3. Results and Discussion
The miscibility of phospholipids in a hydrated bilayer is an issue of fundamental importance to membrane biophysics [
28]. This research aimed to investigate the impact of the unsaturation of lipid acyl chains on the molecular organization of mixed lipid membranes composed of phosphatidylcholines (PCs), and to understand the driving forces causing the immiscibility of PCs. It has previously been shown experimentally that DOPC and DPPC exhibit limited miscibility in a mixed bilayer [
5]. Therefore, we used this lipid pair to further explore the reason for differences in immiscibility between PC molecules. Both lipids share the same headgroup but differ in the length and saturation of the acyl chains. This enabled us to isolate the effect of the hydrophobic part of the lipids on their behavior in the mixed bilayers.
DSC Measurements. DSC measurements were used to determine the phase diagram for the mixed DPPC/DOPC bilayers. DSC thermograms for systems with various
XDOPC are depicted in
Figure S1. The thermogram for the pure DPPC membrane displays two endothermic peaks at
Tp = 35 °C and
Tm = 41.2 °C. The first peak corresponds to the pre-transition from the gel phase (
Lβ′, also called the solid-ordered phase) to the ripple phase (
Pβ′). In the
Lβ′ state, the lipids are arranged on a two-dimensional triangular lattice in the plane of the membrane [
1]. DPPC chains are predominantly ordered into the all-
trans configurations and tilted with respect to the membrane normal. For fully hydrated DPPC bilayers at 19–25 °C, the tilt angle is about 30° [
29]. The
Pβ′ phase is characterized by periodic one-dimensional ripples on the membrane surface. The ripples are probably formed by periodic arrangements of the linear gel (
Lβ′) and fluid (
Lα) lipid domains [
30,
31]. The second peak corresponds to the main transition (membrane melting) from the
Pβ′ phase to the liquid phase (
Lα, also known as the liquid-disordered or liquid-crystalline phase). The main transition peak is narrow, which indicates that the melting is strongly cooperative. DPPC molecules melt simultaneously in large clusters of
n lipids (these
n lipids are considered as the size of the cooperative unit) [
1]. The larger the size of the cooperative unit, the narrower the transition peak. In the
Lα phase, the acyl chains are mostly disordered with many
trans- and
gauche configurations along the chain, and the lattice order is lost [
1]. In turn, the DOPC membrane has a main transition temperature of −16.5 °C [
32]. This means that the DOPC bilayer is in the liquid phase over the whole temperature range studied here.
The admixture of DOPC into the DPPC bilayer resulted in alterations in the gel-to-liquid state transition. The pre-transition peak disappeared, while the main transition peak became broader, and the Tm value decreased with increasing DOPC content in the membrane. This shows that the various lipid species in mixtures do not melt independently but rather influence each other in the melting process, and that the presence of DOPC affects the DPPC melting process. The addition of DOPC to the DPPC membrane leads to a broadening of the heat capacity profiles and thus a reduction in the DPPC cooperative unit size.
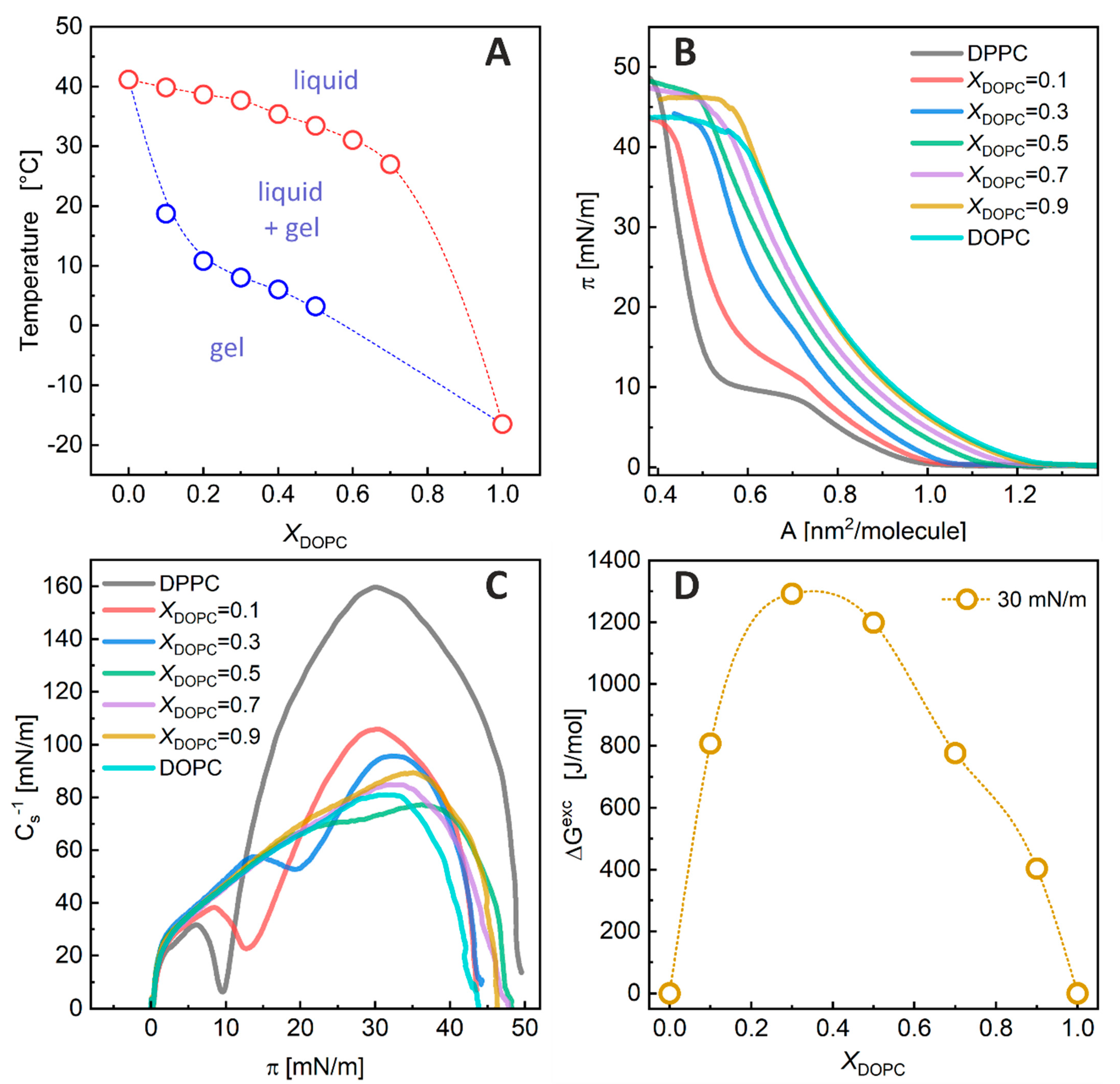
A pseudo-binary phase diagram was constructed from the thermograms by plotting the transition temperatures from a single-phase state to the coexistence of two phases as a function of the sample composition. The transition onset and completion temperatures were determined by analyzing the heat capacity curves using the tangent method [
33].
Figure 2A shows the phase boundaries among the gel phase, liquid-crystalline phase, and their coexistence region for the DPPC/DOPC membrane. The phase diagram indicates that at 50 °C, the DPPC/DOPC membranes of any composition were in the liquid phase. In contrast, at 22 °C, the systems containing
XDOPC ≤ 0.1 were in the gel phase and those with
XDOPC ≥ 0.8 were in the liquid-crystalline state, while the coexistence of both phases was observed within these limits.
The DPPC/DOPC phase diagram. Our phase diagram determined for multilamellar vesicles shows that at 22 °C, the DPPC/DOPC membranes with
XDOPC ≤ 0.1 are in the gel phase, and those containing
XDOPC ≥ 0.8 are in the liquid phase, while the coexistence of liquid–gel phases is observed within these limits. As expected, for temperatures higher than 41 °C, the mixed DPPC/DOPC bilayers are solely in the liquid state. Due to the nature of DSC measurements, we were only able to determine a part of the phase diagram (for temperatures higher than 5 °C). A more detailed phase diagram for DOPC/DPPC-
d62 mixtures was previously determined using
2H NMR experiments [
6]. According to this phase diagram, two-phase systems at 22 °C for large multilamellar DOPC/DPPC-
d62 liposomes at pH 7.0 were observed in the range of 0.3 <
XDOPC ≤ 0.65. Using X-ray scattering, Furuya and Mitsui determined the phase diagram for DPPC/DOPC multilamellar vesicles [
5], which indicated that the liquid–gel phase coexistence occurs within 0.1 <
XDOPC ≤ 0.7. In turn, the fluorescence measurement revealed that this range could change depending on the method of liposome preparation [
34]. Therefore, the observed difference in phase diagrams for mixed lipid membranes may be due to the difference in the method of liposome preparation, as well as due to the experimental method used in the measurements. The DSC experiments used in this study clearly showed that at
XDOPC = 0.7, there are still DPPC domains that melt with increasing temperature, so the DPPC/DOPC membranes with this composition undergo phase separation at lower temperatures.
Langmuir Monolayer Experiments. The measured π–A isotherms and the values of the compression modulus (C
S−1) calculated on their basis are shown in
Figure 2B,C, respectively. The isotherms for the pure DPPC and DOPC monolayers are consistent with those previously published [
12,
35]. The shape of the π–A isotherm for the DOPC film and its C
S−1 − π dependency (
Figure 2C) indicate that this monolayer is in the liquid (L) phase. For DPPC, an inflection region in the isotherm and a minimum in the C
S−1 vs. π plot indicate that this monolayer changes its physical state during compression. The phase transition corresponds to a change from the liquid-expanded (Le) to the liquid-condensed (Lc) state. The addition of DOPC to the DPPC monolayer shifts the Le-Lc phase transition towards higher surface pressures (π
Le-Lc = 9.6, 12.8, 19.6, and ~30 mN/m for
XDOPC = 0.0, 0.1, 0.3, and 0.5, respectively), indicating that the mixed DOPC/DPPC films are less condensed than the pure DPPC monolayer. For
XDOPC ≥ 0.7, the inflections resulting from the Le-Lc phase transition vanished in the isotherms—indicating that at 22 °C, the monolayers of these compositions are in the L phase, similar to the pure DOPC film.
It has been shown that the properties of lipid monolayers at surface pressures in the range of 30–35 mN/m correspond to the properties of bilayers [
36]. Therefore, to shed more light on the miscibility of monolayer components, the A values at a surface pressure of 30 mN/m were determined from the isotherms, and the values of ΔG
Exc [
37] were calculated and plotted versus the DOPC mole fraction (
Figure 2D) to compare the magnitude of interactions between components in the DPPC/DOPC monolayers. The excess mixing free energy values were positive for all the DPPC/DOPC monolayers, showing that the intermolecular interactions in the mixed films were unfavorable compared to those in the one-component films. The strongest repulsion between lipid molecules was observed for an
XDOPC in the range of 0.3–0.5. The maximum of ΔG
Exc was about 1.3 kJ/mol.
The excess free energy of mixing. Our experiments showed a significant influence of the saturation of the lipid chains on the mutual miscibility between PCs. A comparison of one-component DPPC and DOPC monolayers showed that DPPC molecules are more densely packed in the monolayer, and thus more ordered than DOPC molecules in their films, due to the bending of the hydrocarbon chains in the latter lipid. Miscibility between DPPC and DOPC lipids is thermodynamically unfavorable in all mixed membrane compositions, as the values of the excess free energy of mixing are positive and reach a maximum of 1.3 kJ/mol at an XDOPC of about 0.3–0.5.
We previously investigated miscibility in mixed phosphatidylcholine (PC)–phosphatidic acid (PA) bilayers in which the lipids differed in their degree of unsaturation [
12,
35]. In the case of the DPPC–DPPA system, the maximal value of the excess free energy of mixing was about −0.5 kJ/mol, indicating that the interactions between the components are more attractive than for the pure DPPC and DPPA monolayers. BAM images for selected DPPC–DPPA film compositions, showing monolayer morphology, demonstrated that the film components are miscible over the entire range of monolayer compositions. For DOPC–DOPA mixtures, the ΔG
Exc values were only slightly positive (<0.08 kJ/mol), and the monolayers were completely homogeneous for all film compositions. This indicates an ideal mixing between DOPC and DOPA. In contrast, the mixture of DOPC with DPPA showed large positive deviations from the ideal, and the maximal ΔG
Exc value was ca. 1.7 kJ/mol, indicating that the mixing of DOPC and DPPA molecules is thermodynamically unfavorable. Phase separation resulting from the immiscibility of the monolayer components was clearly visible in the BAM images, where, regardless of the monolayer composition, bright condensed DPPA domains dispersed in the fluid DOPC matrix were visible. More complicated behavior was observed for the DPPC/DOPA mixed monolayers [
12]. The positive ΔG
Exc values for films containing higher amounts of DOPA (about 0.4 kJ/mol) indicated that lipid miscibility was not thermodynamically favorable. However, the BAM images showed no signs of phase separation. This means that the repulsive interactions between the mixed pairs were rather weak to cause the immiscibility of the DPPC/DOPA membrane. At a low DOPA content (
XDOPA ≤ 0.3), the mixing of DPPC and DOPA lipids in monolayers was thermodynamically favorable due to the strong attractive electrostatic interactions between their headgroups, resulting in monolayer condensation. The maximal ΔG
Exc value for the DPPC/DOPC system is comparable to that for the DOPC/DPPA system. This indicates that the structure of the acyl chains of individual lipids has a much greater impact on their miscibility compared to possible interactions within the headgroups. In conclusion, phase separation is observed when ΔG
Exc exceeds a certain limit. Based on the aforementioned values, we suppose that this limit is in the range of 0.8–1.2 kJ/mol. For positive ΔG
Exc values which remain below this limit, small lipid aggregates may form, but these are too small to be observed by BAM methods. However, this hypothesis requires further experimental studies.
MD Simulations. We performed MD simulations of mixed DPPC/DOPC bilayers with different lipid compositions at 22 °C (295 K) and 50 °C (323 K).
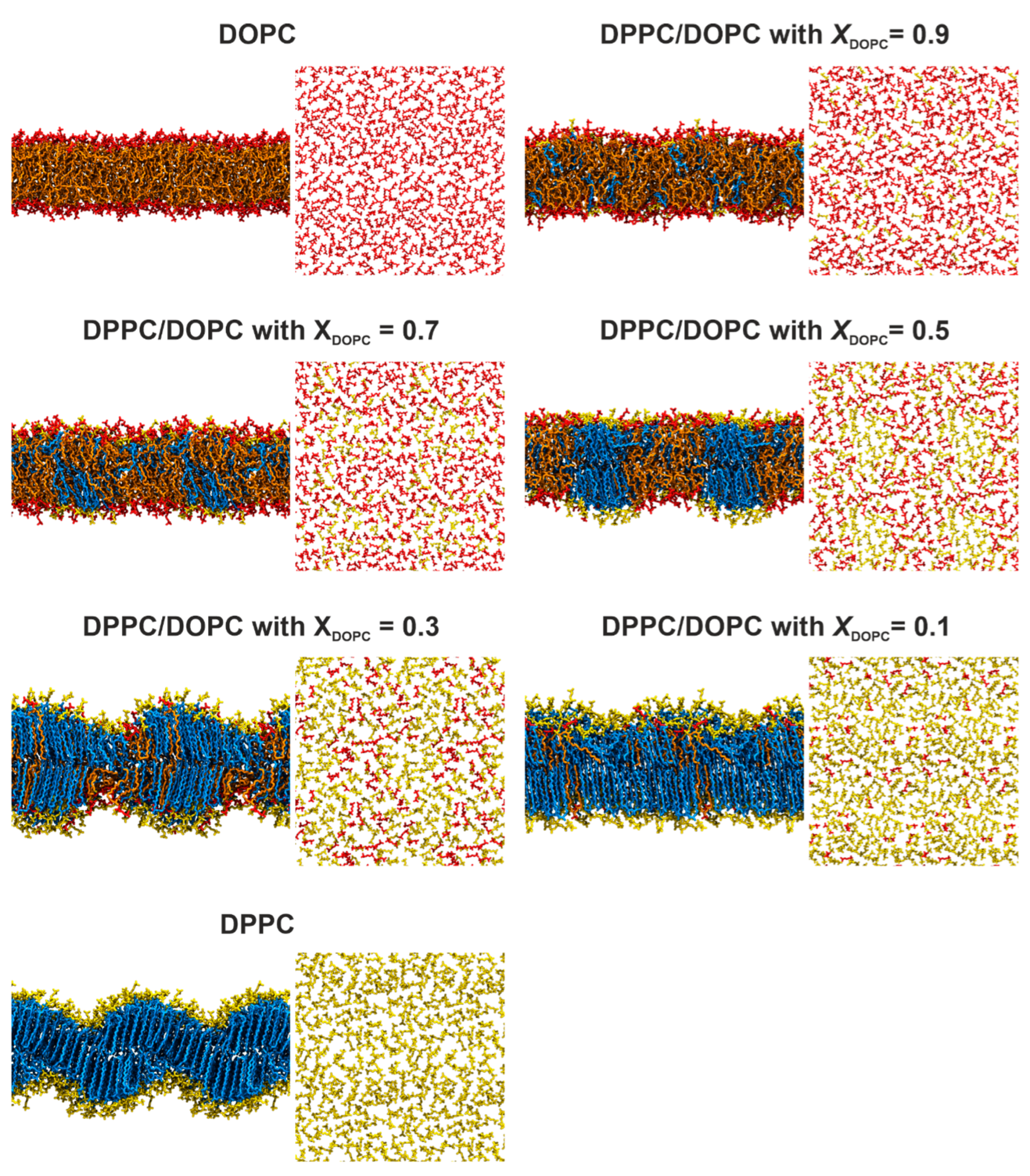
Figure 3 and
Figure S2 present typical snapshots showing the organization of the DPPC/DOPC membranes at the end of the simulation at 295 K and 323 K, respectively. At the higher temperature, all the bilayers exhibit a pronounced disorder of the acyl chains with a large number of
gauche configurations, which is characteristic of membranes in the liquid phase. The lipid molecules are evenly distributed in the mixed bilayer, which indicates a good miscibility of the lipids at temperatures above the phase transition. On the contrary, at the lower temperature (
Figure 3), the DPPC membrane shows a structure that resembles the ripple phase (
Pβ′). The presence of two regions with different molecular ordering patterns can be observed. In one region, the DPPC molecules are stretched and tightly packed in a manner characteristic of the gel phase. The two leaflets are well separated, resulting in increased membrane thickness. In the other region, the acyl chains are disordered, and the leaflets are partially interdigitated, which reduces the thickness of the bilayer.
For the mixed DPPC/DOPC bilayers at 295 K, containing 50 mol% or more DPPC, two regions with very distinctive molecular ordering patterns were observed. The first region contains tightly packed lipid molecules (mainly DPPC) with stretched chains arranged similarly to the pure DPPC bilayer described above. The second region contains loosely packed lipid molecules (mostly DOPC) with partially interdigitated hydrocarbon chains. This molecular arrangement changes for the bilayers with a lower DPPC content (XDOPC = 0.7 and 0.9). In these cases, there is no clear phase separation between DPPC and DOPC lipids, although DPPC molecules seem to adhere to each other to form small aggregates. The lipid bilayers are much more disordered and there is no clear separation between lipid leaflets.
Area per lipid. In order to gain more insight into the lipid organization in the DPPC/DOPC bilayers, the average area per lipid (APL), the average area per DPPC (APP), and the average area per DOPC (APO) were calculated (
Table 2). APL was computed by dividing the area of the simulation box by the number of lipids in one leaflet, while the APP and APO in each membrane were calculated using the Voronoi tessellation method implemented in the APL@VORO software [
38].
The calculated APL value for the pure DPPC membrane is 0.664 nm
2 at 323 K. This value is very close to experimental results (ca. 0.629 [
39], 0.64 [
40], 0.681 [
41], and 0.68 [
42] nm
2) determined at the same temperature. Similarly, the calculated APLs for the pure DOPC bilayer are 0.744 nm
2 and 0.722 nm
2 for 323 K and 295 K, respectively, and are also in good agreement with the experimental values of 0.721 nm
2 [
41] and 0.722 nm
2 [
43] measured at 298 K and 303 K, respectively. As expected, an increase in DOPC content in the mixed membrane leads to a gradual increase in APL. The average values of APP and APO also increase with the increasing DOPC concentration in the bilayer. At lower temperature, APPs are much smaller compared to APOs for
XDOPC ≤ 0.5, but for higher unsaturated lipid content both values become similar. This shows that in the membranes with
XDOPC > 0.5, the palmitoyl chains of DPPC become more disordered, which increases the surface area occupied by this lipid. At a higher temperature, the differences between APL and APP or APO are insignificant.
It should be noted that the average value of area per DPPC (APP) in the pure DPPC bilayer at 323 K is lower than the average value of the area per lipid (APL). The same is true for the APO and APL values for the pure DOPC bilayer. This difference is because the Voronoi tessellation method does not take into account voids between lipids when reporting the area per lipid (APO and APP), while the average APL values include them. At the higher temperature, the differences between APL and APP or APO are greater than those at the lower temperature, confirming that at the higher temperature the lipids are further apart, making the structure much looser.
Membrane thickness. The mass density profiles for selected lipid groups for the systems simulated at 295 K are depicted in
Figure 4. These profiles show the distributions of the various lipid groups along the normal to the bilayer. The distributions of DOPC phosphorus atoms are shifted towards the center of the mixed bilayer compared to the distributions of DPPC phosphorus atoms (
Figure 4). This is particularly evident for
XDOPC < 0.7 and suggests that DOPC lipids are, on average, located deeper in the mixed lipid bilayer compared to DPPC molecules. At 323 K (
Figure S3, Supplementary Materials), this effect is much weaker, showing that at the higher temperature the headgroups of both lipids are in similar positions in the membrane. The distributions of DPPC phosphorus atoms at 295 K are much more asymmetric compared to the distributions of the DOPC phosphorus atoms. This trend disappears at 323 K, where the distributions of DPPC phosphorus atoms become fully symmetric. Interestingly, the water-mass-density profiles (
Figure 4) show that the depth of the water penetration deep into the lipid bilayer increases with decreasing DOPC content in the mixed membranes. This effect is more pronounced at lower temperatures.
The bilayer thickness (
dP) was calculated as an average distance between the phosphorus atoms in the opposite leaflets for each type of lipid separately (
Table S1). The calculated
dP values for pure DPPC and DOPC membranes are in good agreement with the experimental results. The bilayer thickness of DPPC, DOPC, and an equimolar mixture of these two phospholipids at 50 °C was determined experimentally (using X-ray diffraction measurements) to be 4.02, 3.64, and 3.74 nm, respectively [
44]. It follows that the thickness of the bilayer formed by the equimolar mixture is intermediate between the two pure phospholipid bilayers. The thickness of mica-supported DPPC and DOPC bilayers was previously determined using atomic-force microscopy (AFM) [
45]. For DOPC, the AFM measurements revealed a membrane thickness of about 4.1 nm at 22 °C. In the case of DPPC, AFM topography images showed a bilayer thickness of 4.8 ± 0.3 nm at 22 °C (gel phase), and this value decreased to 3.3 ± 0.3 nm when the temperature was increased to 52 °C. The authors noted that the membrane thicknesses obtained from the cross-section of AFM images at room temperature were larger than the values determined by other experimental methods. As an explanation, they stated that AFM can overestimate the thickness of the lipid membrane at lower temperatures due to the reduced mobility of both lipids and water molecules associated with the bilayer interface.
At the lower temperature, the membrane thickness with respect to DOPC is almost constant in all simulated bilayers and is significantly smaller than that for DPPC. For DPPC, the
dP value changed with the composition of the mixed bilayer. It first increases to a value of about 4.3 nm at
XDOPC = 0.3 and then decreases to a constant value of about 3.9 nm. Such a non-linear relationship is probably related to the fact that in our simulations the pure DPPC bilayer has a ripple phase structure (see
Figure 2), where the membrane thickness varies periodically. Therefore, the averaged
dP value is underestimated. The introduction of DOPC leads to the gradual disappearance of the ripple structure, so the resulting average value reaches a value characteristic of the thickness of the DPPC membrane in the solid phase. At 323 K, the thickness of the mixed membrane is constant for both lipids, with the
dP value for DPPC tending to be larger compared to DOPC, indicating that the unsaturated lipid molecules are located slightly deeper in the membrane. Despite the good agreement between the experimental and calculated values of the bilayer thickness, it is worth noting that the choice of the force field and the method of calculating bilayer thickness (phosphorus-to-phosphorus distance vs. hydrophobic thickness) affect the calculation outcome.
Ordering of lipid acyl chains. The profiles of the hydrogen order parameters (
SCH) along the
sn-1 chains of DPPC lipids were then calculated as previously described [
35].
Figure 5 shows that, at the higher temperature, the presence of DOPC mainly reduces the ordering of the deeper segments of the DPPC chains (C10–C14 carbons). The effect of the unsaturated lipid on
SCH in the upper regions of the mixed bilayer (i.e., the most ordered part of the bilayer) is negligible. On the contrary, at 295 K, the addition of DOPC has a significant impact on the ordering of DPPC hydrocarbon chains. The
SCH value strongly decreases as the mole fraction of DOPC increases. The greatest decrease in the ordering of the mixed DPPC/DOPC membrane is observed when
XDOPC = 0.3 is exceeded. Interestingly, the addition of 10 mol% DOPC increases the ordering of the DPPC hydrocarbon chains relative to the pure bilayer. This can be explained by the fact that the pure DPPC membrane in our simulations is in the ripple phase (
Figure 2), in which some of the lipids show high disorder. The introduction of DOPC causes a partial disappearance of the ripple, and thus an increase in the ordering of the DPPC alkyl chains.
Orientation of lipids in the mixed membrane. In order to describe the arrangement of the lipid molecules inside the mixed membranes, we calculated the tilt angle (
θc) between the acyl chains (both
sn-1 and
sn-2) and the normal to the membrane, and the tilt angle (
θh) between the headgroups of DPPC and DOPC lipids and the bilayer normal (see vector definitions in
Figure 1). The normalized probability distributions of the angles are shown in
Figure 6 and
Figure S5 for 295 K and 323 K, respectively.
In all membranes at the lower temperature, the
θc distributions for DOPC are wider than the corresponding distributions for DPPC, and the maxima are shifted toward larger angles (
Figure 6A,B). This is probably due to the bending of the hydrophobic chain caused by the presence of the double bond. The exception is the membrane with
XDOPC = 0.1, in which the DOPC molecules adopt an orientation similar to DPPC. In turn, in the lipid bilayers containing more DOPC (
XDOPC > 0.5), the average tilt angle of DPPC acyl chains becomes similar to that for DOPC. Interestingly, the DOPC acyl chains can adopt a nearly perpendicular orientation with respect to the bilayer normal (non-zero probability for angles above 80°,
Figure 6A), while the DPPC acyl chains do not exhibit such behavior. The
θh distributions (
Figure 6C,D) suggest that the most probable orientation of the headgroups of both DPPC and DOPC is their alignment almost parallel to the membrane surface, but the wide width of the distributions indicates a large degree of freedom in the arrangement of these groups.
Orientation of headgroups in the mixed membrane. We then focused on the arrangement of the headgroups relative to the entire lipid molecule. Two conformations with different headgroup orientations have been identified in the crystal structure of 1,2-dimyristoyl-
sn-glycero-3-phosphatidylcholine (DMPC) by X-ray scattering [
46]. Conformation A shows the headgroup pointing away from the rest of the molecule, while the headgroup in conformation B is folded back, resulting in a more compact structure. These conformations correspond to different orientations of the glycerol moiety in the lipid molecule. The situations where the CG-CG-CG-O dihedral angles (CG stands for carbon from the glycerol moiety) are in
gauche and
trans configurations are described by the structures A and B, respectively.
Figure 7 shows the two conformations adopted by the DPPC molecules in the gel state taken from the MD simulations of the DPPC system. The headgroups can be oriented either above or outside the lipid molecule; however, in both cases, they are arranged almost parallel to the membrane surface. The distributions of the
gauche and
trans configurations in the equimolar mixture of DPPC and DOPC at 295 K and 323 K are shown in
Figure 8. All distributions have three maxima; those at −60° and 60° correspond to the
gauche configurations of the CG-CG-CG-O dihedral angles (conformation A), while the maximum at about 180° is associated with the
trans configuration (conformation B). Our results show that both lipids have a lot of freedom to change the arrangement of the headgroups, but the
trans configuration (with the choline group located above the lipid molecule shielding the hydrophobic part from contact with water) dominates. The contribution of the
trans configuration decreases with temperature for both lipids, but this effect is stronger for DPPC.
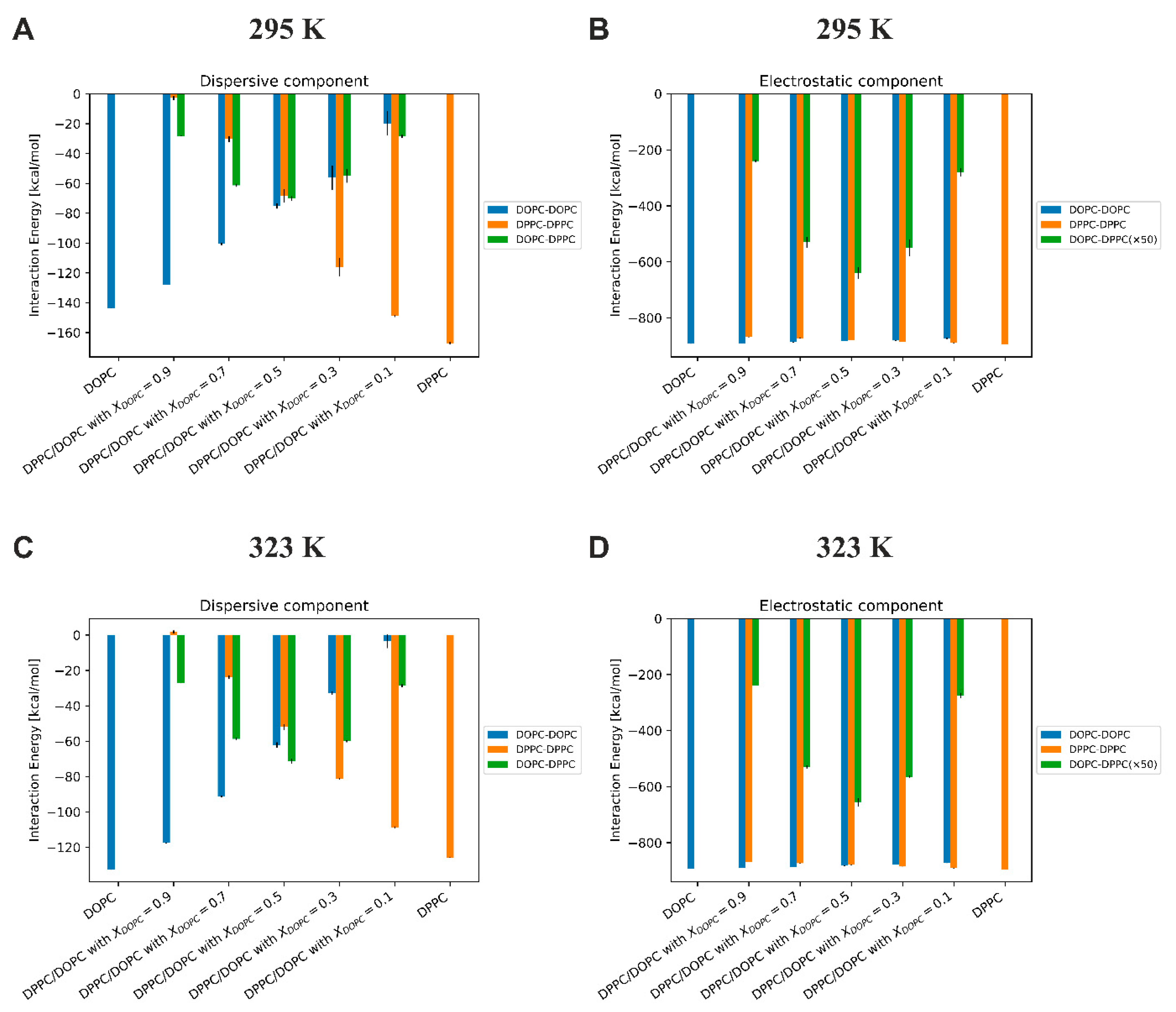
Energy of interactions between lipids. In order to quantify local interactions between lipids in the mixed DPPC/DOPC bilayers, we calculated the interaction energy between lipid molecules, distinguishing between electrostatic and dispersive components (
Figure 9). The dispersion energies, calculated from MD simulations, contain all the terms from the Lennard–Jones interaction potential, which includes both dispersive energy (always attractive) and Pauli repulsion (acting at much shorter interatomic distances). All energies were calculated as averages over time and the number of repeats.
Several conclusions can be drawn about the interactions between lipids in the mixed DOPC/DPPC membrane from
Figure 9: (i) the most important outcome is that temperature has only a slight influence on the value of both dispersive and Coulomb interactions; (ii) the dispersive interactions are much weaker compared to electrostatic interactions (as much as six times as in the case of the pure DOPC membrane); (iii) for the membrane with
XDOPC = 0.5, the energy of dispersion interactions between all lipid molecules is similar, while the Coulomb interactions for lipid like-pairs are more than 75 times stronger than for mixed-pairs.
The molecular organization of mixed DPPC/DOPC membrane. Our simulation results are consistent with experimental findings. The exception is the DPPC pure membrane simulated at 22 °C, which the simulations show is in the ripple phase, which is not in line with the DSC findings. All mixed DPPC/DOPC membranes simulated at temperatures above the DPPC phase transition were in the liquid phase; this means that the acyl chains of both lipids were highly disordered, as indicated by the order parameters (
Figure 5). In addition, the distributions of individual lipids in the bilayer were uniform, so the DPPC/DOPC membranes were single-phase systems.
At the lower temperature, for
XDOPC = 0.1, a densely packed membrane was observed with DOPC molecules dispersed evenly among DPPC lipids (
Figure 3), showing that this mixed membrane is a single-phase system. Importantly, the DOPC molecules adopt the orientation of the surrounding DPPC membrane (
Figure 6A), but their headgroups are located deeper compared to those of the saturated lipid. The APO value is much smaller compared to that in the pure DOPC membrane, showing that DOPC molecules adopt a more compact conformation in the mixed membrane. Interestingly, the introduction of a small amount of DOPC results in the disappearance of the ripple structure of the DPPC membrane. This is in good agreement with the DSC measurements, in which no pretransition was observed for all mixed systems, showing that no ripple phase is formed. Increasing the content of unsaturated lipid to
XDOPC = 0.3 causes its molecules to cluster together to form domains. Their thickness is much smaller compared to the membrane fragments composed mainly of DPPC. This indicates that a mixed membrane of this composition is a biphasic system, which is consistent with our DSC measurements. The introduction of DOPC into the DPPC membrane results in a significant broadening of the main transition peak, indicating a decrease in the size of cooperative units. DPPC-rich membrane fragments show high ordering, as demonstrated by the
SCH values; thus, they are in the gel state.
In the case of the bilayer with
XDOPC = 0.5, our simulations show a clear phase separation that leads to DPPC-rich domains suspended in a DOPC-rich phase. Interestingly, these domains are in register, which means that the domains in one leaflet are coupled to the same domains in the other leaflet. The image of the DPPC/DOPC membrane with
XDOPC = 0.5 obtained in our simulations (
Figure 3) is strongly supported by experimental results. X-ray diffraction measurements of an equimolar mixture of DOPC and DPPC at 30 °C showed that the two phospholipids are completely separated within the bilayer [
44]. In addition, the bilayers are completely in register, so that DPPC domains in one leaflet are invariably aligned with DPPC in the opposing leaflet and likewise with DOPC. The DPPC domains are in the gel phase, while the DOPC domains are characterized by the presence of disordered chains. The formation of domains in the DOPC/DPPC (1:1) membrane at room temperature has also been demonstrated directly by AFM visualizations [
47,
48]. AFM measures the lateral organization of lipid bilayers deposited on a substrate by imaging the topographical details of the membrane in the form of domain lateral size and height relative to surrounding lipids in the liquid state. However, AFM images showed that the substrate affects the domain organization. Micrometer-sized domains with rounded boundaries were formed in the DOPC/DPPC (1:1) bilayers on mica [
47]. In contrast, domains with irregular shape and sizes between 100 and 200 nm were observed for the DOPC/DPPC (1:1) bilayer prepared on glass [
48].
In the DPPC/DOPC membranes with a predominance of DOPCs (XDOPC ≥ 7), the phase separation gradually disappears. For the system with 70 mol% of the unsaturated lipid, we observed the formation of small aggregates, which is strongly supported by the DSC measurements. However, the properties of both lipids such as surface area per lipid (APP and APO) are comparable. This indicates that the boundary between the system being monophasic or biphasic is quite blurred. This can be the reason for the discrepancies in the phase diagrams determined by different experimental methods.
We also focused on the arrangement of the headgroups of both lipids in the mixed membranes. The results show that the phosphocholine groups have a lot of freedom in orientation but mainly make an angle in the range of 60–90° to the bilayer normal, which means that they are aligned almost parallel to the membrane surface. This orientation of the headgroups is almost independent of temperature and membrane composition. This agrees with neutron-scattering experiments, which report that the average orientation of the phosphocholine group of DPPC in the gel state, as well as in the liquid-crystalline state, is almost parallel to the membrane surface [
49]. Semchyschyn and Macdonald reported that for 1,2-dimyristoyl-
sn-glycero-3-phosphocholine (DMPC), the headgroup vector is about 0.45 nm in length and oriented at an angle of about 30° above the bilayer surface [
50]. The parallel alignment of the PC headgroup to the membrane surface can be achieved by two different conformations within the glycerol group. The
gauche configuration of the CG-CG-CG-O dihedral angle forces the phosphocholine group to point away from the rest of the molecule (conformer A), while in the
trans configurations the headgroup is located above the lipid molecule (conformer B). Our simulations indicate that in the gel phase, the contribution of conformer B is about 70% and decreases slightly when the membrane is in the liquid phase. Our simulation results are in line with experimental findings. Using two-dimensional
13C-
1H chemical shift correlation spectroscopy, it was shown that the arrangement of the DMPC molecule, in which its headgroup and the beginning of the
sn-2 chain bend toward each other, is significantly populated in the liquid bilayer, suggesting that conformer B dominates in the liquid-crystalline phase [
51].
The driving forces for phase separation in the DPPC/DOPC systems. The driving forces for phase separation between different lipids in the bilayer have long been identified as resulting from adverse interactions between lipid molecules. It is assumed that the miscibility of lipids depends mainly on the difference in the interaction energies among the like-pairs and mixed-pairs formed in the mixed membrane [
12,
28]. Lateral phase separation and domain formation can occur when like-pair interactions are stronger than those of the mixed-pairs. Interactions between lipid molecules occur both in their hydrophobic part and between their headgroups. The hydrophobic part consists of hydrocarbon chains, which can vary in length and degree of unsaturation. The forces acting in the hydrocarbon region of the bilayer are dominated by van der Waals attractions between the acyl chains. In contrast, lipid headgroups interact with each other through electrostatic and van der Waals interactions.
Our experimental results show that the free excess energy of mixing of DPPC and DOPC is positive at 22 °C, indicating that the miscibility of these lipids is thermodynamically unfavorable. The Gibbs–Helmholtz equation for the free excess Gibbs energy of mixing is giving by the following equation:
where ΔH
Exc, ΔS
Exc, and
T are the enthalpy and entropy of mixing and the temperature, respectively. Thus, the free excess energy of mixing is divided into an entropic component, mainly due to the loss of order of the hydrocarbon chains, and the enthalpic component, resulting mainly from interactions between lipid molecules. Our simulations showed that the electrostatic and dispersive components of the interaction energy in the DPPC/DOPC bilayer are almost constant with temperature change. In addition, the interaction energies for lipid like-pairs are more than 75 times stronger than for mixed-pairs. Therefore, we believe that the change in miscibility of these two lipids with increasing temperature is related to the entropic component. The entropy gain arises due to an increase in the disorder of the acyl chains above the transition temperature
Tm. As a result, the value of the
T ΔS
Exc term increases with increasing temperature, causing a decrease in ΔG
Exc, and making the miscibility in the DPPC/DOPC system thermodynamically favorable.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}