


A Practical Guide to Preparation and Applications of Giant Unilamellar Vesicles Formed via Centrifugation of Water-in-Oil Emulsion Droplets

Abstract
:1. Introduction
2. Practical Guide for the Protocol of W/O Emulsion Template Method with Centrifugation

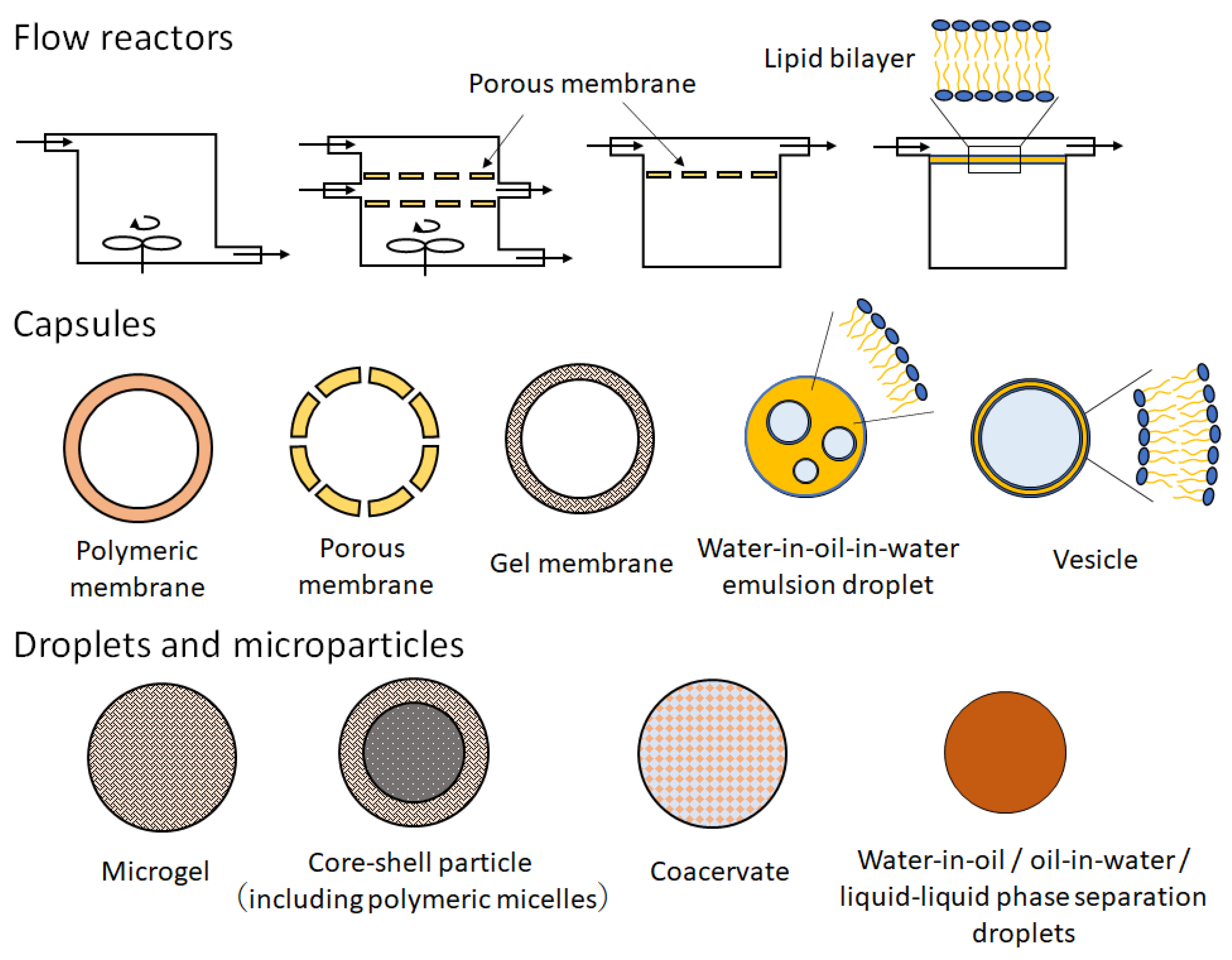{kind=link}
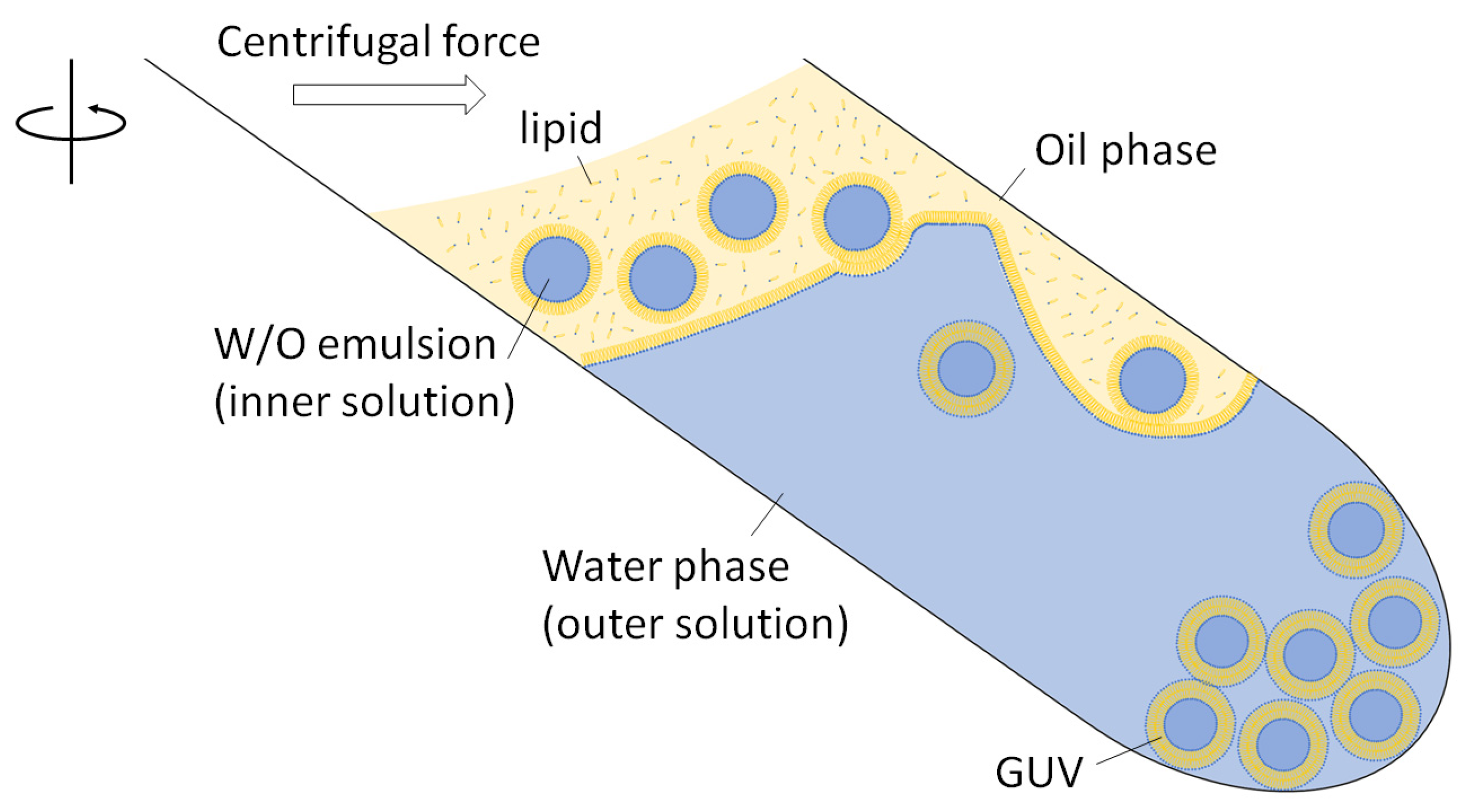{kind=link}
{kind=link}
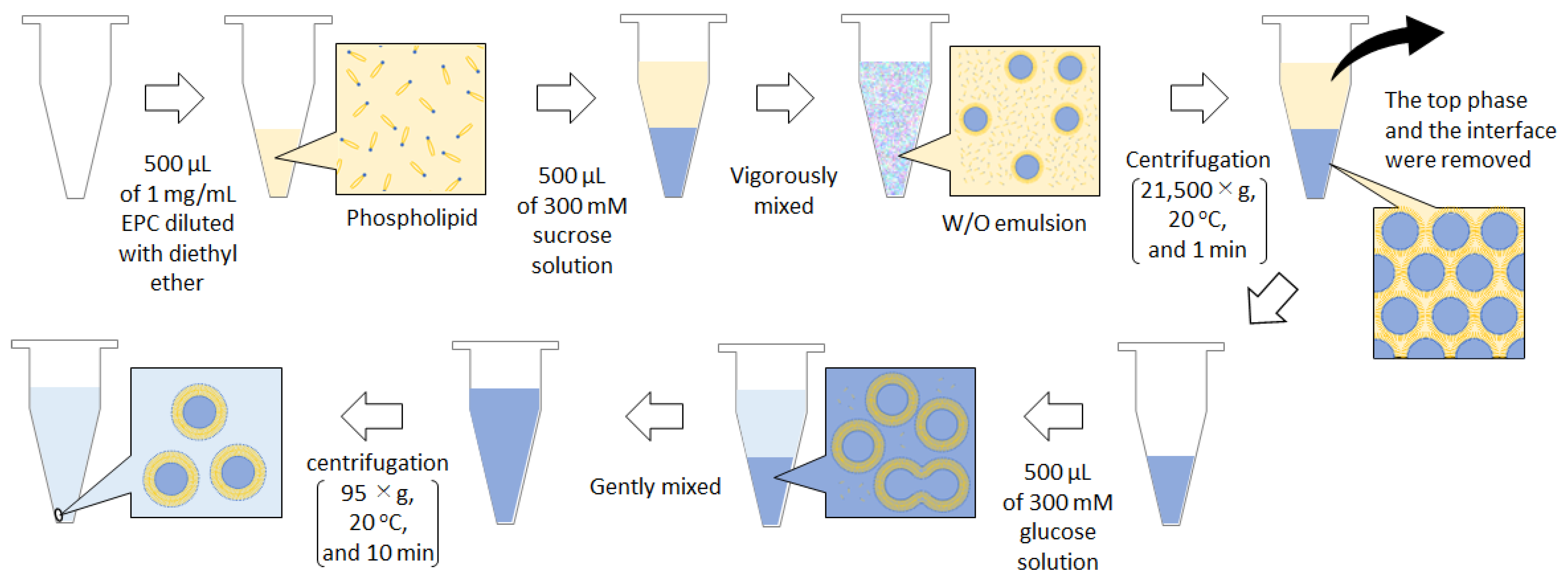{kind=link}
{kind=link}
{kind=link}
| Ref. | Oil | Inner and Outer Solutions 1 | Lipids 2 | Emulsifying Techniques | Container Volume | Centrifugation Conditions | Collecting GUVs |
|---|---|---|---|---|---|---|---|
| [21] | Dodecane and squalene | NaCl and Tris-HCl | Egg-PC, POPC, POPS, and DOPS | Filtration, gentle stirring, shear mixing, and ultrasonication | 50 mL | 5–10 min, unknown temp., and 120× g | – |
| [22] | Liquid paraffin | Glucose + sucrose, glucose, and Tris-HCl | DOPC | Shear mixing | 1.5 mL | 30 min, 20 °C, and 18,800× g | Opening hole |
| [29] | Mineral oil, 1-octanol, oleic acid, anisole, silicon oil, and squalene | Sucrose, glucose, Tris-HCl, HEPES-NaOH, and PBS | POPC | Rubbing on a rumble strip, vortex mixing, hand tapping, and ultrasonication | 300 μL 3 | 0.5–10 min, 4–37 °C, and 50–1200× g | N/A 4 |
| [30] | 1-Octanol, squalene, and liquid paraffin | Sucrose, glucose, and HEPES-NaOH | POPC + POPG | Rubbing on a rumble strip | 1.5 mL | 30 min, 20 °C, and 18,800× g | Opening hole or pipette aspiration |
| [31] | Liquid paraffin | Sucrose and NaCl + Tris-HCl | Egg-PC + DMPG + cholesterol | Hand tapping | 1.5 mL | 30 min, 20 °C, and 18,800× g | Opening hole |
| [32] | Liquid paraffin + squalene | Sucrose, glucose, and NaCl + Tris-HCl | Egg-PC + PEG5000-DOPE + cholesterol | Hand tapping | 1.5 mL | 30 min, 0 °C, and 18,800× g | Opening hole |
| [33] | Mineral oil | Sucrose, glucose, and HEPES-NaOH | POPC | Rubbing on a rumble strip, vortex mixing, and ultrasonication | 1.5 mL | 15 min, r.t. 5, and 16,100× g | Pipette aspiration |
| [36] | Mineral oil | Sucrose and glucose | POPC + PEG2000-DSPE | Ultrasonication and gentle stirring | 300 μL 3 | – | Pipette aspiration |
| [37] | Liquid paraffin + squalene | PIPES-HCl + MgCl2 + EGTA | DOPC + PEG2000-DSPE + cholesterol | Hand tapping | 200 μL | 30 min, 4 °C, and 18,800× g | Pipette aspiration |
| [38] | Liquid paraffin | Sucrose and glucose | POPC + cholesterol | Vortex mixing | 1.5 mL | 30 min, 4 °C, and 9000× g | Opening hole |
| [39] | Squalene | Sucrose, glucose, and Tris-HCl | Polyglycerol- polyricinoleate | Hand tapping | 15 mL | 30 min, r.t. 5, and 2250× g | Pipette aspiration |
| [40] | Mineral oil | Sucrose, glucose, and HEPES-NaOH | POPC + cholesterol | Vortex mixing | 15 mL | 10 min, 4 °C, and 100× g/400× g 6 | Pipette aspiration |
| [47] | Toluene | Sucrose + dextran and glucose | PEO-b-PBD, PEG-b-PA444, and PEG-b-PA6ester1 | Gentle pipetting | 1.5 mL 7 | 12 min, unknown temp., and 100× g | – |
| [49] | Toluene | Sucrose and glucose | PEO-b-PBD | Hand tapping | 1.5 mL 7 | 5 min, r.t. 5, and 500× g | – |
3. Construction and Application of GUV-Based Artificial Cells
3.1. Repetitive Transformation via Protein Polymerization and Depolymerization
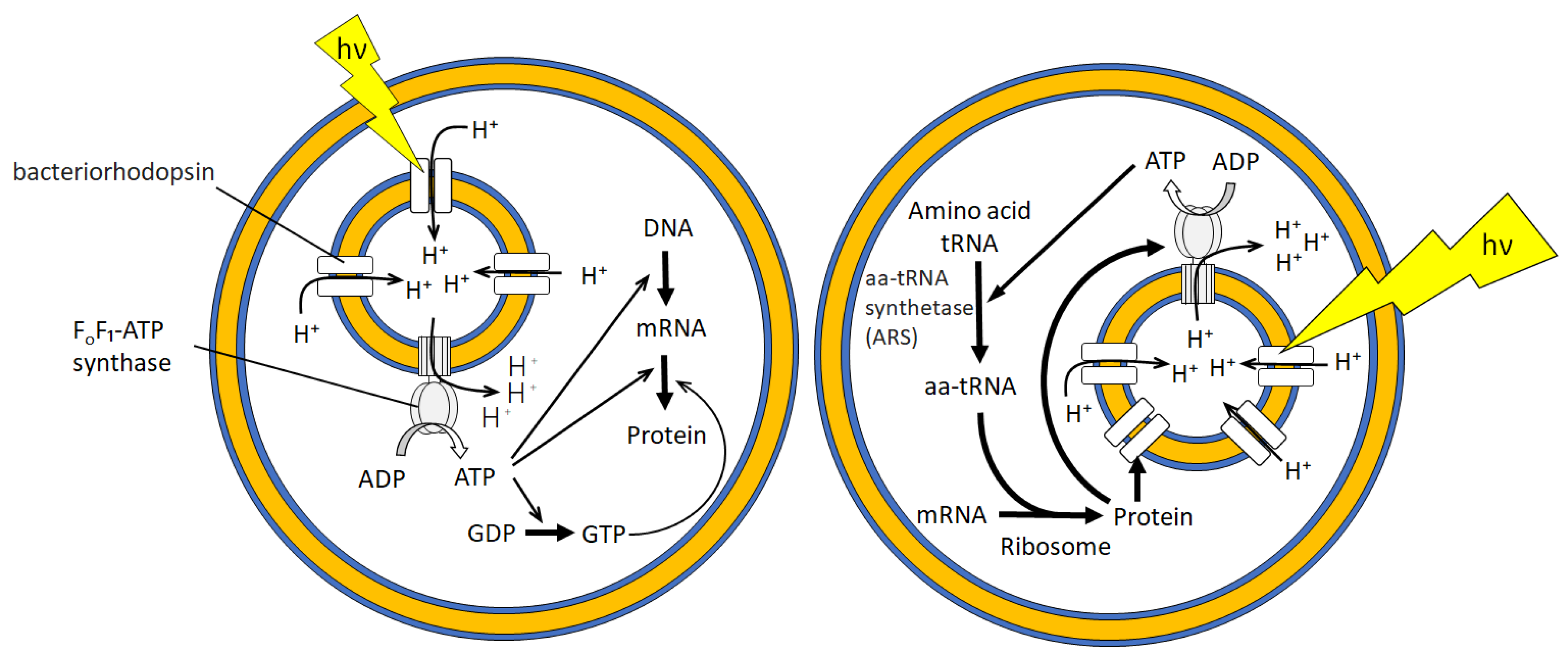
3.2. Bioreactor for Cell-Free Protein Synthesis
3.3. DNA Nanotechnology Platform
3.4. Fusion and Fission Assisted by a Microfabricated Platform
3.5. Encapsulation of Micrometer-Sized Particles
3.6. Intercellular Communication Model
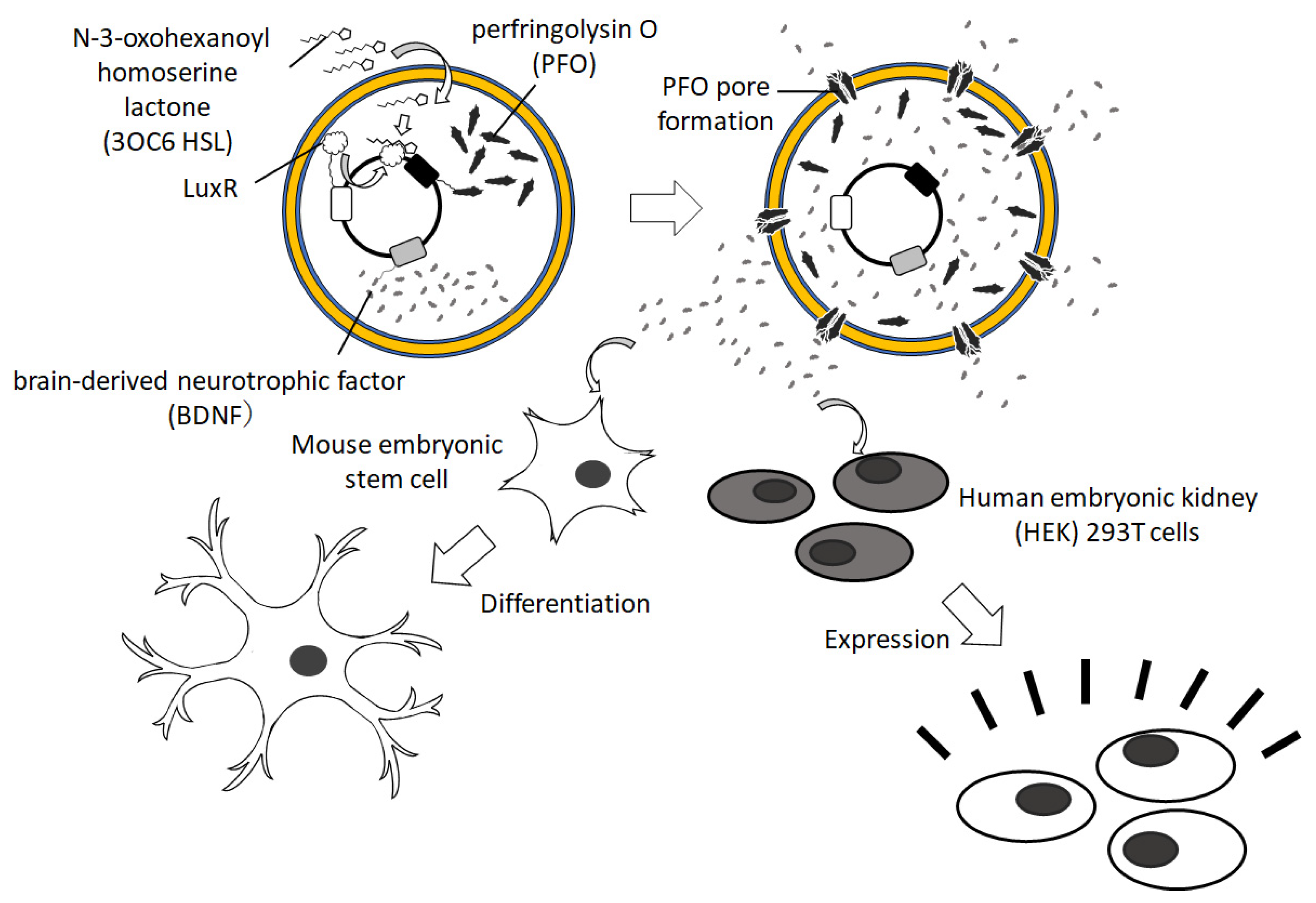
3.7. Stimulus-Responsive Sensor and Releaser
4. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimova, R.; Stano, P.; Marques, C.M.; Walde, P.B. Preparation methods for giant unilamellar vesicles. In The Giant Vesicle Book; Dimova, R., Marques, C.M., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 3–20. [Google Scholar]
- Reeves, J.P.; Dowben, R.M. Formation and properties of thin-walled phospholipid vesicles. J. Cell Physiol. 1969, 73, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.M.S. Artificial cells with emphasis on bioencapsulation in biotechnology. Biotech. Ann. Rev. 1995, 1, 267–295. [Google Scholar]
- Jiang, W.; Wu, Z.; Gao, Z.; Wan, M.; Zhou, M.; Mao, C.; Shen, J. Artificial Cells: Past, Present and Future. ACS Nano 2022, 16, 15705–15733. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cho, E. Compartmentalizing Cell-Free Systems: Toward Creating Life-Like Artificial Cells and Beyond. ACS Synth. Biol. 2020, 9, 2881–2901. [Google Scholar]
- Jeong, S.; Nguyen, H.T.; Kim, C.H.; Ly, M.N.; Shin, K. Toward Artificial Cells: Novel Advances in Energy Conversion and Cellular Motility. Adv. Funct. Mater. 2020, 30, 1907182. [Google Scholar] [CrossRef]
- Matosevic, S. Synthesizing artificial cells from giant unilamellar vesicles: State-of-the art in the development of microfluidic technology. BioEssays 2012, 34, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Buddingh’, B.C.; van Hest, J.C.M. Artificial Cells: Synthetic Compartments with Life-like Functionality and Adaptivity. Acc. Chem. Res. 2017, 50, 769–777. [Google Scholar] [CrossRef]
- Ai, Y.; Xie, R.; Xiong, J.; Liang, Q. Microfluidics for Biosynthesizing: From Droplets and Vesicles to Artificial Cells. Small 2020, 16, 1903940. [Google Scholar] [CrossRef]
- Lo, C.H.; Zeng, J. Application of polymersomes in membrane protein study and drug discovery: Progress, strategies, and perspectives. Bioeng. Transl. Med. 2023, 8, e10350. [Google Scholar] [CrossRef]
- Messager, L.; Gaitzsch, J.; Chierico, L.; Battaglia, G. Novel aspects of encapsulation and delivery using polymersomes. Curr. Opin. Pharmacol. 2014, 18, 104–111. [Google Scholar] [CrossRef]
- Le Meins, J.-F.; Sandre, O.; Lecommandoux, S. Recent trends in the tuning of polymersomes’ membrane properties. Eur. Phys. J. E 2011, 34, 14. [Google Scholar] [CrossRef] [PubMed]
- Méléard, P.; Gerbeaud, C.; Pott, T.; Fernandez-Puente, L.; Bivas, I.; Mitov, M.D.; Dufourcq, J.; Bothorel, P. Bending elasticities of model membranes: Influences of temperature and sterol content. Biophys. J. 1997, 72, 2616–2629. [Google Scholar] [CrossRef] [PubMed]
- Angelova, M.I.; Dimitrov, D.S. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Tsumoto, K.; Matsuo, H.; Tomita, M.; Yoshimura, T. Efficient formation of giant liposomes through the gentle hydration of phosphatidylcholine films doped with sugar. Colloids Surf. B Biointerfaces 2009, 68, 98–105. [Google Scholar] [CrossRef]
- Yamashita, Y.; Oka, M.; Tanaka, T.; Yamazaki, M. A new method for the preparation of giant liposomes in high salt concentrations and growth of protein microcrystals in them. Biochim. Biophys. Acta-Biomembr. 2002, 1561, 129–134. [Google Scholar] [CrossRef]
- Funakoshi, K.; Suzuki, H.; Takeuchi, S. Formation of giant lipid vesiclelike compartments from a planar lipid membrane by a pulsed jet flow. J. Am. Chem. Soc. 2007, 129, 12608–12609. [Google Scholar] [CrossRef]
- Stachowiak, J.C.; Richmond, D.L.; Li, T.H.; Liu, A.P.; Parekh, S.H.; Fletcher, D.A. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc. Natl. Acad. Sci. USA 2008, 105, 4697–4702. [Google Scholar] [CrossRef]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant vesicles: Preparations and applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef]
- Moscho, A.; Orwar, O.W.E.; Chiu, D.T.; Modi, B.P.; Zare, R.N. Rapid preparation of giant unilamellar vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 11443–11447. [Google Scholar] [CrossRef]
- Pautot, S.; Frisken, B.J.; Weitz, D.A. Production of Unilamellar Vesicles Using an Inverted Emulsion. Langmuir 2003, 19, 2870–2879. [Google Scholar] [CrossRef]
- Natsume, Y.; Toyota, T. Giant vesicles containing microspheres with high volume fraction prepared by water-in-oil emulsion centrifugation. Chem. Lett. 2013, 42, 295–297. [Google Scholar] [CrossRef]
- Faizi, H.A.; Tsui, A.; Dimova, R.; Vlahovska, P.M. Bending rigidity, capacitance, and shear viscosity of giant vesicle membranes prepared by spontaneous swelling, electroformation, gel-assisted, and phase transfer methods: A comparative study. Langmuir 2022, 38, 10548–10557. [Google Scholar] [CrossRef]
- Abkarian, M.; Loiseau, E.; Massiera, G. Continuous droplet interface crossing encapsulation (cDICE) for high throughput monodisperse vesicle design. Soft Matter 2011, 7, 4610–4614. [Google Scholar] [CrossRef]
- Morita, M.; Onoe, H.; Yanagisawa, M.; Ito, H.; Ichikawa, M.; Fujiwara, K.; Saito, H.; Takinoue, M. Droplet-Shooting and Size-Filtration (DSSF) Method for Synthesis of Cell-Sized Liposomes with Controlled Lipid Compositions. ChemBioChem 2015, 16, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Siddharth, D.; Dekker, C. On-chip microfluidic production of cell-sized liposomes. Nat. Protoc. 2018, 13, 856–874. [Google Scholar]
- Yandrapalli, N.; Petit, J.; Bäumchen, O.; Robinson, R. Surfactant-free production of biomimetic giant unilamellar vesicles using PDMS-based microfluidics. Commun. Chem. 2021, 4, 100. [Google Scholar] [CrossRef] [PubMed]
- Ushiyama, R.; Koiwai, K.; Suzuki, H. Plug-and-play microfluidic production of monodisperse giant unilamellar vesicles using droplet transfer across Water-Oil interface. Sens. Actuators B Chem. 2022, 355, 131281. [Google Scholar] [CrossRef]
- Moga, A.; Yandrapalli, N.; Dimova, R.; Robinson, T. Optimization of the inverted emulsion method for high-yield production of biomimetic giant unilamellar vesicles. ChemBioChem 2019, 20, 2674–2682. [Google Scholar] [CrossRef]
- Toyota, T.; Zhang, Y. Effect of an Oil Medium on Giant Vesicles Prepared with Water-in-Oil Emulsion. Bunseki Kagaku 2022, 71, 83–89. [Google Scholar] [CrossRef]
- Toyota, T.; Ohguri, N.; Maruyama, K.; Fujinami, M.; Saga, T.; Aoki, I. Giant vesicles containing superparamagnetic iron oxide as biodegradable cell-tracking MRI probes. Anal. Chem. 2012, 84, 3952–3957. [Google Scholar] [CrossRef]
- Hamada, S.; Tabuchi, M.; Toyota, T.; Sakurai, T.; Hosoi, T.; Nomoto, T.; Nakatani, K.; Fujinami, M.; Kanzaki, R. Giant vesicles functionally expressing membrane receptors for an insect pheromone. Chem. Commun. 2014, 50, 2958–2961. [Google Scholar] [CrossRef] [PubMed]
- Matsushita-Ishiodori, Y.; Hanczyc, M.M.; Wang, A.; Szostak, J.W.; Yomo, T. Using imaging flow cytometry to quantify and optimize giant vesicle production by water-in-oil emulsion transfer methods. Langmuir 2019, 35, 2375–2382. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, M. The characterization of acoustic cavitation bubbles—An overview. Ultrason Sonochem. 2011, 18, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Hadorn, M.; Eggenberger, P. Towards Personalized Drug Delivery-Preparation of an Encapsulated Multicompartment System. Towards Personalized Drug Delivery. In Proceedings of the Third International Conference on Biomedical Electronics and Devices, Valencia, Spain, 20–23 January 2010; pp. 5–12. [Google Scholar]
- Hayashi, M.; Nishiyama, M.; Kazayama, Y.; Toyota, T.; Harada, Y.; Takiguchi, K. Reversible morphological control of tubulin-encapsulating giant liposomes by hydrostatic pressure. Langmuir 2016, 32, 3794–3802. [Google Scholar] [CrossRef]
- Fujii, S.; Matsuura, T.; Sunami, T.; Nishikawa, T.; Kazuta, Y.; Yomo, T. Liposome display for in vitro selection and evolution of membrane proteins. Nat. Protoc. 2014, 9, 1578–1591. [Google Scholar] [CrossRef]
- Yahagi, R.; Yoshida, K.; Zhang, Y.; Ebata, M.; Toyota, T.; Yamaguchi, T.; Hayashi, H. Destruction of giant cluster-like vesicles by an ultrasonically activated device. Jpn. J. Appl. Phys. 2016, 55, 07KF21. [Google Scholar] [CrossRef]
- Adir, O.; Sharf-Pauker, N.; Chen, G.; Kaduri, M.; Krinsky, N.; Shainsky-Roitman, J.; Shklover, J.; Schroeder, A. Preparing Protein Producing Synthetic Cells using Cell Free Bacterial Extracts, Liposomes and Emulsion Transfer. J. Vis. Exp. 2020, 158, e60829. [Google Scholar]
- Toyota, T.; Zhang, Y. Identifying and Manipulating Giant Vesicles: Review of Recent Approaches. Micromachines 2022, 13, 644. [Google Scholar] [CrossRef]
- Sackmann, E. Supported membranes: Scientific and practical applications. Science 1996, 271, 43–48. [Google Scholar] [CrossRef]
- Gruhn, T.; Franke, T.; Dimova, R.; Lipowsky, R. Novel method for measuring the adhesion energy of vesicles. Langmuir 2007, 23, 5423–5429. [Google Scholar] [CrossRef]
- Rädler, J.O.; Feder, T.J.; Strey, H.H.; Sackmann, E. Fluctuation analysis of tension-controlled undulation forces between giant vesicles and solid substrates. Phys. Rev. E 1995, 51, 4526. [Google Scholar] [CrossRef] [PubMed]
- Lira, R.B.; Steinkühler, J.; Knorr, R.L.; Dimova, R.; Riske, K.A. Posing for a picture: Vesicle immobilization in agarose gel. Sci. Rep. 2016, 6, 25254. [Google Scholar] [CrossRef]
- Tsumoto, K.; Oohashi, M.; Tomita, M. Monitoring of membrane collapse and enzymatic reaction with single giant liposomes embedded in agarose gel. Colloid Polym. Sci. 2011, 289, 1337–1346. [Google Scholar] [CrossRef]
- Mabrouk, E.; Cuvelier, D.; Pontani, L.-L.; Xu, B.; Lévy, D.; Keller, P.; Brochard-Wyart, F.; Nassoy, P.; Li, M.-H. Formation and material properties of giant liquid crystal polymersomes. Soft Matter 2009, 5, 1870–1878. [Google Scholar] [CrossRef]
- Mabrouk, E.; Cuvelier, D.; Brochard-Wyart, F.; Nassoy, P.; Li, M.-H. Bursting of sensitive polymersomes induced by curling. Proc. Natl. Acad. Sci. USA 2009, 106, 7294–7298. [Google Scholar] [CrossRef] [PubMed]
- Marguet, M.; Sandre, O.; Lecommandoux, S. Polymersomes in “Gelly” Polymersomes: Toward Structural Cell Mimicry. Langmuir 2012, 28, 2035–2043. [Google Scholar] [CrossRef]
- Marguet, M.; Edembe, L.; Lecommandoux, S. Polymersomes in Polymersomes: Multiple Loading and Permeability Control. Angew. Chem. 2012, 124, 1199–1202. [Google Scholar] [CrossRef]
- Peyret, A.; Ibarboure, E.; Pippa, N.; Lecommandoux, S. Liposomes in Polymersomes: Multicompartment System with Temperature-Triggered Release. Langmuir 2017, 33, 7079–7085. [Google Scholar] [CrossRef]
- Chiba, M.; Miyazaki, M.; Ishiwata, S. Quantitative analysis of the lamellarity of giant liposomes prepared by the inverted emulsion method. Biophys. J. 2014, 107, 346–354. [Google Scholar] [CrossRef]
- Yamazaki, M. Advances in Planar Lipid Bilayers and Liposomes, 1st ed.; Academic Press: Cambridge, MA, USA, 2008; Chapter 5; pp. 121–142. ISBN 978-00-8092-162-4. [Google Scholar]
- Elani, Y.; Law, R.; Ces, O. Vesicle-based artificial cells as chemical microreactors with spatially segregated reaction pathways. Nat. Commun. 2014, 5, 5305. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, H.; Osaki, T.; Takeuchi, S.; Toyota, T. Perfusion Chamber for Observing a Liposome-Based Cell Model Prepared by a Water-in-Oil Emulsion Transfer Method. ACS Omega 2020, 5, 19429–19436. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Kawano, R.; Osaki, T.; Akiyoshi, K.; Takeuchi, S. Cell-sized asymmetric lipid vesicles facilitate the investigation of asymmetric membranes. Nat. Chem. 2016, 8, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Ma, Y.; Kodali, D.R.; Liang, J.; Davis, H.T. Ternary phase diagram of soybean phosphatidylcholine-water-soybean oil and its application to the water degumming process. J. Am. Oil Chem. Soc. 2003, 80, 383–388. [Google Scholar] [CrossRef]
- Shimane, Y.; Kuruma, Y. Rapid and Facile Preparation of Giant Vesicles by the Droplet Transfer Method for Artificial Cell Construction. Front. Bioeng. Biotechnol. 2022, 10, 873854. [Google Scholar] [CrossRef]
- Tsumoto, K.; Hayashi, Y.; Tabata, J.; Tomita, M. A reverse-phase method revisited: Rapid high-yield preparation of giant unilamellar vesicles (GUVs) using emulsification followed by centrifugation. Colloids Surf. A Physicochem. Eng. Asp. 2018, 546, 74–82. [Google Scholar] [CrossRef]
- Lach, S.; Yoonm, S.M.; Grzybowski, B.A. Tactic, reactive, and functional droplets outside of equilibrium. Chem. Soc. Rev. 2016, 45, 4766–4796. [Google Scholar] [CrossRef]
- Sato, Y.; Takinoue, M. Creation of artificial cell-like structures promoted by microfluidics technologies. Micromachines 2019, 10, 216. [Google Scholar] [CrossRef]
- Robinson, T. Microfluidic handling and analysis of giant vesicles for use as artificial cells: A review. Adv. Biosyst. 2019, 3, 1800318. [Google Scholar] [CrossRef]
- Lyu, Y.; Peng, R.; Liu, H.; Kuai, H.; Mo, L.; Han, D.; Li, J.; Tan, W. Protocells programmed through artificial reaction networks. Chem. Sci. 2020, 11, 631–642. [Google Scholar] [CrossRef]
- Olivi, L.; Berger, M.; Creyghton, R.N.P.; De Franceschi, N.; Dekker, C.; Mulder, B.M.; Claassens, N.J.; ten Wolde, P.R.; van der Oost, J. Towards a synthetic cell cycle. Nat. Commun. 2021, 12, 4531. [Google Scholar] [CrossRef]
- Wang, X.; Tian, L.; Du, H.; Li, M.; Mu, W.; Drinkwater, B.W.; Han, X.; Mann, S. Chemical communication in spatially organized protocell colonies and protocell/living cell micro-arrays. Chem. Sci. 2019, 10, 9446–9453. [Google Scholar] [CrossRef]
- Li, Q.; Li, S.; Zhang, X.; Xu, W.; Han, X. Programmed magnetic manipulation of vesicles into spatially coded prototissue architectures arrays. Nat. Commun. 2020, 11, 232. [Google Scholar] [CrossRef] [PubMed]
- Villar, G.; Graham, A.D.; Bayley, H. A tissue-like printed material. Science 2013, 340, 48–52. [Google Scholar] [CrossRef]
- Mukwaya, V.; Mann, S.; Dou, H. Chemical communication at the synthetic cell/living cell interface. Commun. Chem. 2021, 4, 161. [Google Scholar] [CrossRef]
- Murata, S.; Toyota, T.; Nomura, S.M.; Nakakuki, T.; Kuzuya, A. Molecular Cybernetics: Challenges toward Cellular Chemical Artificial Intelligence. Adv. Funct. Mater. 2022, 32, 2201866. [Google Scholar] [CrossRef]
- Takiguchi, K.; Yamada, A.; Negishi, M.; Tanaka-Takiguchi, Y.; Yoshikawa, K. Entrapping desired amounts of actin filaments and molecular motor proteins in giant liposomes. Langmuir 2008, 24, 11323–11326. [Google Scholar] [CrossRef]
- Tanaka, S.; Takiguchi, K.; Hayashi, M. Repetitive stretching of giant liposomes utilizing the nematic alignment of confined actin. Commun. Phys. 2018, 1, 18. [Google Scholar] [CrossRef]
- Nishimura, K.; Matsuura, T.; Nishimura, K.; Sunami, T.; Suzuki, H.; Yomo, T. Cell-free protein synthesis inside giant unilamellar vesicles analyzed by flow cytometry. Langmuir 2012, 28, 8426–8432. [Google Scholar] [CrossRef]
- Noireaux, V.; Libchaber, A. A vesicle bioreactor as a step toward an artificial cell assembly. Proc. Natl. Acad. Sci. USA 2004, 101, 17669–17674. [Google Scholar] [CrossRef]
- Berhanu, S.; Ueda, T.; Kuruma, Y. Artificial photosynthetic cell producing energy for protein synthesis. Nat. Commun. 2019, 10, 1325. [Google Scholar] [CrossRef]
- Sato, Y.; Komiya, K.; Kawamata, I.; Murata, S.; Nomura, S.I.M. Isothermal amplification of specific DNA molecules inside giant unilamellar vesicles. Chem. Commun. 2019, 55, 9084–9087. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hiratsuka, Y.; Kawamata, I.; Murata, S.; Nomura, S.I.M. Micrometer-sized molecular robot changes its shape in response to signal molecules. Sci. Robot. 2017, 2, eaal3735. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, H.; Tsuda, S.; Suzuki, H.; Yomo, T. Liposome-based liquid handling platform featuring addition, mixing, and aliquoting of femtoliter volumes. PLoS ONE 2014, 9, e101820. [Google Scholar] [CrossRef]
- Terasawa, H.; Nishimura, K.; Suzuki, H.; Yomo, T. Coupling of the fusion and budding of giant phospholipid vesicles containing macromolecules. Proc. Natl. Acad. Sci. USA 2012, 109, 5942–5947. [Google Scholar] [CrossRef]
- Schmid, Y.R.F.; Scheller, L.; Buchmann, S.; Dittrich, P.S. Calcium-Mediated Liposome Fusion to Engineer Giant Lipid Vesicles with Cytosolic Proteins and Reconstituted Mammalian Proteins. Adv. Biosyst. 2020, 4, 2000153. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.C.; Ogura, T.; Fujiwara, K.; Murata, S.; Nomura, S.M. Introducing micrometer-sized artificial objects into live cells: A method for cell–giant unilamellar vesicle electrofusion. PLoS ONE 2014, 9, e106853. [Google Scholar] [CrossRef] [PubMed]
- Natsume, Y.; Noguchi, E.; Kurihara, K. Spontaneous localization of particles in giant vesicles owing to depletion force. J. Phys. Soc. Jpn. 2019, 88, 033001. [Google Scholar] [CrossRef]
- Natsume, Y.; Toyota, T. Asymmetrical polyhedral configuration of giant vesicles induced by orderly array of encapsulated colloidal particles. PLoS ONE 2016, 11, e0146683. [Google Scholar] [CrossRef]
- Suzuki, K.; Machida, K.; Yamaguchi, K.; Sugawara, T. Photo-triggered recognition between host and guest compounds in a giant vesicle encapsulating photo-pierceable vesicles. Chem. Phys. Lipids 2018, 210, 70–75. [Google Scholar] [CrossRef]
- Bolognesi, G.; Friddin, M.S.; Salehi-Reyhani, A.; Barlow, N.E.; Brooks, N.J.; Ces, O.; Elani, Y. Sculpting and fusing biomimetic vesicle networks using optical tweezers. Nat. Commun. 2018, 9, 1882. [Google Scholar] [CrossRef] [PubMed]
- Buddingh’, B.C.; Elzinga, J.; van Hest, J.C. Intercellular communication between artificial cells by allosteric amplification of a molecular signal. Nat. Commun. 2020, 11, 1652. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Toyota, T.; Goto, S.; Ooishi, A.; Gao, T.; Ee, L.B.; Hatayama, H.; Nomoto, T.; Fujinami, M.; Matsubara, H. Development of a non-blurring, dual-imaging tissue marker for gastrointestinal tumor localization. Surg. Endosc. 2015, 29, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Hatayama, H.; Toyota, T.; Hayashi, H.; Nomoto, T.; Fujinami, M. Application of a novel near infrared-fluorescence giant vesicle-and polymerasome-based tissue marker for endoscopic and laparoscopic navigation. Anal. Sci. 2014, 30, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Kitaguchi, T.; Ohmuro-Matsuyama, Y.; Seah, T.; Ghadessy, J.F.; Hoon, S.; Ueda, H. Transmembrane signaling on a protocell: Creation of receptor-enzyme chimeras for immunodetection of specific antibodies and antigens. Sci. Rep. 2019, 9, 18189. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Lu, P.; Gamal El-Din, T.M.; Pei, X.Y.; Johnson, M.C.; Uyeda, A.; Bick, M.J.; Xu, Q.; Jiang, D.; Bai, H.; et al. Computational design of transmembrane pores. Nature 2020, 585, 129–134. [Google Scholar] [CrossRef]
- Krinsky, N.; Kaduri, M.; Zinger, A.; Shainsky-Roitman, J.; Goldfeder, M.; Benhar, I.; Hershkovitz, D.; Schroeder, A. Synthetic Cells Synthesize Therapeutic Proteins inside Tumors. Adv. Healthc. Mater. 2018, 7, e1701163. [Google Scholar] [CrossRef]
- Toparlak, Ö.D.; Zasso, J.; Bridi, S.; Serra, M.D.; Macchi, P.; Conti, L.; Baudet, M.; Mansy, S. Artificial cells drive neural differentiation. Sci. Adv. 2020, 6, eabb4920. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Obuchi, H.; Toyota, T. A Practical Guide to Preparation and Applications of Giant Unilamellar Vesicles Formed via Centrifugation of Water-in-Oil Emulsion Droplets. Membranes 2023, 13, 440. https://doi.org/10.3390/membranes13040440
Zhang Y, Obuchi H, Toyota T. A Practical Guide to Preparation and Applications of Giant Unilamellar Vesicles Formed via Centrifugation of Water-in-Oil Emulsion Droplets. Membranes. 2023; 13(4):440. https://doi.org/10.3390/membranes13040440
Chicago/Turabian StyleZhang, Yiting, Haruto Obuchi, and Taro Toyota. 2023. "A Practical Guide to Preparation and Applications of Giant Unilamellar Vesicles Formed via Centrifugation of Water-in-Oil Emulsion Droplets" Membranes 13, no. 4: 440. https://doi.org/10.3390/membranes13040440
APA StyleZhang, Y., Obuchi, H., & Toyota, T. (2023). A Practical Guide to Preparation and Applications of Giant Unilamellar Vesicles Formed via Centrifugation of Water-in-Oil Emulsion Droplets. Membranes, 13(4), 440. https://doi.org/10.3390/membranes13040440