–297. [Google Scholar] [CrossRef]
- EFSA NDA Panel (EFSA Panel on Nutrition, Novel Foods and Food Allergens); Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Scientific Opinion on the safety of dried yellow mealworm (Tenebrio molitor larva) as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06343. [Google Scholar] [CrossRef]
- Wang, J.; Jousse, M.; Jayakumar, J.; Fernández-Arteaga, A.; de Lamo-Castellví, S.; Ferrando, M.; Güell, C. Black soldier fly (Hermetia illucens) protein concentrates as a sustainable source of stabilize O/W emulsions produced by a low-energy high-throughput emulsification technology. Foods 2021, 10, 1048. [Google Scholar] [CrossRef]
- Gkinali, A.A.; Matsakidou, A.; Vasileiou, E.; Paraskevopoulou, A. Potentiality of Tenebrio molitor larvae-based ingredients for the food industry: A review. Trends Food Sci. Tech. 2022, 119, 495–507. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Unlocking the biological potential of proteins from edible insects through enzymatic hydrolysis: A review. Innov. Food Sci. Emerg. 2017, 43, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Sousa, P.; Borges, S.; Pintado, M. Enzymatic hydrolysis of insect: Alphitobius diaperinus towards the development of bioactive peptide hydrolysates. Food Funct. 2020, 11, 3539–3548. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvements of techno-fucntional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1033. [Google Scholar] [CrossRef]
- Jantzen da Silva Lucas, A.; Menegon de Oliveira, L.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef] [PubMed]
- de Matos, F.M.; Novelli, P.K.; de Castro, R.J.S. Enzymatic hydrolysis of black cricket (Gryllus assimilis) proteins positively affects their antioxidant properties. J. Food Sci. 2021, 86, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Pino, F.; Guadix, A.; Guadix, E.M. Identification of novel dipeltidyl peptidase IV and a-glucosidase inhibitory peptides from Tenebrio molitor. Food Funct. 2021, 12, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, E.; Karas, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Mishyna, M.; Martínez, J.J.I.; Chen, J.; Benjamin, O. Extractions, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhu, Y.; Zheng, Y.; Liu, Z.; Zhong, Y.; Deng, Y.; Zhao, Y. Effects of salting-in/out assisted extractions of structural, physicochemical and functional properties of Tenebrio molitor larvae protein isolates. Food Chem. 2021, 338, 128158. [Google Scholar] [CrossRef]
- Azagoh, C.; Ducept, F.; García, R.; Rakotozafy, L.; Cuvelier, M.E.; Keller, S.; Lewandowski, R.; Mezdour, S. Extraction and physicochemical characterization of Tenebrio molitor proteins. Food Res. Int. 2016, 88, 24–31. [Google Scholar] [CrossRef]
- Santiago, L.A.; Fadel, O.M.; Tavares, G.M. How does the thermal-aggregation behavior of black cricket protein isolate affect its foaming and gelling properties? Food Hydrocolloid. 2021, 110, 106169. [Google Scholar] [CrossRef]
- Mshayisa, V.V.; Van Wyk, J.; Zozo, B. Nutritional, techno-functional and structural properties of black solider fly (Hermetia illucens) larvae flours and protein concentrates. Foods 2022, 11, 724. [Google Scholar] [CrossRef]
- Wang, J.; Ballon, A.; Schroën, K.; de Lamo-Castellví, S.; Ferrando, M.; Güell, C. Polyphenol loaded W1/O/W2 emulsions stabilized with lesser mealworm (Alphitobius diaperinus) protein concentrate produced by membrane emulsification: Stability under simulated storage, process, and digestion conditions. Foods 2021, 10, 2997. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Vanqa, N.; Mshayisa, V.V.; Basitere, M. Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours. Foods 2022, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.; Dey, T.K.; Ghosh, M.; Dhar, P. Enzymatic modification of sesame seed protein, sourced from waste resource for nutraceutical application. Food Bioprod. Process. 2015, 94, 70–81. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Permeate Flux (L/h m2) |
|---|---|
| 10 g/L BSFPC | 18.32 ± 0.73 |
| 7.5 g/L BSFPC | 18.42 ± 0.53 |
| 10 g/L TMPC | 15.58 ± 0.59 |
| 7.5 g/L TMPC | 23.64 ± 0.48 |
| Fractionation Method | Insect Protein | Fraction | Label | Protein Content (g/L) |
|---|---|---|---|---|
| Ultrafiltration | 7.5 g/L BSFPC | Fraction I | BSFPC-7.5-FI | 15.09 ± 0.24 |
| 7.5 g/L TMPC | Fraction I | TMPC-7.5-FI | 15.15 ± 0.29 | |
| 7.5 g/L BSFPC | Fraction II | BSFPC-7.5-FII | 1.14 ± 0.01 | |
| 7.5 g/L TMPC | Fraction II | TMPC-7.5-FI | 1.41 ± 0.01 | |
| 10 g/L BSFPC | Fraction I | BSFPC-10-FI | 15.88 ± 2.22 | |
| 10 g/L TMPC | Fraction I | TMPC-10-FI | 26.76 ± 1.80 | |
| 10 g/L BSFPC | Fraction II | BSFPC-10-FII | 0.91 ± 0.03 | |
| 10 g/L TMPC | Fraction II | TMPC-10-FII | 1.80 ± 0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranasinghe, M.K.; Ballon, A.; de Lamo-Castellví, S.; Ferrando, M.; Güell, C. Ultrafiltration of Black Soldier Fly (Hermetia illucens) and Mealworm (Tenebrio molitor) Protein Concentrates to Enhance Emulsifying and Foaming Properties. Membranes 2023, 13, 137. https://doi.org/10.3390/membranes13020137
Ranasinghe MK, Ballon A, de Lamo-Castellví S, Ferrando M, Güell C. Ultrafiltration of Black Soldier Fly (Hermetia illucens) and Mealworm (Tenebrio molitor) Protein Concentrates to Enhance Emulsifying and Foaming Properties. Membranes. 2023; 13(2):137. https://doi.org/10.3390/membranes13020137
Chicago/Turabian StyleRanasinghe, Madushika K., Aurélie Ballon, Sílvia de Lamo-Castellví, Montserrat Ferrando, and Carme Güell. 2023. "Ultrafiltration of Black Soldier Fly (Hermetia illucens) and Mealworm (Tenebrio molitor) Protein Concentrates to Enhance Emulsifying and Foaming Properties" Membranes 13, no. 2: 137. https://doi.org/10.3390/membranes13020137
APA StyleRanasinghe, M. K., Ballon, A., de Lamo-Castellví, S., Ferrando, M., & Güell, C. (2023). Ultrafiltration of Black Soldier Fly (Hermetia illucens) and Mealworm (Tenebrio molitor) Protein Concentrates to Enhance Emulsifying and Foaming Properties. Membranes, 13(2), 137. https://doi.org/10.3390/membranes13020137