Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics


 ,
,  ,
,  ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Methods
3. Results and Discussion
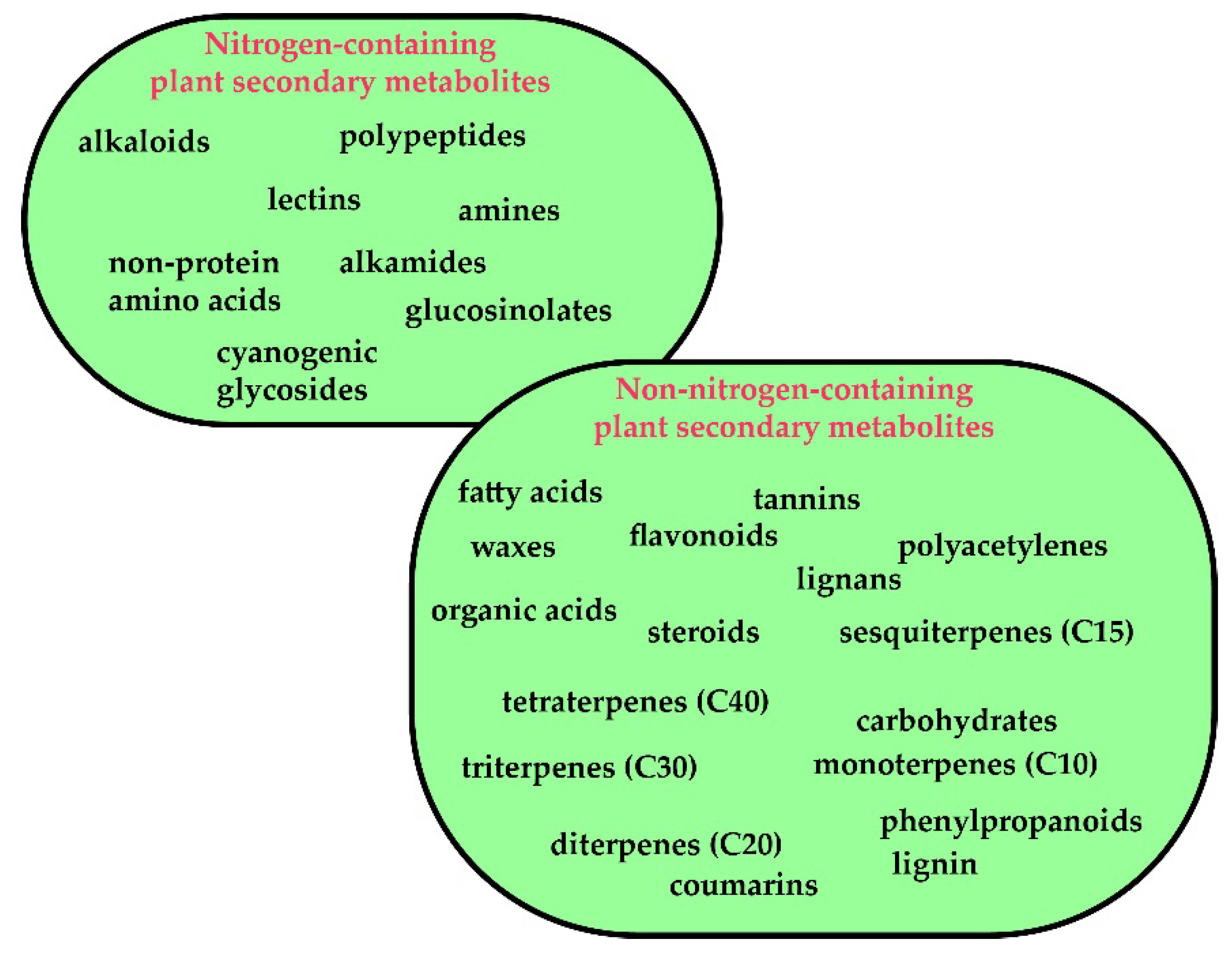
3.1. Plants Secondary Metabolites: A Brief Overview
3.1.1. Background
3.1.2. Ethnopharmacological Relevance of Natural Compounds for NDDs
3.2. Pharmacological Activities of Plants Secondary Metabolites on Neurodegenerative Disorders (NDDs): in vitro and in vivo Studies.
3.2.1. Preventing Protein Misfolding and Aggregation
3.2.2. Antioxidant Activity
3.2.3. Anti-inflammatory Activity
3.2.4. Antiapoptotic and Neurotrophic Activities
3.2.5. Acetylcholinesterase Inhibition Activity
3.2.6. Monoamine Oxidase Inhibitors (MAOs)
3.2.7. Antithrombotic Activity
4. Future Perspectives
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Dugger, B.N.; Dickson, D.W. Pathology of neurodegenerative diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement (Chinese edition). J. Chin. Integr. Med. 2009, 7, 889–896. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Sharifi-Rad, M.; Salehi, B.; Iriti, M.; Roointan, A.; Mnayer, D.; Soltani-Nejad, A.; Afshari, A. In vitro and in vivo assessment of free radical scavenging and antioxidant activities of Veronica persica Poir. Cell. Mol. Biol. 2018, 64, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Melgar-Lalanne, G.; Hernández-Álvarez, A.J.; Taheri, Y.; Shaheen, S.; Kregiel, D.; Antolak, H.; Pawlikowska, E.; Brdar-Jokanović, M.; Rajkovic, J.; et al. Malva species: Insights on its chemical composition towards pharmacological applications. Phytother. Res. 2019, 34, 546–567. [Google Scholar] [CrossRef]
- Patti, F.; Taheri, Y.; Sharifi-Rad, J.; Martorell, M.; C Cho, W.; Pezzani, R. Erythrina suberosa: Ethnopharmacology, Phytochemistry and Biological Activities. Medicines 2019, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Rad, J.; Ayatollahi, S.A.; Varoni, E.M.; Salehi, B.; Kobarfard, F.; Sharifi-Rad, M.; Iriti, M.; Sharifi-Rad, M. Chemical composition and functional properties of essential oils from Nepeta schiraziana Boiss. Farmacia 2017, 65, 802–812. [Google Scholar]
- Salehi, B.; Stojanović-Radić, Z.; Matejić, J.; Sharopov, F.; Antolak, H.; Kręgiel, D.; Sen, S.; Sharifi-Rad, M.; Acharya, K.; Sharifi-Rad, R.; et al. Plants of genus Mentha: From farm to food factory. Plants 2018, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Wink, M. Introduction: Biochemistry, Physiology and Ecological Functions of Secondary Metabolites. In Annual Plant Reviews; Wink, M., Ed.; Wiley-Blackwell: Oxford, UK, 2010; Volume 40. [Google Scholar]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural Products (Secondary Metabolites). In Biochemistry & Molecular Biology of Plants; Buchanan, B.B., Gruissem, W., Jones, R.L., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Salehi, B.; Albayrak, S.; Antolak, H.; Kręgiel, D.; Pawlikowska, E.; Sharifi-Rad, M.; Uprety, Y.; Fokou, P.V.T.; Yousef, Z.; Zakaria, Z.A.; et al. Aloe genus plants: From farm to food applications and phytopharmacotherapy. Int. J. Mol. Sci. 2018, 19, 2843. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Zucca, P.; Orhan, I.E.; Azzini, E.; Adetunji, C.O.; Mohammed, S.A.; Banerjee, S.K.; Sharopov, F.; Rigano, D.; Sharifi-Rad, J.; et al. Allicin and health: A comprehensive review. Trends Food Sci. Technol. 2019, 86, 502–516. [Google Scholar] [CrossRef]
- Salehi, B.; Selamoglu, Z.; Sener, B.; Kilic, M.; Kumar Jugran, A.; de Tommasi, N.; Sinisgalli, C.; Milella, L.; Rajkovic, J.; Morais-Braga, F.B. Berberis Plants—Drifting from Farm to Food Applications, Phytotherapy, and Phytopharmacology. Foods 2019, 8, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Capanoglu, E.; Adrar, N.; Catalkaya, G.; Shaheen, S.; Jaffer, M.; Giri, L.; Suyal, R.; Jugran, A.K.; Calina, D. Cucurbits plants: A key emphasis to its pharmacological potential. Molecules 2019, 24, 1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Krochmal-Marczak, B.; Skiba, D.; Patra, J.K.; Das, S.K.; Das, G.; Popović-Djordjević, J.B.; Kostić, A.Ž.; Anil Kumar, N.V.; Tripathi, A. Convolvulus plant—A comprehensive review from phytochemical composition to pharmacy. Phytother. Res. 2020, 34, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.P.; Khanum, F. Neuroprotective potential of phytochemicals. Pharmacogn. Rev. 2012, 6, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howes, M.J.; Simmonds, M.S. The role of phytochemicals as micronutrients in health and disease. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 558–566. [Google Scholar] [CrossRef]
- Adams, M.; Gmünder, F.; Hamburger, M. Plants traditionally used in age related brain disorders—A survey of ethnobotanical literature. J. Ethnopharmacol. 2007, 113, 363–381. [Google Scholar] [CrossRef]
- Nahin, R.L.; Barnes, P.M.; Stussman, B.J.; Bloom, B. Costs of Complementary and Alternative Medicine (CAM) and Frequency of Visits to CAM Practitioners, United States, 2007. Natl. Health Stat. Rep. 2009, 18, 1–14. [Google Scholar]
- Tella, A. The practice of traditional medicine in Africa. Niger. Med. J. J. Niger. Med. Assoc. 1979, 9, 607. [Google Scholar]
- Stafford, G.I.; Pedersen, M.E.; van Staden, J.; Jäger, A.K. Review on plants with CNS-effects used in traditional South African medicine against mental diseases. J. Ethnopharmacol. 2008, 119, 513–537. [Google Scholar] [CrossRef]
- Fennell, C.; Lindsey, K.; McGaw, L.; Sparg, S.; Stafford, G.; Elgorashi, E.; Grace, O.; Van Staden, J. Assessing African medicinal plants for efficacy and safety: Pharmacological screening and toxicology. J. Ethnopharmacol. 2004, 94, 205–217. [Google Scholar] [CrossRef]
- Nxumalo, N.; Alaba, O.; Harris, B.; Chersich, M.; Goudge, J. Utilization of traditional healers in South Africa and costs to patients: Findings from a national household survey. J. Public Health Policy 2011, 32, S124–S136. [Google Scholar] [CrossRef] [PubMed]
- de Montellano, B.O. Aztec Medicine, Health, and Nutrition; Rutgers University Press: New Brunswick, NJ, USA; Newark, NJ, USA ; Camden, NJ, USA, 1990. [Google Scholar]
- Kavasch, E.B.; Baar, K. American Indian Healing Arts: Herbs, Rituals, and Remedies for Every Season of Life; Bantam Books (USA): New York, NY, USA, 1999. [Google Scholar]
- Roeder, B.A. Chicano Folk Medicine from Los Angeles, California; University of California Press: Berkeley, CA, USA, 1985. [Google Scholar]
- But, P.-H.; Hu, S.-Y.; Kong, Y.C. Vascular plants used in Chinese medicine. Fitoterapia 1980, 51, 245–264. [Google Scholar]
- Huang, K.C. The Pharmacology of Chinese Herbs; Routledge: Abingdon, UK, 1998. [Google Scholar]
- Chiu, H.F.K.; Zhang, M. Dementia research in China. Int. J. Geriatr. Psychiatry 2000, 15, 947–953. [Google Scholar] [CrossRef]
- Kapoor, L. Handbook of Ayurvedic Medicinal Plants: Herbal Reference Library; CRC press: Boca Raton, FL, USA, 2000; Volume 2. [Google Scholar]
- Manyam, B.V. Dementia in ayurveda. J. Altern. Complement. Med. 1999, 5, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.M. Major Herbs of Ayurveda; Churchill Livingstone: London, UK, 2002. [Google Scholar]
- Angiolella, L.; Sacchetti, G.; Efferth, T. Antimicrobial and antioxidant activities of natural compounds. Evid. Based Complement. Altern. Med. 2018, 2018, 1945179. [Google Scholar] [CrossRef]
- Salehi, B.; Sharopov, F.; Martorell, M.; Rajkovic, J.; Ademiluyi, A.O.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Iriti, M.; Sharifi-Rad, J. Phytochemicals in Helicobacter pylori infections: What are we doing now? Int. J. Mol. Sci. 2018, 19, 2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Ozleyen, A.; Tumer, T.B.; Adetunji, C.O.; Omari, N.E.; Balahbib, A.; Taheri, Y.; Bouyahya, A.; Martorell, M.; Martins, N.; et al. Natural products and synthetic analogs as a source of antitumor drugs. Biomolecules 2019, 9, 679. [Google Scholar] [CrossRef]
- Rios, J.L.; Francini, F.; Schinella, G.R. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [Green Version]
- Gautam, R.; Jachak, S.M. Recent developments in anti-inflammatory natural products. Med. Res. Rev. 2009, 29, 767–820. [Google Scholar] [CrossRef]
- Velmurugan, B.K.; Rathinasamy, B.; Lohanathan, B.P.; Thiyagarajan, V.; Weng, C.F. Neuroprotective role of phytochemicals. Molecules 2018, 23, 2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, G.G. Current concepts of neurodegenerative diseases. Eur. Med. J. Neurol. 2014, 2, 78–86. [Google Scholar]
- Magalingam, K.B.; Radhakrishnan, A.; Ping, N.S.; Haleagrahara, N. Current concepts of neurodegenerative mechanisms in Alzheimer’s disease. Biomed. Res. Int. 2018, 2018, 3740461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, E.M.; De Miranda, B.; Sanders, L.H. Alpha-synuclein: Pathology, mitochondrial dysfunction and neuroinflammation in Parkinson’s disease. Neurobiol. Dis. 2018, 109, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Bhattacharya, R.; Mukherjee, A.; Pandey, D.K. Natural products against Alzheimer’s disease: Pharmaco-therapeutics and biotechnological interventions. Biotechnol. Adv. 2017, 35, 178–216. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. Baicalein as a potent neuroprotective agent: A review. Biomed. Pharmacother. 2017, 95, 1021–1032. [Google Scholar] [CrossRef]
- Carmona, V.; Martin-Aragon, S.; Goldberg, J.; Schubert, D.; Bermejo-Bescos, P. Several targets involved in Alzheimer’s disease amyloidogenesis are affected by morin and isoquercitrin. Nutr. Neurosci. 2018, 1–16. [Google Scholar] [CrossRef]
- Chakraborty, S.; Basu, S. Multi-functional activities of citrus flavonoid narirutin in Alzheimer’s disease therapeutics: An integrated screening approach and in vitro validation. Int. J. Biol. Macromol. 2017, 103, 733–743. [Google Scholar] [CrossRef]
- Hussain, G.; Zhang, L.; Rasul, A.; Anwar, H.; Sohail, M.U.; Razzaq, A.; Aziz, N.; Shabbir, A.; Ali, M.; Sun, T. Role of plant-derived flavonoids and their mechanism in attenuation of Alzheimer’s and Parkinson’s diseases: An update of recent data. Mol. (BaselSwitz.) 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Kim, S.H.; Yang, W.M. Mechanisms of action of phytochemicals from medicinal herbs in the treatment of Alzheimer’s disease. Planta Med. 2014, 80, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H.; Manczak, M.; Yin, X.; Grady, M.C.; Mitchell, A.; Tonk, S.; Kuruva, C.S.; Bhatti, J.S.; Kandimalla, R.; Vijayan, M.; et al. Protective effects of Indian spice curcumin against amyloid-beta in Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2018, 61, 843–866. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bai, L.; He, J.; Zhong, L.; Duan, X.; Ouyang, L.; Zhu, Y.; Wang, T.; Zhang, Y.; Shi, J. Recent advances in discovery and development of natural products as source for anti-Parkinson’s disease lead compounds. Eur. J. Med. Chem. 2017, 141, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Calina, D.; Docea, A.O.; Koirala, N.; Aryal, S.; Lombardo, D.; Pasqua, L.; Taheri, Y.; Marina Salgado Castillo, C.; Martorell, M. Curcumin’s Nanomedicine Formulations for Therapeutic Application in Neurological Diseases. J. Clin. Med. 2020, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wightman, E.L. Potential benefits of phytochemicals against Alzheimer’s disease. Proc. Nutr. Soc. 2017, 76, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabogal-Guaqueta, A.M.; Osorio, E.; Cardona-Gomez, G.P. Linalool reverses neuropathological and behavioral impairments in old triple transgenic Alzheimer’s mice. Neuropharmacology 2016, 102, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Bandyopadhyay, J.; Chakraborty, S.; Basu, S. Multi-target screening mines hesperidin as a multi-potent inhibitor: Implication in Alzheimer’s disease therapeutics. Eur. J. Med. Chem. 2016, 121, 810–822. [Google Scholar] [CrossRef]
- Finley, J.W.; Gao, S. A perspective on Crocus sativus L. (Saffron) constituent crocin: A potent water-soluble antioxidant and potential therapy for Alzheimer’s disease. J. Agric. Food Chem. 2017, 65, 1005–1020. [Google Scholar] [CrossRef]
- de Andrade Teles, R.B.; Diniz, T.C.; Costa Pinto, T.C.; de Oliveira Junior, R.G.; Gama, E.S.M.; de Lavor, E.M.; Fernandes, A.W.C.; de Oliveira, A.P.; de Almeida Ribeiro, F.P.R.; da Silva, A.A.M.; et al. Flavonoids as therapeutic agents in Alzheimer’s and Parkinson’s diseases: A systematic review of preclinical evidences. Oxid. Med. Cell Longev. 2018, 2018, 7043213. [Google Scholar] [CrossRef]
- Derf, A.; Sharma, A.; Bharate, S.B.; Chaudhuri, B. Aegeline, a natural product from the plant Aegle marmelos, mimics the yeast SNARE protein Sec22p in suppressing alpha-synuclein and Bax toxicity in yeast. Bioorg. Med. Chem. Lett. 2018, 29, 454–460. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Padureanu, R.; Albu, C.V.; Mititelu, R.R.; Bacanoiu, M.V.; Docea, A.O.; Calina, D.; Padureanu, V.; Olaru, G.; Sandu, R.E.; Malin, R.D. Oxidative stress and inflammation interdependence in multiple sclerosis. J. Clin. Med. 2019, 8, 1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, S. Oxidative Stress in Brain Ischemia. Brain Pathol. 1999, 9, 119–131. [Google Scholar] [CrossRef]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and Parkinson’s disease. Front. Neuroanat. 2015, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Hsu, L.J.; Sagara, Y.; Arroyo, A.; Rockenstein, E.; Sisk, A.; Mallory, M.; Wong, J.; Takenouchi, T.; Hashimoto, M.; Masliah, E. α-Synuclein promotes mitochondrial deficit and oxidative stress. Am. J. Pathol. 2000, 157, 401–410. [Google Scholar] [CrossRef]
- Jenner, P. Oxidative stress in Parkinson’s disease. Ann. Neurol. 2003, 53, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Lopez-Jornet, P.; Pons-Fuster López, E.; Calina, D.; Sharifi-Rad, M.; Ramírez-Alarcón, K.; Forman, K.; Fernández, M.; Martorell, M.; Setzer, W.N. Plant-Derived Bioactives in Oral Mucosal Lesions: A Key Emphasis to Curcumin, Lycopene, Chamomile, Aloe vera, Green Tea and Coffee Properties. Biomolecules 2019, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Martorell, M.; Arbiser, J.L.; Sureda, A.; Martins, N.; Maurya, P.K.; Sharifi-Rad, M.; Kumar, P.; Sharifi-Rad, J. Antioxidants: Positive or Negative Actors? Biomolecules 2018, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Rad, M.; Roberts, T.H.; Matthews, K.R.; Bezerra, C.F.; Morais-Braga, M.F.B.; Coutinho, H.D.M.; Sharopov, F.; Salehi, B.; Yousaf, Z.; Sharifi-Rad, M.; et al. Ethnobotany of the genus Taraxacum—Phytochemicals and antimicrobial activity. Phytother. Res. 2018, 32, 2131–2145. [Google Scholar] [CrossRef]
- Zang, C.X.; Bao, X.Q.; Li, L.; Yang, H.Y.; Wang, L.; Yu, Y.; Wang, X.L.; Yao, X.S.; Zhang, D. The protective effects of Gardenia jasminoides (Fructus Gardenia) on amyloid-beta-Induced mouse cognitive impairment and neurotoxicity. Am. J. Chin. Med. 2018, 46, 389–405. [Google Scholar] [CrossRef]
- Kim, S.M.; Park, Y.J.; Shin, M.S.; Kim, H.R.; Kim, M.J.; Lee, S.H.; Yun, S.P.; Kwon, S.H. Acacetin inhibits neuronal cell death induced by 6-hydroxydopamine in cellular Parkinson’s disease model. Bioorganic. Med. Chem. Lett. 2017, 27, 5207–5212. [Google Scholar] [CrossRef]
- Nataraj, J.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M. Neuroprotective effect of asiatic acid on rotenone-induced mitochondrial dysfunction and oxidative stress-mediated apoptosis in differentiated SH-SYS5Y cells. Nutr. Neurosci. 2017, 20, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Shal, B.; Ding, W.; Ali, H.; Kim, Y.S.; Khan, S. Anti-neuroinflammatory potential of natural products in attenuation of Alzheimer’s disease. Front. Pharm. 2018, 9, 548. [Google Scholar] [CrossRef] [PubMed]
- Sivanantham, B.; Krishnan, U.; Rajendiran, V. Amelioration of oxidative stress in differentiated neuronal cells by rutin regulated by a concentration switch. Biomed. Pharm. 2018, 108, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Sureda, A.; Habtemariam, S.; Nabavi, S.M. Ginsenoside Rd and ischemic stroke: A short review of literatures. J. Ginseng Res. 2015, 39, 299–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.W.; Jhang, K.A.; Woo, S.Y.; Kang, J.L.; Chong, Y.H. Sulforaphane exerts its anti-inflammatory effect against amyloid-beta peptide via STAT-1 dephosphorylation and activation of Nrf2/HO-1 cascade in human THP-1 macrophages. Neurobiol. Aging 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Bertram, E. The relevance of kindling for human epilepsy. Epilepsia 2007, 48, 65–74. [Google Scholar] [CrossRef]
- Kaur, H.; Bal, A.; Sandhir, R. Curcumin supplementation improves mitochondrial and behavioral deficits in experimental model of chronic epilepsy. Pharm. Biochem. Behav. 2014, 125, 55–64. [Google Scholar] [CrossRef]
- Tambe, R.; Patil, A.; Jain, P.; Sancheti, J.; Somani, G.; Sathaye, S. Assessment of luteolin isolated from Eclipta alba leaves in animal models of epilepsy. Pharm. Biol. 2017, 55, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Van Kampen, J.M.; Baranowski, D.B.; Shaw, C.A.; Kay, D.G. Panax ginseng is neuroprotective in a novel progressive model of Parkinson’s disease. Exp. Gerontol. 2014, 50, 95–105. [Google Scholar] [CrossRef]
- Zipp, F.; Aktas, O. The brain as a target of inflammation: Common pathways link inflammatory and neurodegenerative diseases. Trends Neurosci. 2006, 29, 518–527. [Google Scholar] [CrossRef]
- Salehi, B.; Shivaprasad Shetty, M.; V Anil Kumar, N.; Živković, J.; Calina, D.; Oana Docea, A.; Emamzadeh-Yazdi, S.; Sibel Kılıç, C.; Goloshvili, T.; Nicola, S. Veronica Plants—Drifting from Farm to Traditional Healing, Food Application, and Phytopharmacology. Molecules 2019, 24, 2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicha, P.; Tocharus, J.; Janyou, A.; Jittiwat, J.; Changtam, C.; Suksamrarn, A.; Tocharus, C. Hexahydrocurcumin protects against cerebral ischemia/reperfusion injury, attenuates inflammation, and improves antioxidant defenses in a rat stroke model. PLoS ONE 2017, 12, e0189211. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, J.; Shen, J. Herbal medicines for ischemic stroke: Combating inflammation as therapeutic targets. J. Neuroimmune Pharmacol. 2014, 9, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, Y.; Chen, Z.; Leng, S.X. Connection between systemic inflammation and neuroinflammation underlies neuroprotective mechanism of several phytochemicals in neurodegenerative diseases. Oxid. Med. Cell. Longev. 2018, 2018, 1972714. [Google Scholar] [CrossRef] [Green Version]
- Ianoşi, S.; Ianoşi, G.; Neagoe, D.; Ionescu, O.; Zlatian, O.; Docea, A.O.; Badiu, C.; Sifaki, M.; Tsoukalas, D.; Tsatsakis, A.M. Age-dependent endocrine disorders involved in the pathogenesis of refractory acne in women. Mol. Med. Rep. 2016, 14, 5501–5506. [Google Scholar] [CrossRef] [Green Version]
- Petrakis, D.; Vassilopoulou, L.; Mamoulakis, C.; Psycharakis, C.; Anifantaki, A.; Sifakis, S.; Docea, A.O.; Tsiaoussis, J.; Makrigiannakis, A.; Tsatsakis, A.M. Endocrine disruptors leading to obesity and related diseases. Int. J. Environ. Res. Public Health 2017, 14, 1282. [Google Scholar] [CrossRef]
- Liu, Z.; Ran, Y.; Huang, S.; Wen, S.; Zhang, W.; Liu, X.; Ji, Z.; Geng, X.; Ji, X.; Du, H.; et al. Curcumin protects against ischemic stroke by titrating microglia/macrophage polarization. Front. Aging Neurosci. 2017, 9, 233. [Google Scholar] [CrossRef]
- Akinmoladun, A.C.; Akinrinola, B.L.; Olaleye, M.T.; Farombi, E.O. Kolaviron, a Garcinia kola biflavonoid complex, protects against ischemia/reperfusion injury: Pertinent mechanistic insights from biochemical and physical evaluations in rat brain. Neurochem. Res. 2015, 40, 777–787. [Google Scholar] [CrossRef]
- Devinsky, O.; Vezzani, A.; O’Brien, T.J.; Jette, N.; Scheffer, I.E.; de Curtis, M.; Perucca, P. Epilepsy. Nat. Rev. Dis. Primers 2018, 4, 18024. [Google Scholar] [CrossRef]
- Xu, J.; Lacoske, M.H.; Theodorakis, E.A. Neurotrophic natural products: Chemistry and biology. Angew. Chem. (Int. Ed. Engl.) 2014, 53, 956–987. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Gwak, J.; Park, S.; Yang, C.S. Green tea polyphenol EGCG suppresses W nt/β-catenin signaling by promoting GSK-3β-and PP2A-independent β-catenin phosphorylation/degradation. Biofactors 2014, 40, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.; Ramalho, M.J.; Pereira, M.d.C.; Loureiro, J.A. Resveratrol brain delivery for neurological disorders prevention and treatment. Front. Pharmacol. 2018, 9, 1261. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.L.; Silva, V.D.; Dos Santos Souza, C.; Santos, C.C.; Paris, I.; Munoz, P.; Segura-Aguilar, J. Impact of plant-derived flavonoids on neurodegenerative diseases. Neurotox. Res. 2016, 30, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, R.; Zhan, Z.; Li, X.; Zhou, F.; Xing, A.; Jiang, C.; Chen, Y.; An, L. Beneficial effects of sulforaphane treatment in Alzheimer’s disease may be mediated through reduced HDAC1/3 and increased P75NTR expression. Front. Aging Neurosci. 2017, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, T.C.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.A. Naturally occurring acetylcholinesterase inhibitors and their potential use for Alzheimer’s disease therapy. Front. Pharmacol. 2018, 9, 1192. [Google Scholar] [CrossRef] [Green Version]
- Francis, P.T.; Palmer, A.M.; Snape, M.; Wilcock, G.K. The cholinergic hypothesis of Alzheimer’s disease: A review of progress. J. Neurol. Neurosurg. Psychiatry 1999, 66, 137–147. [Google Scholar] [CrossRef]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the cholinergic system. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Durães, F.; Pinto, M.; Sousa, E. Old drugs as new treatments for neurodegenerative diseases. Pharmaceuticals 2018, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Finberg, J.P.; Rabey, J.M. Inhibitors of MAO-A and MAO-B in psychiatry and neurology. Front. Pharmacol. 2016, 7, 340. [Google Scholar] [CrossRef] [Green Version]
- Larit, F.; Elokely, K.M.; Chaurasiya, N.D.; Benyahia, S.; Nael, M.A.; Leon, F.; Abu-Darwish, M.S.; Efferth, T.; Wang, Y.H.; Belouahem-Abed, D.; et al. Inhibition of human monoamine oxidase A and B by flavonoids isolated from two Algerian medicinal plants. Phytomed. Int. J. Phytother. Phytopharm. 2018, 40, 27–36. [Google Scholar] [CrossRef]
- Carradori, S.; Petzer, J.P. Novel monoamine oxidase inhibitors: A patent review (2012–2014). Expert Opin. Ther. Pat. 2015, 25, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Zanforlin, E.; Zagotto, G.; Ribaudo, G. The medicinal chemistry of natural and semisynthetic compounds against Parkinson’s and Huntington’s diseases. ACS Chem. Neurosci. 2017, 8, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wang, S.; Ma, F.; Zhang, Y.; Peng, Y.; Xing, C.; Feng, Y.; Wang, X.; Peng, Y. From stroke to neurodegenerative diseases: The multi-target neuroprotective effects of 3-n-butylphthalide and its derivatives. Pharmacol. Res. 2018, 135, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowski, V.V.; Calina, D.; Tsarouhas, K.; Pivnik, A.V.; Sergievich, A.A.; Kodintsev, V.V.; Filatova, E.A.; Ozcagli, E.; Docea, A.O.; Arsene, A.L. A guide to acquired vitamin K coagulophathy diagnosis and treatment: The Russian perspective. Daru J. Pharm. Sci. 2017, 25, 10. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.H.; Yen, T.L.; Hsu, C.Y.; Thomas, P.A.; Sheu, J.R.; Jayakumar, T. Multi-targeting andrographolide, a novel NF-kappaB inhibitor, as a potential therapeutic agent for stroke. Int. J. Mol. Sci. 2017, 18, 1638. [Google Scholar] [CrossRef]
- Pirzada, A.M.; Ali, H.H.; Naeem, M.; Latif, M.; Bukhari, A.H.; Tanveer, A. Cyperus rotundus L.: Traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2015, 174, 540–560. [Google Scholar] [CrossRef]
- Handin, R.I. The history of antithrombotic therapy: The discovery of heparin, the vitamin K antagonists, and the utility of aspirin. Hematol. Oncol. Clin. 2016, 30, 987–993. [Google Scholar] [CrossRef]
- Copeland, C.E.; Six, C.K. A tale of two anticoagulants: Warfarin and heparin. J. Surg. Educ. 2009, 66, 176–181. [Google Scholar] [CrossRef]
- Chen, C.; Yang, F.Q.; Zhang, Q.; Wang, F.Q.; Hu, Y.J.; Xia, Z.N. Natural Products for Antithrombosis. Evid. Based Complement. Altern. Med. 2015, 2015, 876426. [Google Scholar] [CrossRef] [Green Version]
- Santos, G.; Giraldez-Alvarez, L.D.; Avila-Rodriguez, M.; Capani, F.; Galembeck, E.; Neto, A.G.; Barreto, G.E.; Andrade, B. SUR1 receptor interaction with hesperidin and linarin predicts possible mechanisms of action of Valeriana officinalis in Parkinson. Front. Aging Neurosci. 2016, 8, 97. [Google Scholar] [CrossRef]
- Nataraj, J.; Manivasagam, T.; Justin Thenmozhi, A.; Essa, M.M. Neurotrophic effect of asiatic acid, a triterpene of Centella asiatica against chronic 1-methyl 4-phenyl 1, 2, 3, 6-tetrahydropyridine hydrochloride/probenecid mouse model of Parkinson’s disease: The role of MAPK, PI3K-Akt-GSK3beta and mTOR signalling pathways. Neurochem. Res. 2017, 42, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Ryu, H.W.; Kang, M.G.; Park, D.; Lee, H.; Shin, H.M.; Oh, S.R.; Kim, H. Potent inhibition of monoamine oxidase A by decursin from Angelica gigas Nakai and by wogonin from Scutellaria baicalensis Georgi. Int. J. Biol. Macromol. 2017, 97, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, D.; Kaur Dogra, A.; Tahrani, A.; Herrmann, F.; Wink, M. Extracts from traditional chinese medicinal plants inhibit acetylcholinesterase, a known Alzheimer’s disease target. Molecules (Basel Switzerland ) 2016, 21, 1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoncello, K.T.; Aguiar, G.P.S.; Oliveira, J.V.; Siebel, A.M. Micronization potentiates curcumin’s anti-seizure effect and brings an important advance in epilepsy treatment. Sci. Rep. 2018, 8, 2645. [Google Scholar] [CrossRef]
- Seo, E.J.; Fischer, N.; Efferth, T. Phytochemicals as inhibitors of NF-kappaB for treatment of Alzheimer’s disease. Pharmacol. Res. 2018, 129, 262–273. [Google Scholar] [CrossRef]
- El Alaoui, C.; Chemin, J.; Fechtali, T.; Lory, P. Modulation of T-type Ca2+ channels by Lavender and Rosemary extracts. PLoS ONE 2017, 12, e0186864. [Google Scholar] [CrossRef] [Green Version]
- Pohl, F.; Kong Thoo Lin, P. The potential use of plant natural products and plant extracts with antioxidant properties for the prevention/treatment of neurodegenerative diseases: In vitro, in vivo and clinical trials. Molecules (Basel Switzerland ) 2018, 23, 3283. [Google Scholar] [CrossRef] [Green Version]
- Elufioye, T.O.; Berida, T.I.; Habtemariam, S. Plants-derived neuroprotective agents: Cutting the cycle of cell death through multiple mechanisms. Evid. Based Complement. Altern. Med. 2017, 2017, 3574012. [Google Scholar] [CrossRef] [Green Version]
- Ngoungoure, V.L.; Schluesener, J.; Moundipa, P.F.; Schluesener, H. Natural polyphenols binding to amyloid: A broad class of compounds to treat different human amyloid diseases. Mol. Nutr. Food Res. 2015, 59, 8–20. [Google Scholar] [CrossRef]
- Wang, M.; Li, Y.J.; Ding, Y.; Zhang, H.N.; Sun, T.; Zhang, K.; Yang, L.; Guo, Y.Y.; Liu, S.B.; Zhao, M.G.; et al. Silibinin prevents autophagic cell death upon oxidative stress in cortical neurons and cerebral ischemia-reperfusion injury. Mol. Neurobiol. 2016, 53, 932–943. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, J.; Fang, L.; Li, X.; Zhao, Y.; Shi, W.; An, L. Neuroprotective effects of sulforaphane on cholinergic neurons in mice with Alzheimer’s disease-like lesions. Int. J. Mol. Sci. 2014, 15, 14396–14410. [Google Scholar] [CrossRef] [PubMed]
- Dar, N.J.; Hamid, A.; Ahmad, M. Pharmacologic overview of Withania somnifera, the Indian Ginseng. Cell. Mol. Life Sci. CMLS 2015, 72, 4445–4460. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Compound/Type | Natural Source | Experimental Model | Effects/Mechanisms of action | Ref |
|---|---|---|---|---|
| Acacetin/flavanoid | Chrysanthemi indici, Calamintha, Linaria spp | In vitro model of PD | ↓6-hydroxydopamine-induced cell death ↓caspase-3, ↓caspase-9, ↓PARP and cytochrome c ↑Bcl-2/Bax, ↓ROS, ↓phosphorylation of JNK, ↓p38, ↓ERK1/2 MAPK | [69] |
| Aegeline/alkaloid-amide | Aegle marmelos | In vitro yeast model of PD | Prevented α-synuclein-induced apoptosis, ↓ROS | [58] |
| Andrographolide/diterpene lactone | Andrographis paniculata | In vitro model of PD | ↓PAF-induced platelet aggregation, ↓collagen-stimulated platelet activation, ↑TXA2, ↑phosphorylation of PKC, MAPK and AKT ↑eNOS, ↑NO, ↑eNOS-NO/cyclic GMP pathways, ↓PI3K/Akt/p38 MAPK ↓ PLC-γ2/PKC | [104] |
| Apigenin/flavanoid | common constituent in plants | In vitro induced neurogenesis In vivo mouse model of AD | ↓inflammatory cytokines, ↓cortical hyperexcitation ↓Aβ burden, ↓oxidative stress, ↑ERK/CREB/BDNF pathway ↓β-amyloid neurotoxicity, ↑mitochondrion protection | [44,92] [109] |
| Asiatic acid/triterpene | Centella asiatica | In vitro model of PD In vivo mouse model of PD | ↓apoptosis, ↓ROS ↑ERK, ↑PI3K/Akt/mTOR/GSK-3β pathways, ↓MAPK/P38, ↓JNK, ↓ERK, ↓dopamine depletion, ↑NTFs | [110] [70] |
| Baicalein/flavanoid | Scutellaria baicalensis | Molecular docking simulation In vivo model of PD | ↓MAO-A, ↓Aβ ↓brain hypoxia, ↓H2O2, ↓iNOS, ↓NF-κB, ↓NO, ↓TNF-α, ↓oxidative stress, ↓mitochondrial dysfunction, ↑JNK, ↓TNF-α, ↓IL-6, ↓NF-κB, ↓MAPK, ↓dopaminergic neuron loss, ↓LDH, ↓NO, ↓glutamate | [111] [45] |
| Berberine/alkaloid | Berberis genus | In vitro model of AD In vivo rodent model of AD | ↓AChE, ↓MAO-B, ↓BACE1, ↑IκB-α, ↑Akt,↑p38 kinase ERK1/2 ↓NF-κB, ↓TNF-α,↓IL-6 production, ↓MCP-1, ↓COX 2,↓iNOS ↓Aβ plaque, ↓ CTF-α, ↓CTF-β (which reflects α- and β-secretase processing of APP) | [112] [101] [49] |
| Borneol/terpene derivative | common constituent in plants | Ex vivo rat blood | ↑PT, ↑TT, ↓thrombosis in veins | [108] |
| Carvacrol/monoterpenoid phenol | Cyperus rotundus | In vitro MAO A and MAO B | ↓antiplatelet aggregation | [105] |
| Chrysin/flavanoid | Hypericum afrum, Cytisus villosus | Molecular docking simulation | ↓MOA-A | [99] |
| Crocin/carotenoid | Gardenia jasminoides Crocus sativus | In vivo mouse model of AD In vivo rat model of AD | ↓oxidative stress, ↑SOD, ↓MDA ↓AChE, ↑ACh activity ↓neuroinflammation, ↓TNF-α, ↓PGE, ↓iNOS, ↓COX2 ↓Tau hyperphosphorylation | [68] [56] |
| Curcumin/carotenoid | Curcuma longa | In vivo Dania rerio (zebrafish) model of NDD In vivo mouse model of stroke in vivo mice model of PD | Neuroprotective,↓tonic-clonic seizures ↓oxidative stress ↑GSH in cortex and hippocampus ↓infarct volumes, ↑M2 polarization of microglia/macrophages, ↓Aβ aggregation, ↓NF-κB, ↓α-synuclein oligomerization | [113] [76] [86] [50,114] |
| Decursin/pyranocoumarin | Angelica gigas, Scutellaria baicalensis | In vitro model of PD | ↓MOA-A | [111] |
| Epigallocatechin gallate/catechin | Camellia sinensis | In vivo rat model of AD rat model of PD | ↓Aβ fibrillogenesis, ↓oxidative stress, ↓AchE ↓α-synuclein aggregation | [18,51] |
| Genistein/flavanoid | Glycine max | Molecular docking simulation In vitro model of AD | ↓MAO ↓inflammation, ↓NF-κB ↓Aβ toxicity, ↑apoptosis | [99] [44] |
| Ginsenoside Rd/triterpene glycosides | Panax ginseng | In vivo rodent model of stroke | ↓excitotoxicity, ↓Ca2+ influx, ↑ GLT-1, ↓ ROS | [73] |
| Ginsenoside Rg1/triterpene glycosides | Panax notoginseng | In vitro cell model of AD | ↓β- and γ-secretases, ↓NO, ↓ROS, ↓lipid peroxidation, ↓IL-1,↓IL-8, ↓TNF-α, ↓Aβ plaque, ↓caspase-9, ↓caspase-3 | [49] |
| Hesperidin/flavanoid | Valeriana officinalis | Molecular docking simulation In vivo rat model of AD | ↓ BACE1 ↓oxidative stress, ↓Aβ fibril formation | [109] [46,55] |
| Isoquercitrin/flavonoid | Common in plants | In vivo rat model of AD | ↓BACE1, ↓γ-secretase, ↓Aβ fibrillogenesis, ↓caspase-3, ↓caspase-9, ↓apoptosis, ↓amyloid plaque, ↓tau hyperphosphorylation | [46] |
| Kolaviron/bioflavanoid complex | Garcinia kola | In vivo rat model of stroke | ↓MPO, ↓necrotic cell death, Preserved Na/K/ATPase activity | [87] |
| Linalool/monoterpene | Lavandula spp. Rosmarinus officinalis Melissa officinalis Cymbopogon citratus | In vivo mouse model of AD In vitro cell model of ND | Anti-inflammatory ↓p38, ↓MAPK, ↓Nos2, ↓COX2, ↓IL-1β↓ Aβ in the hippocampus ↓tauopathy, inhibition of T-type Ca2+ channels | [54] [115] |
| Luteolin/flavanoid | Common constituent in plants | In vivo mouse model of ND In vivo animal model of stroke | ↑GSH, ↓oxidative stress, ↓MDA, ↑Nrf2, antioxidant/anti-inflammatory ↑Nrf-2 dependent transcription of HO-1 neuroprotective against cerebral I/R injury | [77] [116] [82] |
| Morin/flavanoid | Common constituent in plants | In vivo rat models of AD | ↓BACE1, ↓γ-secretase, ↓Aβ fibrillogenesis ↓apoptosis, ↑caspase-3, ↑caspase-9 ↓amyloid plaque, ↓tau hyperphosphorylation | [46] |
| Myricetin/flavanoid | Common constituent in plants | Molecular docking simulation | ↓MAO | [99] |
| Naringenin/flavanoid | Citrus paradise Citrus sinensis | In vitro models of AD | ↓inflammatory cytokines, ↓NF-κB signalling, ↑Nrf2/ARE signaling ↓NO | [71] |
| Naringin/flavanoid | Citrus spp. | In vivo rat model of AD In vivo rodent model of PD | ↓AChE, ↓cognitive deficit, ↓GFAP, ↑neurotrophic factors | [92] [48] |
| Narirutin/flavanoid | Citrus spp. | In vitro | ↓BACE1 ↓Aβ aggregation | [47] |
| Nootkatone/sesquiterpene | Cyperus rotundus | In vitro MAOA and MAOB | ↓platelet aggregation | [105] |
| Quercetin/flavanoid | Tea, citrus | Molecular docking simulation In vivo mouse model of AD In vivo rodent model of PD | ↓MAO, ↓PKC-ε ↓oxidative stress by ERK1/2 phosphorylation, p38MAPK dephosphorylation, ↓TNF-α, ↓IL-6, ↓GFAP, ↓MDA, ↑glutathione peroxidase, ↑AMPK activity ↓apoptosis, ↓GSK3β, ↓tau phosphorylation, ↓ROS, ↓Aβ aggregation ↓ BACE1, ↑NF-κB, ↓ROS, improved 6-OHDA-induced tremors | [99] [116] [48] [40] [57] |
| Resveratrol/stilbenoid | Vitis vinifera | In vivo rat model of PD In vivo rodent model of PD | ↓COX2, ↓TNF-α, ↓NF-κB, ↓β-amyloid plaques ↓TNF-α, ↓IL-6, ↑BDNF, ↑IL-10, ↓TNF-α, ↓NF-κB ↑ERK1-2/CREB, ↑BDNF, ↑GDNF, ↓NO, ↓iNOS, ↓Aβ in glial cells, ↑AMPL-SIRT-1 | [53] [40] |
| Rutin/flavanoid | Abundant in Citrus fruits | In vitro In vitro In vivo rodent model of AD | ↓pro-inflammatory cytokines, ↓ROS Protected neurons against oxidative injury ↑SOD, ↑CAT, ↑GPx, ↓iNOS ↑MAPK, ↑apoptosis, ↑JNK, ↑p38 MAPK ↓ IL-1, ↓IL-6, ↑BDNP expression | [72] [92] [117] [92] |
| Silibinin/flavanoid | Silybinisus laborinum | In vivo rat model of AD In vivo rat model of PD In vivo rat model of stroke | ↓AChE, ↓ROS ↓Aβ aggregation, ↓hypoxic/ischemic injury Protected neurons from H2O2-mediated oxidative stress ↓LC3-II, ↓Beclin-1 levels | [118] [57] [119] |
| Sulforaphane/isothiocynate | Cruciferous vegetables | In vitro cell model of AD In vivo mouse model of AD | ↓IL-1β, ↓Aβ1-42-stimulated THP-1 macrophages Dephosphorylated STAT-1, ↑Nrf2 ↑neurogenesis, ↓aluminium load, ↓Aβ deposition ↑p75NTR, ↓Aβ burden | [74] [120] [93] |
| Withanamides A and C/amido compounds | Withania somnifera | In vivo rat model of AD | ↓Aβ fibril formation | [44] |
| Withanolide A/amido compound | Withania somnifera | In vivo rat model of AD | ↑axonal/dendritic regeneration exhibited neurotrophic activity | [89] |
| Withanone | Withania somnifera | In vivo rat model of AD | Protect neurons and glial cells | [121] |
| Wogonin/flavanoid | Suctellaria baicalensis | In vivo rat model of stroke | ↓synthesis of thrombin, ↓factor-Xa ↓APTT, ↓PT | [108] |
| α-cyperone | Cyperus rotundus | In vivo rodent model of stroke | ↓platelet aggregation | [105] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharifi-Rad, M.; Lankatillake, C.; Dias, D.A.; Docea, A.O.; Mahomoodally, M.F.; Lobine, D.; Chazot, P.L.; Kurt, B.; Boyunegmez Tumer, T.; Catarina Moreira, A.; et al. Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics. J. Clin. Med. 2020, 9, 1061. https://doi.org/10.3390/jcm9041061
Sharifi-Rad M, Lankatillake C, Dias DA, Docea AO, Mahomoodally MF, Lobine D, Chazot PL, Kurt B, Boyunegmez Tumer T, Catarina Moreira A, et al. Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics. Journal of Clinical Medicine. 2020; 9(4):1061. https://doi.org/10.3390/jcm9041061
Chicago/Turabian StyleSharifi-Rad, Mehdi, Chintha Lankatillake, Daniel A. Dias, Anca Oana Docea, Mohamad Fawzi Mahomoodally, Devina Lobine, Paul L. Chazot, Begum Kurt, Tugba Boyunegmez Tumer, Ana Catarina Moreira, and et al. 2020. "Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics" Journal of Clinical Medicine 9, no. 4: 1061. https://doi.org/10.3390/jcm9041061
APA StyleSharifi-Rad, M., Lankatillake, C., Dias, D. A., Docea, A. O., Mahomoodally, M. F., Lobine, D., Chazot, P. L., Kurt, B., Boyunegmez Tumer, T., Catarina Moreira, A., Sharopov, F., Martorell, M., Martins, N., Cho, W. C., Calina, D., & Sharifi-Rad, J. (2020). Impact of Natural Compounds on Neurodegenerative Disorders: From Preclinical to Pharmacotherapeutics. Journal of Clinical Medicine, 9(4), 1061. https://doi.org/10.3390/jcm9041061