
Effect of Knotweed in Diet on Physiological Changes in Pig

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Fodders and Feeding Regime
- Selvico (prestarter) is a complete feed designed for piglets up to 8 kg. It is served 7–14 days before weaning to secure the habit and then 10–14 days after weaning. It contains barley, wheat, genetically modified (GM) soybean meal, dried rhizome of Bohemian knotweed, lactose, secondary products from sweets manufacturing, soy oil, sugar beet pulp, fish meal, soy protein, glucose and dried poultry blood.
- ČOS = EW (Early Weaning) is a complete feed designed for the nutrition of piglets up to 18 kg. It contains barley, wheat, soybean meal GM, dried poultry blood, bran, fish meal, dried rhizome of Bohemian knotweed, lactose, fish oil, calcium dihydrogen phosphate and calcium carbonate.
- A1 (subsequent mixture) is a complete feed intended for the fattening of pigs up to 35 kg. It contains barley, wheat, soybean meal GM, bran, sugar beet pulp, dried rhizome of Bohemian knotweed, soy oil, calcium carbonate, sodium chloride, calcium dihydrogen phosphate and dried poultry blood.
- KPB is a commonly used mixture for pregnant sows, but in our case, we used it to feed pigs from 35 kg to the carcass weight. It contains barley, wheat, soybean meal GM, dried poultry blood, bran, fish meal, dried rhizome of Bohemian knotweed, fish oil, lactose, calcium dihydrogen phosphate and calcium carbonate.
2.3. Measured Characteristics
2.3.1. Feed Consumption
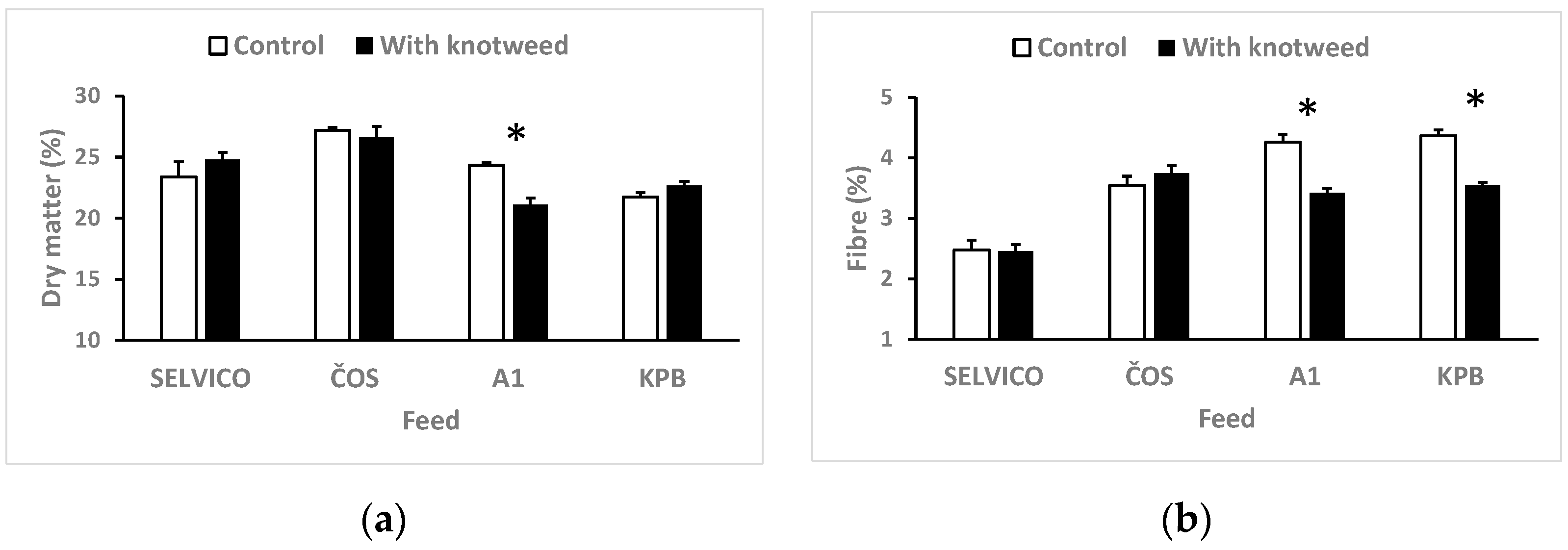
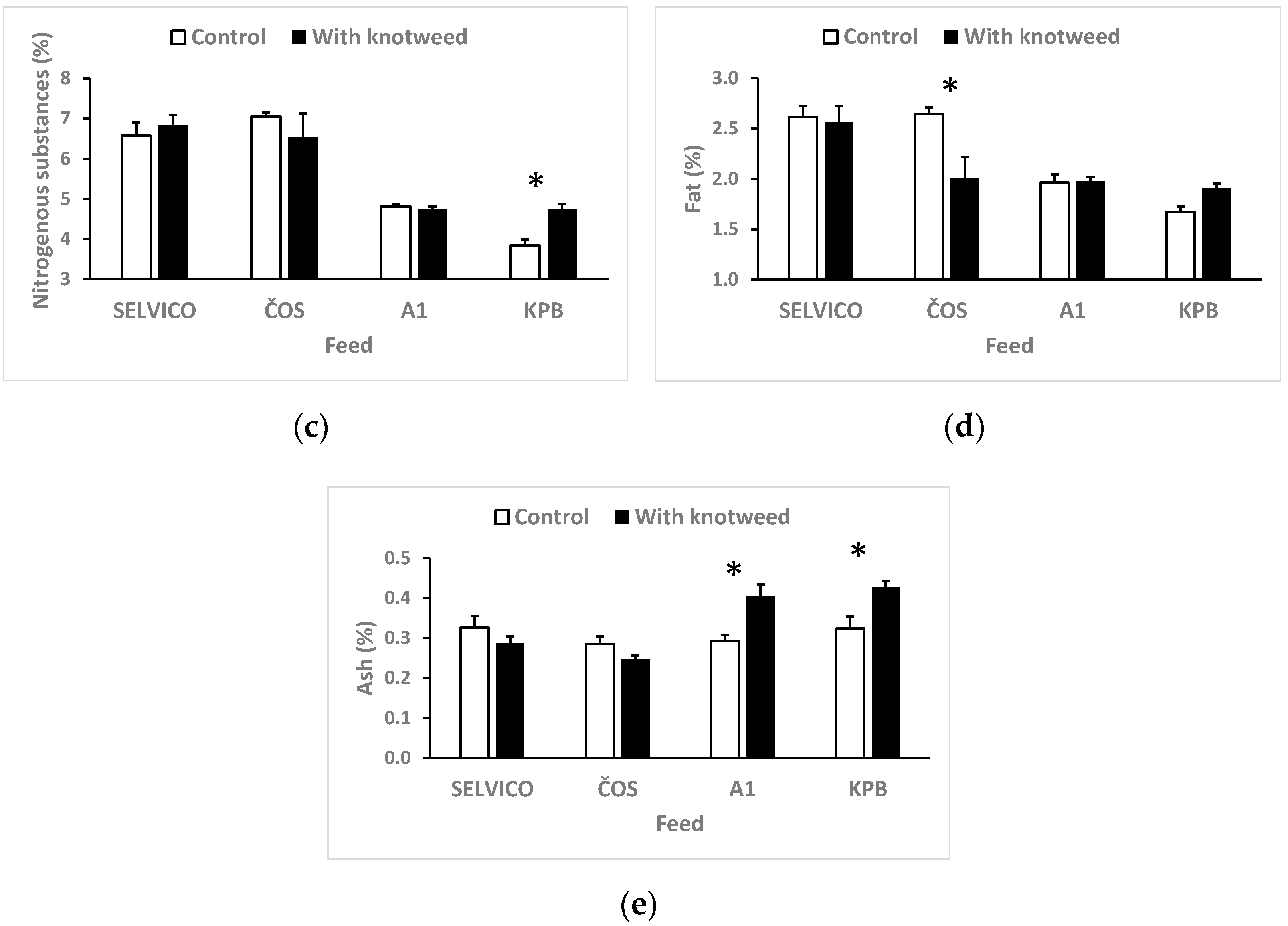
2.3.2. Composition of Excrements
2.3.3. Weight of Piglets
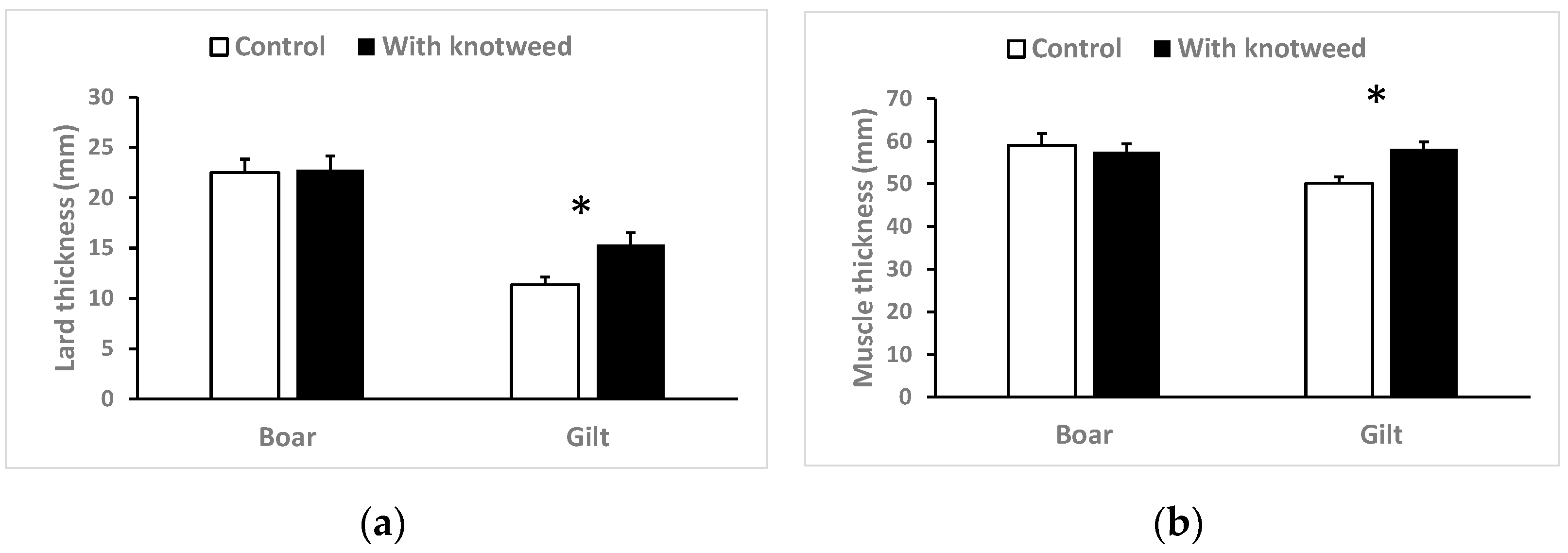
2.3.4. Muscle-To-Fat Ratio
2.3.5. Haematology and Biochemistry
2.3.6. Statistical Data Evaluation
3. Results
3.1. Fodder Consumption and Digestibility
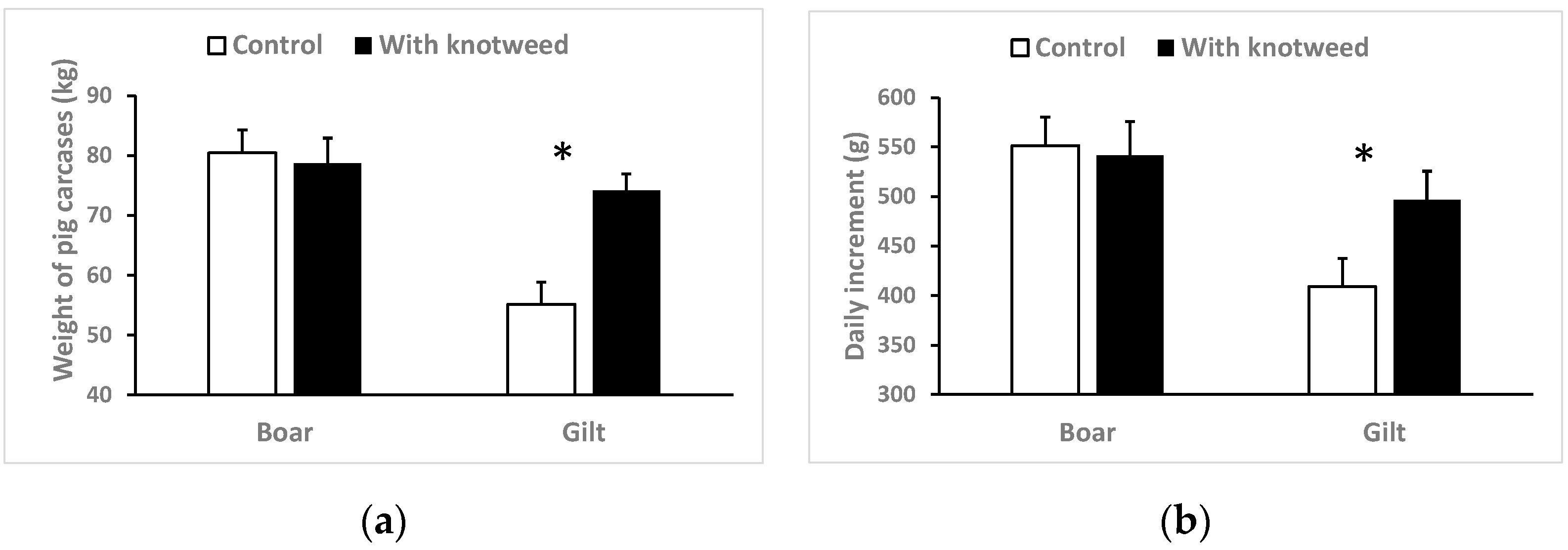
3.2. Total Weight and Increments
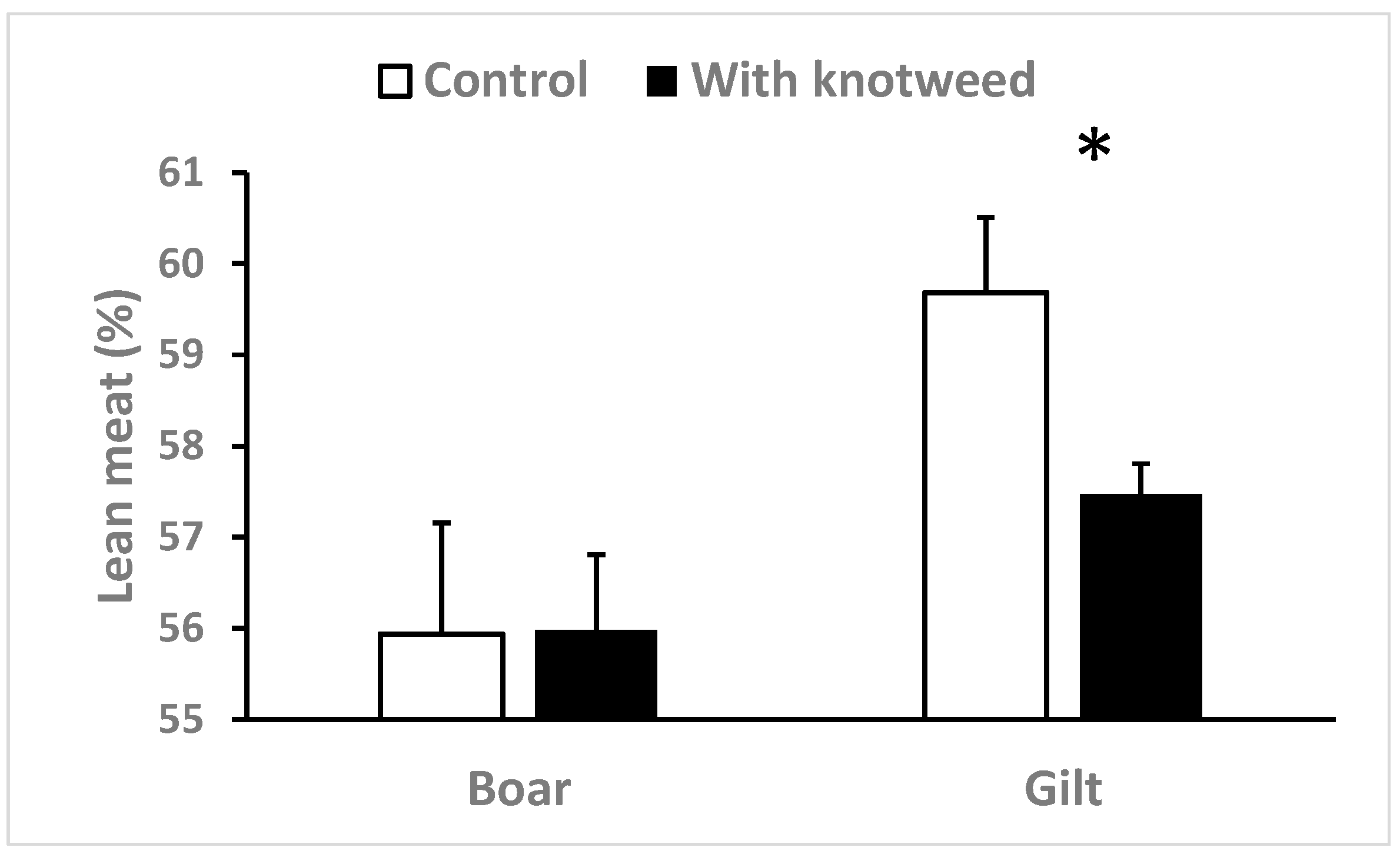
3.3. The Muscle-To-Fat Ratio and Fatty Acid Content
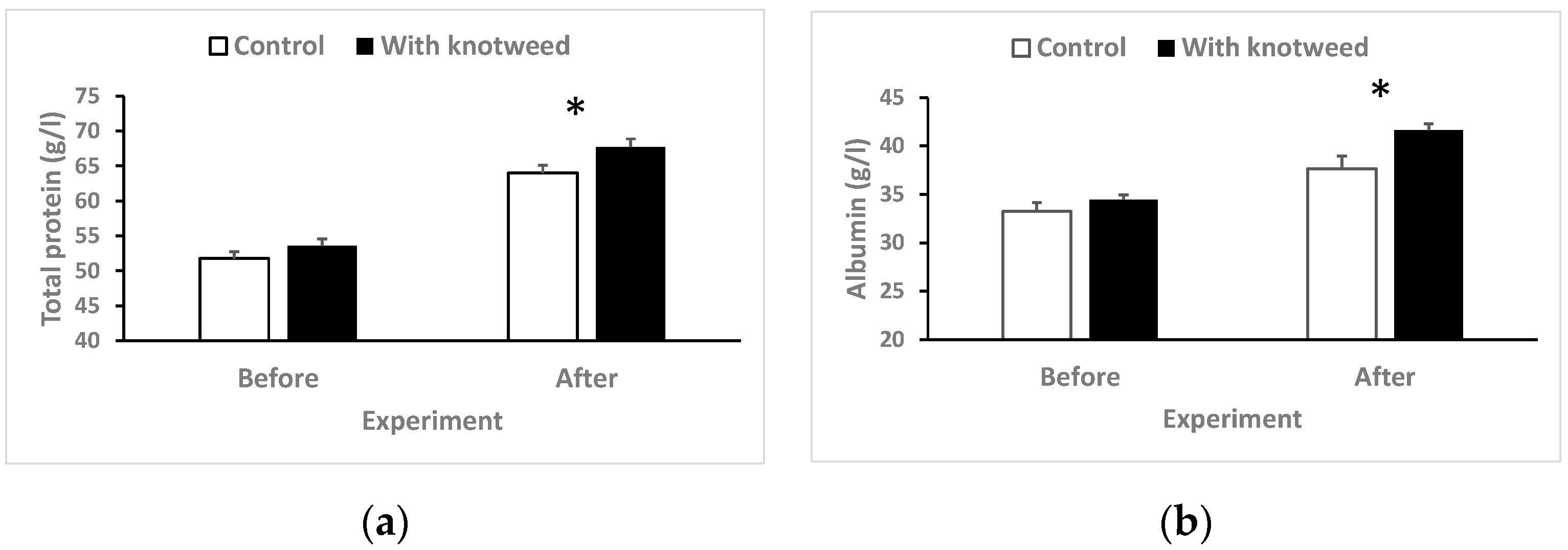
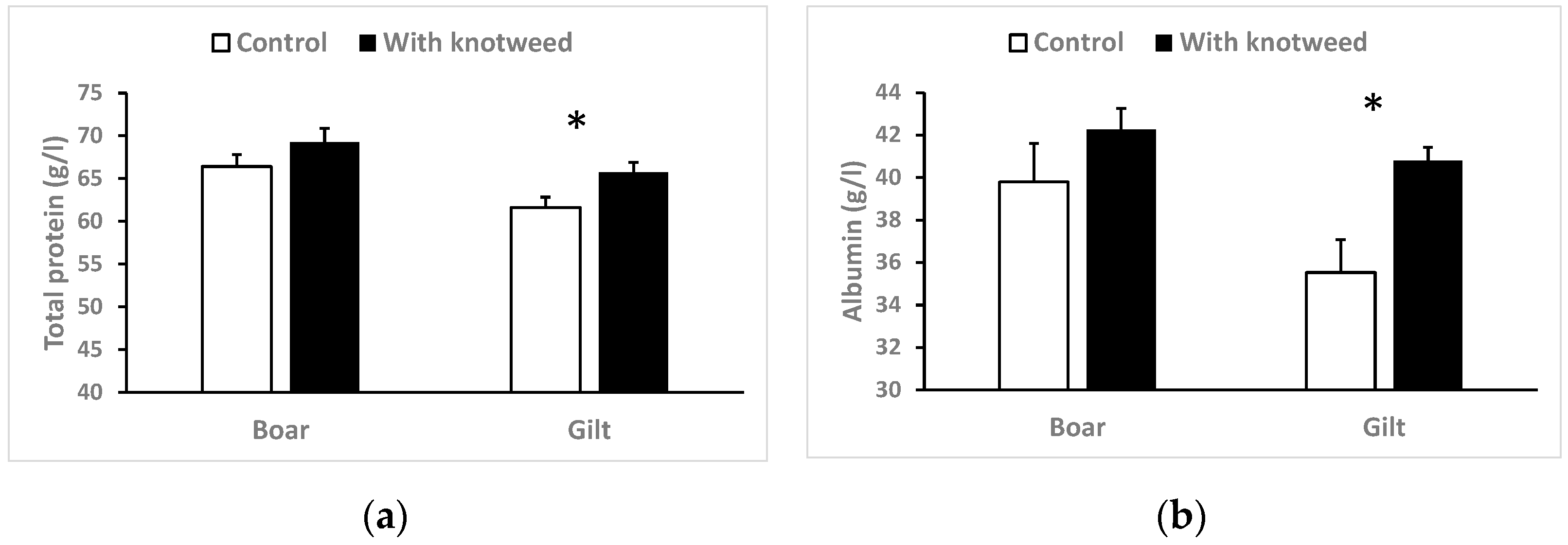
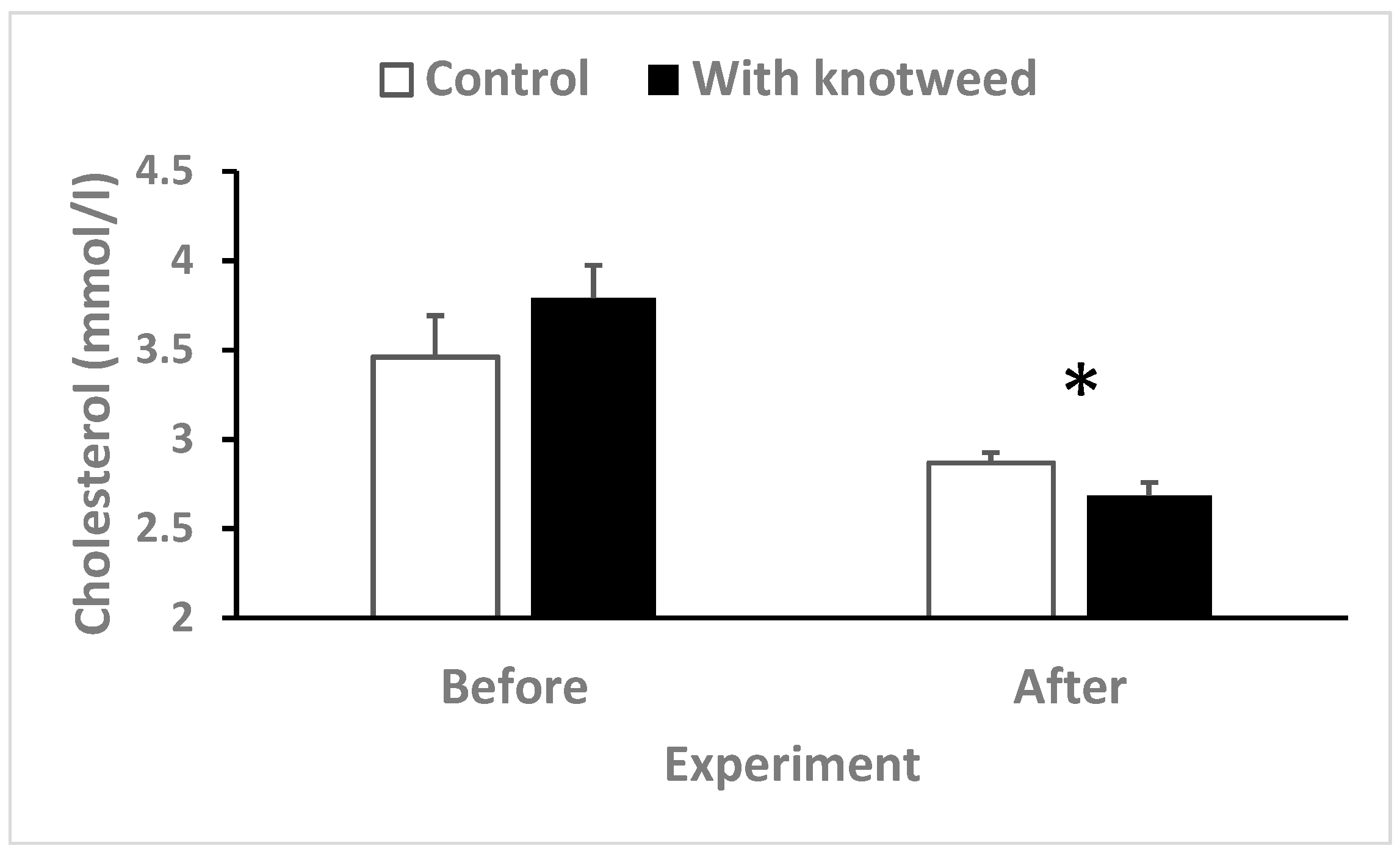
3.4. Biochemistry and Haematology
3.4.1. Total Serum Protein
3.4.2. Serum Cholesterol
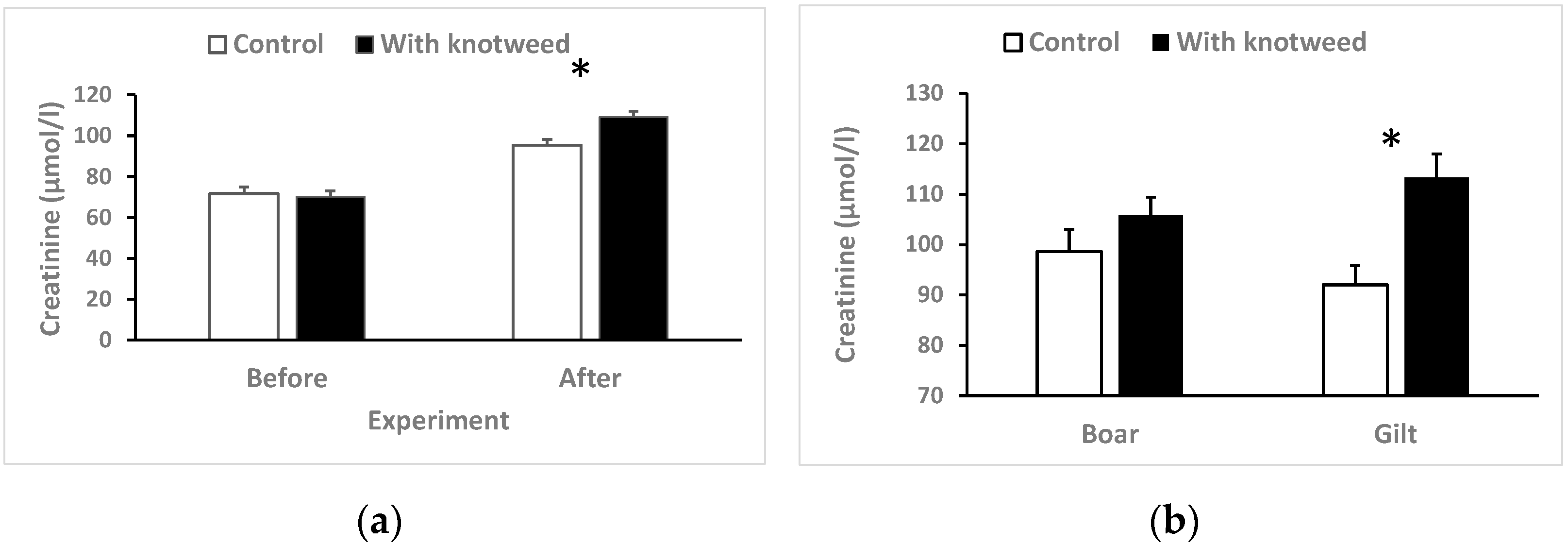
3.4.3. Serum Creatinine
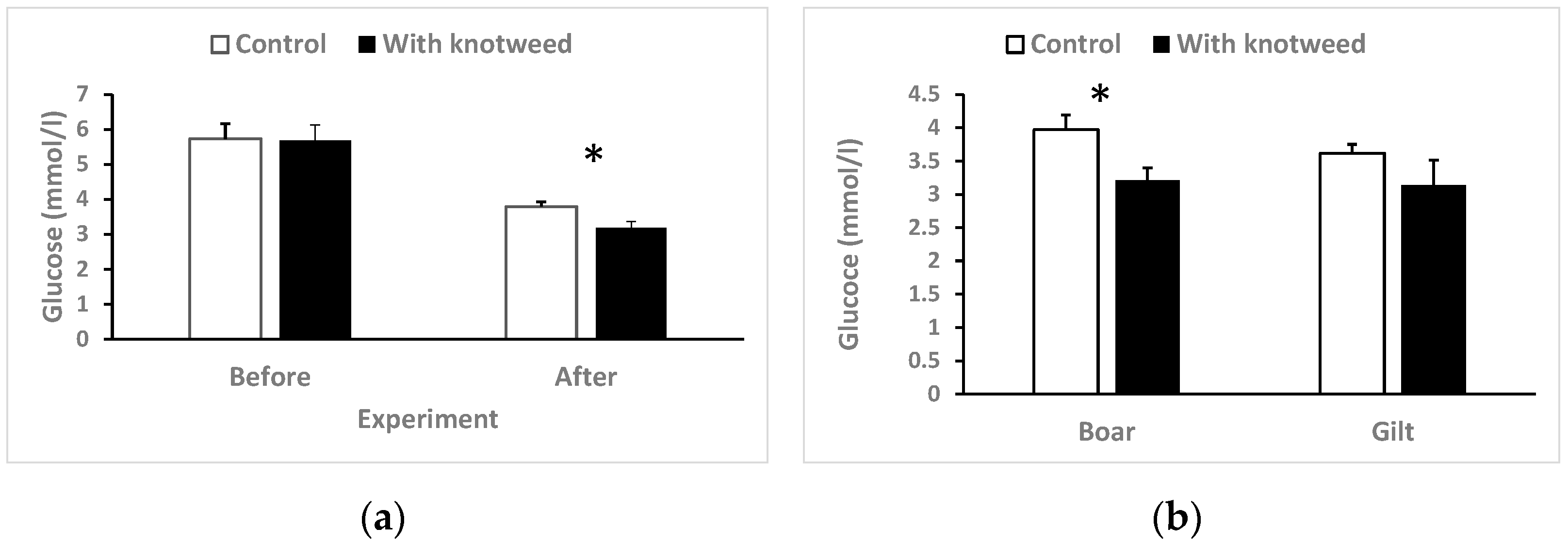
3.4.4. Serum Glucose
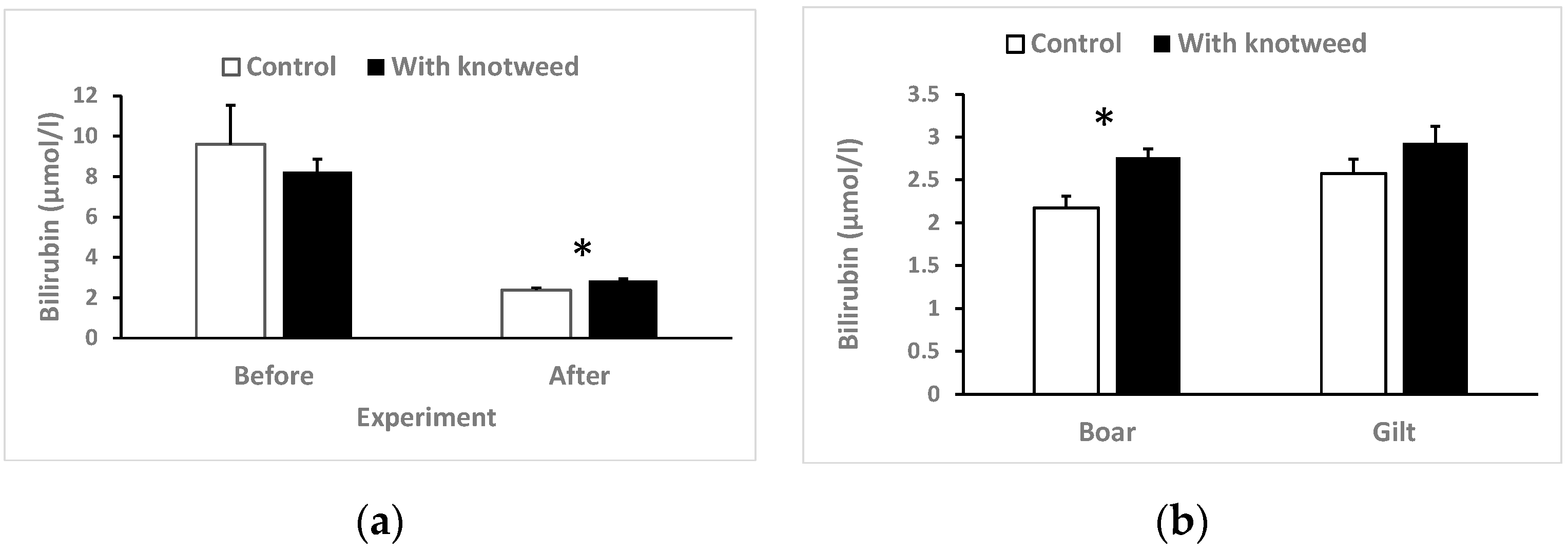
3.4.5. Total Serum Bilirubin
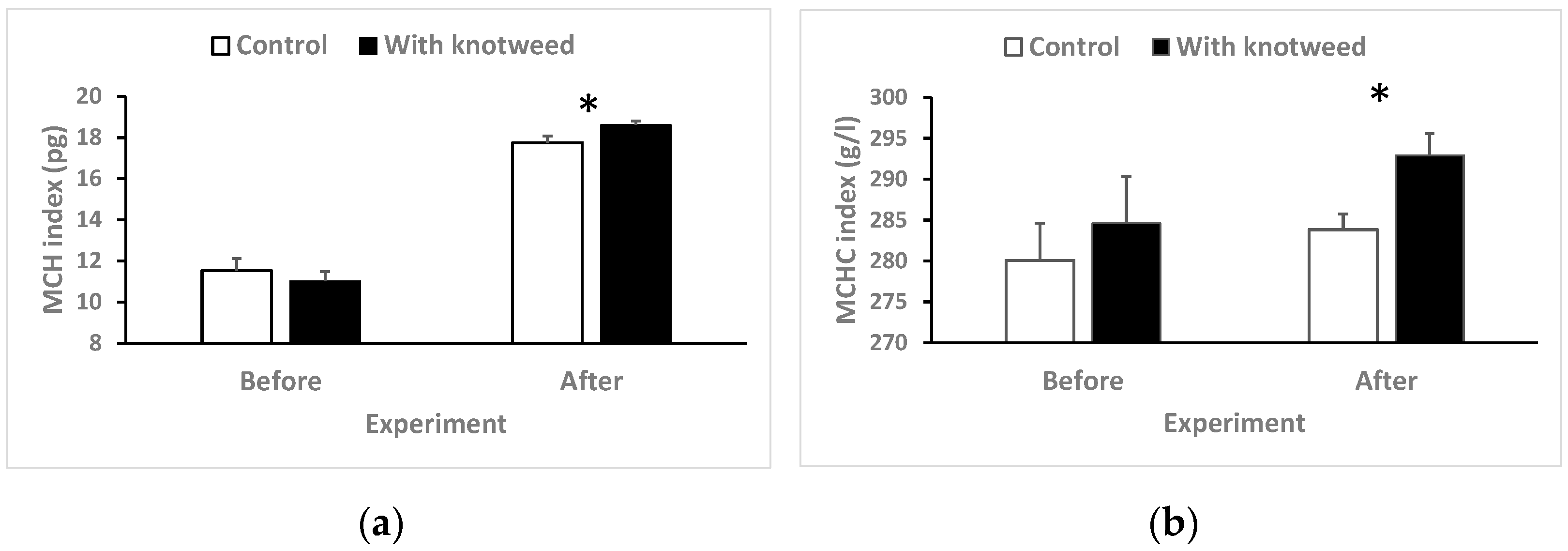
3.4.6. Mean Cell Hemoglobin (MCH) and Mean Corpuscular Hemoglobin Concentration (MCHC) Indexes
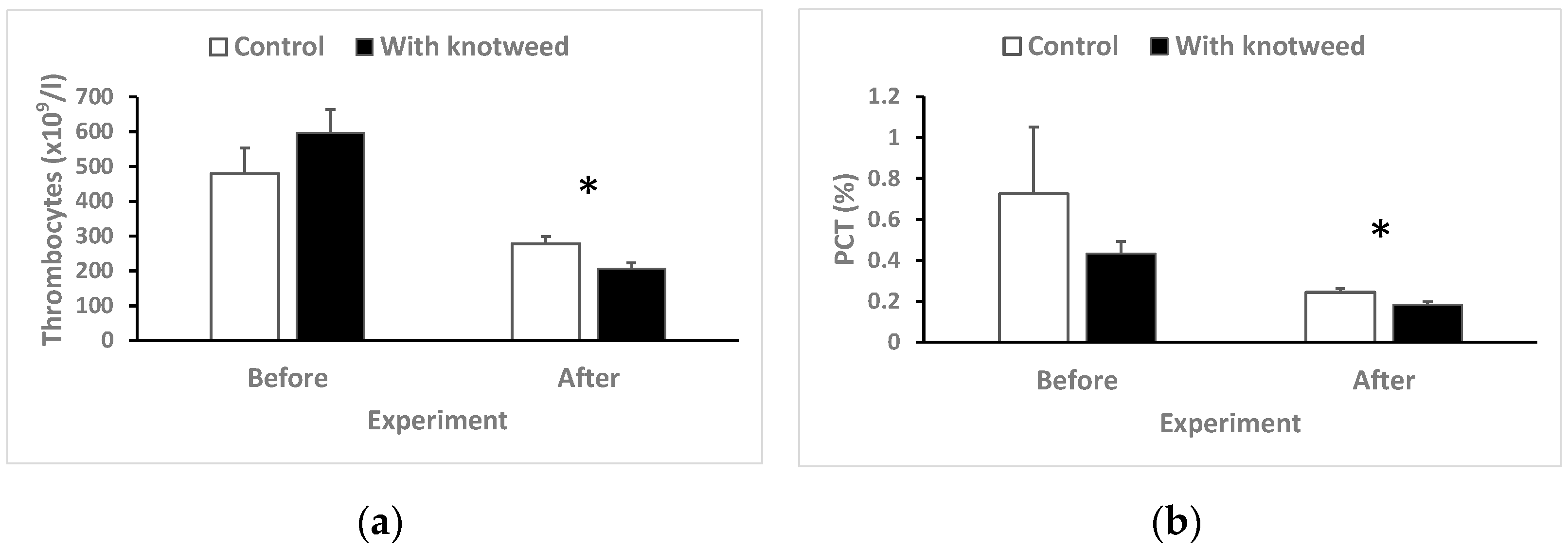
3.4.7. Thrombocytes and Plateletcrit (PCT)
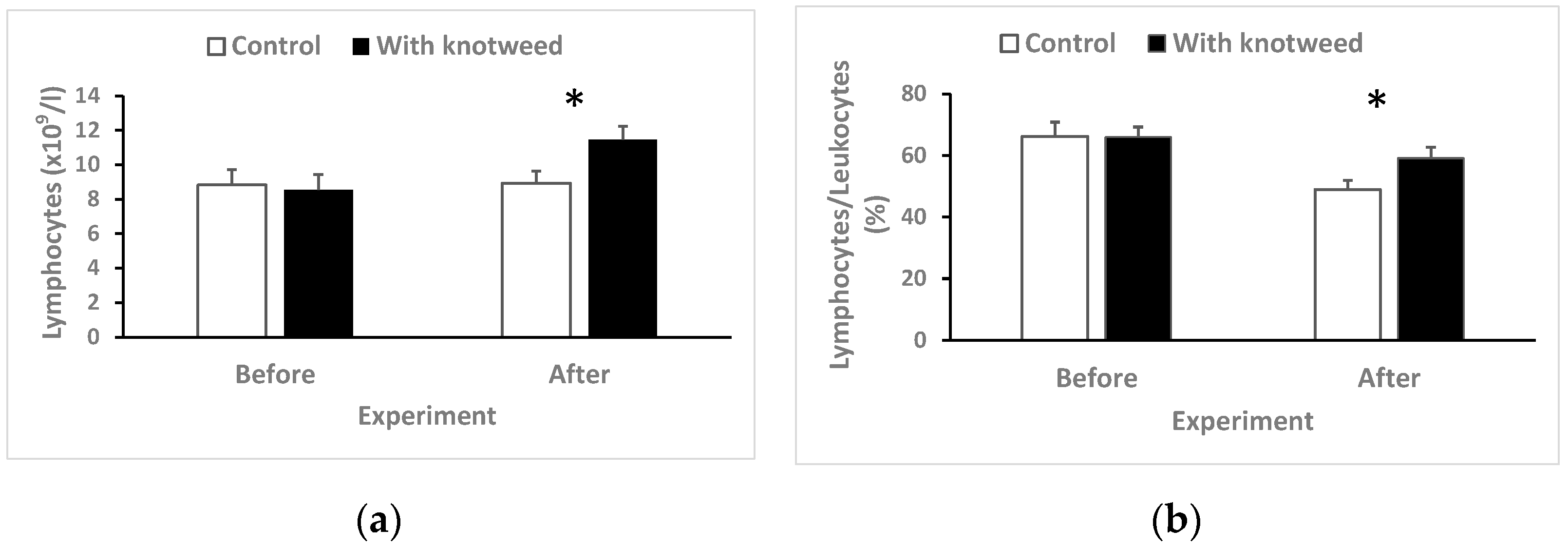
3.4.8. Lymphocytes
4. Discussion
4.1. Knotweed as a Fodder Additive
4.2. Weight Changes
4.3. Lipid Metabolism
4.4. Biochemistry and Haematology
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I.; et al. Grape pomace improves performance, antioxidant status, fecal microbiota and meat quality of piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Adebiyi, O.A.; Adeola, A.T.; Osinowo, O.A.; Brown, D.; Ng’ambi, J.W. Effects of feeding hydroponics maize fodder on per-formance and nutrient digestibility of weaned pigs. Appl. Ecol. Environ. Res. 2018, 16, 2415–2422. [Google Scholar] [CrossRef]
- Václavková, E.; Bělková, J. Kvalita masa přeštických prasat. Náš Chov 2014, 74, 36–37. [Google Scholar]
- Václavková, E.; Bělková, J.; Lustyková, A.; Frydrychová, S.; Lipenský, J.; Rozkot, M.; Truněčková, J. Breeding of Přeštice Black Pied pigs—Review. Res. Pig Breed. 2018, 12, 14–19. [Google Scholar]
- Nevrkla, P.; Václavková, E. Meat quality of indigenous Prestice Black-Pied pig and commercial hybrid pigs. Res. Pig Breed. 2018, 12, 15–18. [Google Scholar]
- Dostálová, A.; Svitáková, A.; Bureš, D.; Vališ, L.; Volek, Z. Effect of an Outdoor Access System on the Growth Performance, Carcass Characteristics, and Longissimus lumborum Muscle Meat Quality of the Prestice Black-Pied Pig Breed. Animals 2020, 10, 1244. [Google Scholar] [CrossRef]
- Claude, L. The impact of invasive knotweed species (Reynoutria spp.) on the environment: Review and research perspectives. Biol. Invasions 2017, 19, 2319–2337. [Google Scholar] [CrossRef]
- Shaw, R.; Bryner, S.; Tanner, R. The life history and host range of the Japanese knotweed psyllid, Aphalara itadori Shinji: Potentially the first classical biological weed control agent for the European Union. Biol. Control 2009, 49, 105–113. [Google Scholar] [CrossRef]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the world’s worst invasive alien species. Biol. Invasions 2014, 16, 981–985. [Google Scholar] [CrossRef]
- Brock, J.H.; Child, L.E.; De Waal, L.C.; Wade, P.M. The invasive nature of Reynoutria japonica is enhanced by vegetative regeneration from stem tissues. In Plant Invasion—General Aspects and Special Problems, 1st ed.; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 131–139. [Google Scholar]
- Horn, P.; Prach, K. Aerial biomass of Reynoutria japonica and its comparison with that of native species. Preslia 1995, 66, 345–348. [Google Scholar]
- Bailey, J.P.; Conolly, A.P. Prize-winners to pariahs. A history of Japanese knotweed s.l. (Polygonaceae) in the British Isles. Watsonia 2000, 23, 93–110. [Google Scholar]
- Barney, J.N. North American history of two invasive plant species: Phytogeographic distribution, dispersal vectors, and multiple introductions. Biol. Invasions 2006, 8, 703–717. [Google Scholar] [CrossRef]
- Sukopp, H.; Starfinger, U. Reynoutria sachalinensis in Europe, and in the Far East: A comparison of the species ecology in its native, and adventive distribution range. In Plant Invasion—General Aspects and Special Problems, 1st ed.; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 151–159. [Google Scholar]
- Frantík, T.; Kovářová, M.; Koblihová, H.; Bartůňková, K.; Nývltová, Z.; Vosátka, M. Production of medically valuable stilbenes and emodin in knotweed. Ind. Crop. Prod. 2013, 50, 237–243. [Google Scholar] [CrossRef]
- Kovářová, M.; Bartůňková, K.; Frantík, T.; Koblihová, H.; Prchlová, K.; Vosátka, M. Factors influencing the production of stilbenes by the knotweed, Reynoutria bohemica. BMC Plant Biol. 2010, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Kovářová, M.; Frantík, T.; Koblihová, H.; Bartůňková, K.; Nývltová, Z.; Vosátka, M. Effect of clone selection, nitrogen supply, leaf damage and mycorrhizal fungi on stilbene and emodin production in knotweed. BMC Plant Biol. 2011, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Tou, J.C. Resveratrol supplementation affects bone acquisition and osteoporosis: Pre-clinical evidence toward translational diet therapy. Biochim. Biophys. Acta Mol. Basis dis. 2015, 1852, 1186–1194. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.W. Effects of polydatin on platelet aggregation of rabbits. Acta Pharm. Sin. 1988, 23, 394–396. [Google Scholar]
- Luo, S.F.; Yu, C.L.; Zhang, P.W. Influences of 3,4,5-trihydroxystilbene 3§-mono-D-glucoside on beat rate and injury of cultured newborn rat myocardial cells. Acta Pharm. Sin. 1990, 11, 147–150. [Google Scholar]
- Chen, C.; Gao, J.; Wang, T.S.; Guo, C.; Yan, Y.J.; Mao, C.Y.; Gu, L.W.; Yang, Y.; Li, Z.F.; Liu, A. NMR-based Metabolomic Techniques Identify the Toxicity of Emodin in HepG2 Cells. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Chan, M.M.Y. Antimicrobial effect of resveratrol on dermatophytes and bacterial pathogens of the skin. Biochem. Pharmacol. 2002, 63, 99–104. [Google Scholar] [CrossRef]
- Docherty, J.J.; Fu, M.M.; Tsai, M. Resveratrol selectively inhibits Neisseria gonorrhoeae and Neisseria meningitidis. J. Antimicrob. Chemother. 2001, 47, 243–244. [Google Scholar] [CrossRef] [Green Version]
- Adrian, M.; Jeandet, P.; Veneau, J.; Weston, L.A.; Bessis, R. Biological activity of resveratrol, a stilbenic compound from grapevines, against Botrytis cinerea, the causal agent for gray mold. J. Chem. Ecol. 1997, 23, 1689–1702. [Google Scholar] [CrossRef]
- Bavaresco, L.; Vezzulli, S.; Battilani, P.; Giorni, P.; Pietri, A.; Bertuzzi, T. Effect of ochratoxin A-producing Aspergilli on stilbenic phytoalexin synthesis in grapes. J. Agric. Food Chem. 2003, 51, 6151–6157. [Google Scholar] [CrossRef]
- Filip, V.; Plocková, M.; Šmidrkal, J.; Špičková, Z.; Melzoch, K.; Schmidt, S. Resveratrol and its antioxidant and antimicrobial effectiveness. Food Chem. 2003, 83, 585–593. [Google Scholar] [CrossRef]
- Jung, H.J.; Hwang, I.A.; Sung, W.S.; Kang, H.; Kang, B.S.; Seu, Y.B.; Lee, D.G. Fungicidal effect of resveratrol on human infectious fungi. Arch Pharm. Res. 2005, 28, 557–560. [Google Scholar] [CrossRef]
- Schulze, K.; Schreiber, L.; Szankowski, I. Inhibiting effects of resveratrol and its glucoside piceid against Venturia inaequalis, the causal agent of apple scab. J. Agric. Food Chem. 2005, 53, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Anantaphruti, M.; Terada, M.; Ishii, A.I.; Kino, H.; Sano, M.; Kuroyanagi, M.; Fukushima, A.S. Chemotherapy of parasitic helminths XI. In vitro effects of various drugs on the motor activity of adult Schistosoma japonicum. Jpn. J. Parasitol. 1982, 31, 321–328. [Google Scholar]
- Yu, M.; Chen, T.-T.; Zhang, T.; Jia, H.-M.; Li, J.-J.; Zhang, H.-W.; Zou, Z.-M. Anti-inflammatory constituents in the root and rhizome of Polygonum cuspidatum by UPLC-PDA-QTOF/MS and lipopolysaccharide-activated RAW264.7 macrophages. J. Pharm. Biomed. Anal. 2021, 195, 113839. [Google Scholar] [CrossRef]
- Potdar, S.; Parmar, M.S.; Ray, S.D.; Cavanaugh, J.E. Protective effects of the resveratrol analog piceid in dopaminergic SH-SY5Y cells. Arch. Toxicol. 2018, 92, 669–677. [Google Scholar] [CrossRef]
- Ferry-Dumazet, H.; Garnier, O.; Mamani-Matsuda, M.; Vercauteren, J.; Belloc, F.; Billiard, C.; Dupouy, M.; Thiolat, D.; Kolb, J.P.; Marit, G.; et al. Resveratrol inhibits the growth and induces the apoptosis of both normal and leukemic hematopoietic cells. Carcinogenesis 2002, 23, 1327–1333. [Google Scholar] [CrossRef]
- Roman, V.; Billard, C.; Kern, C.; Ferry-Dumazet, H.; Izard, J.C.; Mohammad, R.; Mossalayi, D.M.; Kolb, J.P. Analysis of resveratrol-induced apoptosis in human B-cell chronic leukaemia. Br. J. Haematol. 2002, 117, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, S.; Wolter, F.; Stein, J.M. Molecular mechanisms of the chemopreventive effects of resveratrol and its analogs in carcinogenesis. Mol. Nutr. Food Res. 2005, 49, 452–461. [Google Scholar] [CrossRef]
- Wolter, F.; Ulrich, S.; Stein, J. Molecular mechanisms of the chemopreventive effects of resveratrol and its analogs in colorectal cancer: Key role of polyamines? J. Nutr. 2004, 134, 3219–3222. [Google Scholar] [CrossRef]
- El-Mowafy, A.M.; Alkhalaf, M. Resveratrol activates adenylyl-cyclase in human breast cancer cells: A novel, estrogen receptor-independent cytostatic mechanism. Carcinogenesis 2003, 24, 869–873. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.Y.; Zhang, J.L.; Qian, J.M. The effect of emodin on VEGF receptors in human colon cancer cells. Cancer Biother. Radiopharm. 2008, 23, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Hori, M.; Sasaki, Y.; Saitoh, A.; Yasuda, I.; Maekawa, T.; Uchida, T.; Asakura, K.; Nakazato, T.; Kaneda, T.; et al. Emodin has a cytotoxic activity against human multiple myeloma as a Janus-activated kinase 2 inhibitor. Mol. Cancer 2007, 6, 987–994. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.Y.; Han, J.X.; Huang, H.Y. Effects of emodin on gene expression profile in small cell lung cancer NCI-H446 cells. Chin. Med. J. 2007, 120, 1710–1715. [Google Scholar] [CrossRef] [PubMed]
- Pecere, T.; Gazzola, M.V.; Mucignat, C.; Parolin, C.; Vecchia, F.D.; Cavaggioni, A.; Basso, G.; Di-Aspro, A.; Salvato, B.; Carli, M.; et al. Aloe-emodin is a new type of anticancer agent with selective activity against neuroectodermal tumors. Cancer Res. 2000, 60, 2800–2804. [Google Scholar]
- Jeong, E.T.; Jin, M.H.; Kim, M.S.; Chang, Y.H.; Park, S.G. Inhibition of melanogenesis by piceid isolated from Polygonum cuspidatum. Arch. Pharm. Res. 2010, 33, 1331–1338. [Google Scholar] [CrossRef]
- Pezzuto, J.M. Resveratrol as an inhibitor of carcinogenesis. Pharm. Biol. 2008, 46, 443–573. [Google Scholar] [CrossRef]
- Kerem, Z.; Bilkis, I.; Flaishman, M.A.; Sivan, U. Antioxidant activity and inhibition of alpha-glucosidase by trans-resveratrol, piceid, and a novel trans-stilbene from the roots of Israeli Rumex bucephalophorus L. J. Agric Food Chem. 2006, 54, 1243–1247. [Google Scholar] [CrossRef]
- Dal-Pan, A.; Blanc, S.; Aujard, F. Resveratrol suppresses body mass gain in a seasonal non-human primate model of obesity. BMC Physiol. 2010, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Brasnyo, P.; Molnar, G.A.; Mohas, M.; Marko, L.; Laczy, B.; Cseh, J.; Mikolas, E.; Szijarto, I.A.; Merei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Aktpathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Szkudelska, K.; Szkudeslki, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Ge, X.; Li, H.; Tang, Y.; Su, S. miR-144 is the epigenetic target for emodin to ameliorate oxidative stress induced by dietary oxidized fish oil via Nrf2 signaling in Wuchang bream, Megalobrama amblycephala. Aquaculture 2021, 534, 736357. [Google Scholar] [CrossRef]
- Alberdi, G.; Rodriguez, V.M.; Miranda, J.; Macarulla, M.T.; Arias, N.; Andres-Lacueva, C.; Portillo, M.P. Changes in white adipose tissue metabolism induced by resveratrol in rats. Nutr. Metab. 2011, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Ecolan, P.; Plantard, L.; Gueguen, N. New insights into muscle fibre types in the pig. J. Histochem. Cytochem. 2002, 50, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Luo, J.; Yu, B.; Zheng, P.; Huang, Z.; Mao, X.; He, J.; Yu, J.; Chen, J.; Chen, D. Dietary resveratrol supplementation improves meat quality of finishing pigs through changing muscle fibre characteristics and antioxidative status. Meat Sci. 2015, 102, 15–21. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, J.; Yu, B.; Chen, J.; Chen, D. Effects of resveratrol on lipid metabolism in muscle and adipose tissues: A reevaluation in a pig model. J. Funct. Foods 2015, 14, 590–595. [Google Scholar] [CrossRef]
- Macarulla, M.T.; Alberdi, G.; Gomez, S.; Tueros, I.; Bald, C.; Rodriguez, V.M.; Martínez, J.A.; Portillo, M.P. Effects of different doses of resveratrol on body fat and serum parameters in rats fed a hypercaloric diet. J. Physiol. Biochem. 2009, 65, 369–376. [Google Scholar] [CrossRef]
- Qiao, Y.; Sun, J.; Xia, S.; Tang, X.; Shi, Y.; Le, G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014, 5, 1241–1249. [Google Scholar] [CrossRef]
- EU decision 2005/1/ES. Commission decision of 27 December 2004 authorising methods for grading pig carcases in the Czech Republic. Off. J. Eur. Union 2005, 1, 8–11. [Google Scholar]
- CSN ISO 1444. International Organization for Standardization. Meat and Meat Products. Determination of Free Fat Content; Czech Standards Institute: Prague, Czech Republic, 1997. [Google Scholar]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- CSN ISO 5509. Animal and Vegetable Fats and Oils. Preparation of Methyl Esters of Fatty Acids; The Czech Office for Standards, Metrology and Testing Prague: Prague, Czech Republic, 1994. [Google Scholar]
- CSN ISO 5508. Animal and Vegetable Fats and Oils. Analysis by Gas Chromatography of Methyl Esters of Fatty Acids; The Czech Office for Standards, Metrology and Testing Prague: Prague, Czech Republic, 1994. [Google Scholar]
- Jha, R.; Leterme, P. Feed ingredients differing in fermentable fibre and indigestible protein content affect fermentation metabolites and faecal nitrogen excretion in growing pigs. Animal 2012, 6, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Hooda, P.S. (Ed.) Trace Elements in Soil, 1st ed.; Wiley: London, UK, 2010. [Google Scholar]
- Linden, J. High Fibre Swine Diets. The Pig Site. Available online: https://www.thepigsite.com/articles/high-fibre-swine-diets (accessed on 22 January 2021).
- Patience, J.F. Preparing for the Inevitable Increase in Fibre Content in Practical Pig Diets—NPB #09-020. Research Report, Animal Science—Nutritional Efficiency Consortium. 2011. Available online: https://www.pork.org/wp-content/uploads/2011/11/09-020-PATIENCE-ISU.pdf (accessed on 22 January 2021).
- Lindberg, J.E. Fibre effects in nutrition and gut health in pigs. J. Anim. Sci. Biotechnol. 2014, 5, 15. [Google Scholar] [CrossRef]
- Václavková, E. Přeštické černostraké. Náš Chov 2017, 77, 12–14. [Google Scholar]
- Mátlová, V. Přeštické plemeno prasat jako genetický zdroj. In Perspektivy přeštického černostrakatého plemene prasat v podmínkách globalizovaného trhu; Výzkumný ústav živočišné výroby: Praha, Czech Republic, 2014. [Google Scholar]
- Jedlička, M. V ZD Mladotice se s přeštickými prasaty počítá. Náš Chov 2015, 7, 55. [Google Scholar]
- Zhang, H.; Li, Y.; Su, W.; Ying, Z.; Zhou, L.; Zhang, L.; Wang, T. Resveratrol attenuates mitochondrial dysfunction in the liver of intrauterine growth retarded suckling piglets by improving mitochondrial biogenesis and redox status. Mol. Nutr. Food Res. 2017, 61, 1600653. [Google Scholar] [CrossRef]
- Rivera, L.; Morón, R.; Zarzuelo, A.; Galisteo, M. Long-term resveratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem. Pharm. 2009, 77, 1053–1063. [Google Scholar] [CrossRef] [Green Version]
- Aubin, M.C.; Lajoie, C.; Clement, R.; Gosselin, H.; Calderone, A.; Perrault, L.P. Female rats fed a high-fat diet were associated with vascular dysfunction and cardiac fibrosis in the absence of overt obesity and hyperlipidemia: Therapeutic potential of resveratrol. J. Pharm. Exp. 2008, 325, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, N.L.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Rocha, K.K.; Souza, G.A.; Ebaid, G.X.; Seiva, F.R.; Cataneo, A.C.; Novelli, E.L. Resveratrol toxicity: Effects on risk factors for atherosclerosis and hepatic oxidative stress in standard and high-fat diets. Food Chem. Toxicol. 2009, 47, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, L.; Mourot, J.; Insausti, K.; Mendizabal, J.A.; Arana, A. Comparative description of growth, fat deposition, carcass and meat quality characteristics of Basque and Large White pigs. Anim. Res 2005, 54, 33–42. [Google Scholar] [CrossRef]
- Meydani, M.; Hasan, T.S. Dietary Polyphenols and Obesity. Nutrients 2010, 2, 737. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Lee, R.H.; Leon, Y.-J.; Park, M.-S.; Shin, J.-C.; Kim, S.-H.; Jeong, J.I.; Kang, H.; Choi, N.-J.; Seo, K.S.; et al. Proteomic Assessment of the Relevant Factors Affecting Pork Meat Quality Associated with Longissimus dorsi Muscles in Duroc Pigs. Asian Australas. J. Anim. Sci. 2016, 29, 1653–1663. [Google Scholar] [CrossRef] [Green Version]
- Čechová, M.; Mikule, V.; Sládek, L.; Bečková, R. Intramuscular fat content as related to selected carcass value indicators in pigs of sire breeds bred in Czech Republic. Anim. Sci. Pap. Rep. 2006, 24, 65–72. [Google Scholar]
- Yu, K.; Shu, G.; Yuan, F.; Zhu, X.; Gao, P.; Wang, S.; Wang, L.; Xi, Q.; Zhang, S.; Zhang, Y. Fatty acid and transcriptome profiling of longissimus dorsi muscles between pig breeds differing in meat quality. Int. J. Biol. Sci. 2013, 9, 108–118. [Google Scholar] [CrossRef]
- Kasprzyk, A.; Tyra, M.; Babicz, M. Fatty acid profile of pork from a local and a commercial breed. Arch. Anim. Breed. 2015, 58, 379–385. [Google Scholar] [CrossRef]
- Nevrkla, P.; Kapelański, W.; Václavková, E.; Hadaš, Z.; Cebulska, A.; Horký, P. Meat quality and fatty acid profile of pork and backfat from an indigenous breed and a commercial hybrid of pigs. Ann. Anim. Sci 2017, 174, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- Serra, A.; Buccioni, A.; Rodriguez-Estrada, M.T.; Conte, G.; Cappucci, A.; Mele, M. Fatty acid composition, oxidation status and volatile organic compounds in “Colonnata” lard from Large White or Cinta Senese pigs as affected by curing time. Meat Sci. 2014, 97, 504–512. [Google Scholar] [CrossRef]
- Ma, J.I.; Sun, D.-N. Prediction of monounsaturated and polyunsaturated fatty acids of various processed pork meats using improved hyperspectral imaging technique. Food Chem. 2020, 321, 126695. [Google Scholar] [CrossRef]
- He, Z.; Huang, Y.; Li, H.; Qin, G.; Wang, T.; Yang, J. Effect of high-pressure treatment on the fatty acid composition of intramuscular lipid in pork. Meat Sci. 2012, 90, 170–175. [Google Scholar] [CrossRef]
- Čítek, J.; Stupka, R.; Okrouhlá, M.; Vehovský, K.; Brzobohatý, L.; Šprysl, M.; Stádník, L. Effect of dietary linseed and corn supplement on the fatty acid content in the pork loin and back fat tissue. Czechj. Anim. Sci. 2015, 60, 319–326. [Google Scholar] [CrossRef] [Green Version]
- FAO. Fats and Fatty Acids in Human Nutrition. Report of an Expert Consultation. Food Nutr. Pap. 2010, 91, 1–166. [Google Scholar]
- Komprda, T.; Kuchtík, J.; Jarošová, A.; Dračková, E.; Zemánek, L.; Filipčík, R. Meat quality characteristics of lambs of three organically raised breeds. Meat Sci. 2012, 91, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Robina, A.; Viguera, J.; Perez-Palacios, T.; Mayoral, A.I.; Vivo, J.M.; Guillen, M.T.; Ruiz, J. Carcass and meat quality traits of Iberian pigs as affected by sex and crossbreeding with different Duroc genetic lines. Span. J. Agric. Res. 2013, 11, 1057–1067. [Google Scholar] [CrossRef] [Green Version]
- Madzimure, J.; Chimonyo, M.; Hugo, A.; Bakare, A.G.; Katiyatiya, C.L.F.; Muchenje, V. Physico-chemical quality attributes and fatty acid profiles of pork from Windsnyer and Large White gilts. S. Afr. J. Anim. Sci. 2017, 47, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Lassen, E.; Fettman, M. Laboratory evaluation of lipids. In Veterinary Hematology and Clinical Chemistry, 1st ed.; Thrall, M.A., Weiser, G., Allison, R.W., Campbell, T.W., Eds.; Willey Black: London, UK, 2006; pp. 421–429. [Google Scholar]
- Elmadhun, N.Y.; Sabe, A.A.; Robich, M.P.; Chu, L.M.; Lassaletta, A.D.; Sellke, F.W. The pig as a valuable model for testing the effect of resveratrol to prevent cardiovascular disease. Ann. N. Y. Acad. Sci. 2013, 1290, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; González-Sarrías, A.; Larrosa, M.; Vallejo, F.; Pallarés, F.J.; Lucas, R.; Morales, J.C.; Tomás-Barberán, J.A.; García-Conesa, M.-T.; et al. Effects of long-term consumption of low doses of resveratrol on diet-induced mild hypercholesterolemia in pigs: A transcriptomic approach to disease prevention. J. Nutr. Biochem. 2012, 23, 829–837. [Google Scholar] [CrossRef]
- Wang, B.; Sun, J.; Li, L.; Zheng, J.; Shi, Y.; Le, G. Regulatory effects of resveratrol on glucose metabolism and T-lymphocyte subsets in the development of high-fat diet induced obesity in C57BL/6 mice. Food Funct. 2014, 5, 1452–1463. [Google Scholar] [CrossRef]
- Bai, L.; Pang, W.-J.; Yang, Y.-J.; Yang, G.-S. Modulation of Sirt1 by resveratrol and nicotinamide alters proliferation and differentiation of pig preadipocytes. Mol. Cell. Biochem. 2008, 307, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Burges, T.A.; Robich, M.P.; Chu, L.M.; Bianchi, C.; Sellke, F.W. Improving Glucose Metabolism with Resveratrol in a Swine Model of Metabolic Syndrom Through Alterration of Signaling Pathways in the Liver and Skeletal Muscle. Arch. Surg. 2011, 146, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Doubek, J.; Šlosárková, S.; Řeháková, K.; Bouda, J.; Scheer, P.; Piperisová, I.; Tomenendálová, J.; Matalová, E. Interpretace základních biochemických a hematologických nálezů u zvířat, 2nd ed.; NOVIKO s.r.o.: Brno, Czechia, 2010. [Google Scholar]
- Walker, H.; Hall, W.; Hurst, J.; Busher, J. Serum Albumin and Globulin. In Clinical Methods: The History, Physical, and Laboratory Examinations, 3rd ed.; Walker, K., Hall, D., Hurst, W., Eds.; Butterworth-Heinemann: Boston, MA, USA, 1990. [Google Scholar]
- Lassen, E. Laboratory Evaluation of Plasma and Serum Proteins. In Veterinary Hematology and Clinical Chemistry; Thrall, M.A., Baker, D., Campbell, T.W., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 355–416. [Google Scholar]
- Wiliams, L.D.; Burdock, G.A.; Edwards, J.A.; Beck, M.; Bausch, J. Safety studies conducted on high-purity trans-resveratrol in experimental animals. Food Chem. Toxicol. 2019, 47, 2170–2182. [Google Scholar] [CrossRef]
- Fettman, M.; Rebar, A. Laboratory Evaluation of Renal Function. In Veterinary Hematology and Clinical Chemistry; Thrall, M.A., Baker, D., Campbell, T.W., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 301–328. [Google Scholar]
- Sun-Wook, K.; Hee-Won, J.; Cheol-Ho, K.; Kwang-Il, K.; Ho Jun, C.; Hajeong, L. A New Equation to Estimate Muscle Mass from Creatinin and Cystatin C. PLoS ONE 2016, 11, e0148495. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, N.; Jiang, N.; Xue, W.; Guo, X.; Xu, X.; Fang, W.; Wang, H.; Hua, E. Resveratrol treatment improves plasma and blood glucose concentration and lipid metabolism in high-fat-fed C57BL/6J mice. Eur. Food Res. Technol. 2016, 242, 1849–1856. [Google Scholar] [CrossRef]
- Hao, J.; Chen, C.; Huang, K.; Huang, J.; Li, J.; Liu, P.; Huang, H. Polydatin improves glucose and lipid metabolism in experimental diabetes through activating the Akt signaling pathway. Eur. J. Pharmacol. 2014, 745, 152–165. [Google Scholar] [CrossRef]
- Faghihzadeh, F.; Adibi, P.; Rafiei, R.; Hekmatdoost, A. Resveratrol supplementation improves inflammatory biomarkers in patients with non-alcoholic fatty liver disease. Nutr. Res. 2014, 34, 837–843. [Google Scholar] [CrossRef]
- Crowell, J.; Korytko, P.; Morrissey, R.; Booth, T.D.; Levine, B.S. Resveratrol Associated Renal Toxicity. Toxicol. Sci. 2004, 82, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Baker, D. Diagnosis of Disorders of Hemostasis. In Veterinary Hematology and Clinical Chemistry; Thrall, M.A., Baker, D., Campbell, T.W., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 179–196. [Google Scholar]
- Thrall, M.A.; Weiser, G.; Jain, N. Laboratory Evaluation of Bone Marrow. In Veterinary Hematology and Clinical Chemistry; Thrall, M.A., Baker, D., Campbell, T.W., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 149–178. [Google Scholar]
- Doubek, J.; Volný, T.; Lojek, A.; Knotková, Z.; Kotrbáček, V.; Scheer, P.; Holešovská, Z. Effect of Stilbene Resveratrol on Hematological Indices of Rats. Acta Vet. 2005, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Thushara, R.; Hemshekhar, M.; Santhosh, M.; Devaraja, S.; Kemparaju, K.; Girish, K.S. Differential Action of Phytochemicals on Platelet Apoptosis: A Biological Overview. Curr. Med. Chem. 2013, 20, 1018–1027. [Google Scholar] [CrossRef]
- Feng, Y.; Zhon, W.; Wu, Q.; Li, X.; Zhao, W.; Zou, J. Low Dose of Resveratrol Enhanced Immune Response of Mice. Acta Pharm. Sin. 2002, 23, 893–897. [Google Scholar]
- Gao, X.; Xu, Y.X.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. Immunomodulatory activity of resveratrol: Supression of lymfocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production. Biochem. Pharmacol. 2001, 62, 1299–1308. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | ČOS (EW) [kg] | A1 [kg] | KPB [kg] |
|---|---|---|---|
| NK (n = 16) | 715.8 | 826.5 | 3287.0 |
| WK (n = 15) | 810.0 | 900.0 | 3477.5 |
| Feeding duration | 2019.6.20–2019.8.12 | 2019.8.13–2019.9.9 | 2019.9.10–2019.12.8 |
| MLLT | Backfat | |||
|---|---|---|---|---|
| WK | NK | WK | NK | |
| SFA | 43.34 ± 0.44 * | 41.90 ± 0.46 | 42.61 ± 0.50 | 41.28 ± 0.43 |
| nonSFA | 56.66 ± 0.44 * | 58.10 ± 0.46 | 57.39 ± 0.50 | 58.72 ± 0.43 |
| MUFA | 46.88 ± 0.45 * | 45.25 ± 0.61 | 44.38 ± 0.27 * | 41.62 ± 0.60 |
| PUFA | 9.78 ± 0.43 * | 12.85 ± 0.61 | 13.02 ± 0.42 * | 17.10 ± 0.64 |
| PUFA n-3 | 1.09 ± 0.08 * | 1.69 ± 0.10 | 1.37 ± 0.05 * | 2.28 ± 0.12 |
| PUFA n-6 | 8.69 ± 0.37 * | 11.16 ± 0.51 | 11.65 ± 0.37 * | 14.83 ± 0.53 |
| n-6/n-3 | 8.12 ± 0.29 * | 6.69 ± 0.14 | 8.56 ± 0.18 * | 6.68 ± 0.30 |
| MLLT | Backfat | |||||||
|---|---|---|---|---|---|---|---|---|
| Gilts | Boars | Gilts | Boars | |||||
| WK | NK | WK | NK | WK | NK | WK | NK | |
| SFA | 42.37 ± 0.86 | 41.25 ± 0.49 | 44.07 ± 0.23 | 42.41 ± 0.70 | 41.70 ± 0.97 | 41.41 ± 0.52 | 43.29 ± 0.38 | 41.18 ± 0.68 |
| MUFA | 46.71 Aa ± 0.99 | 43.85 AB ± 0.64 | 47.00 A ± 0.38 | 46.30 Aa ± 0.79 | 44.35 AB ± 0.26 | 39.43 B ± 0.66 | 44.40 AB ± 0.44 | 43.26 Bb ± 0.25 |
| PUFA | 10.91 AB ± 0.56 | 14.91 CD ± 0.79 | 8.93 A ± 0.45 | 11.31 ACD ± 0.29 | 13.95 BD ± 0.80 | 19.16 E ± 0.50 | 12.31 BC ± 0.25 | 15.56 D ± 0.63 |
| MUFA/PUFA | 4.35 Aba ± 0.28 | 2.99 CEb ± 0.18 | 5.36 A ± 0.28 | 4.12 BDa ± 0.15 | 3.23 CDb ± 0.17 | 2.07 E ± 0.09 | 3.62 BC ± 0.10 | 2.81 CE ± 0.12 |
| PUFA/SFA | 0.26 Aba ± 0.02 | 0.36 CEb ± 0.02 | 0.20 A ± 0.01 | 0.27 Aa ± 0.01 | 0.34 BC ± 0.03 | 0.46 Ec ± 0.02 | 0.28 AC ± 0.01 | 0.38 CE ± 0.02 |
| MUFA/SFA | 1.11 ± 0.04 | 1.06 ± 0.02 | 1.07 ± 0.01 | 1.10 ± 0.04 | 1.07 ± 0.03 | 0.95 ± 0.02 | 1.03 ± 0.02 | 1.05 ± 0.02 |
| PUFA n-3 | 1.30 Aa ± 0.13 | 2.01 BCb ± 0.14 | 0.94 A ± 0.04 | 1.45 ABc ± 0.07 | 1.43 AB ± 0.11 | 2.60 C ± 0.09 | 1.32 A ± 0.05 | 2.04 Cd ± 0.15 |
| PUFA n-6 | 9.61 AB ± 0.48 | 12.90 CD ± 0.66 | 8.00 A ± 0.41 | 9.86 Abd ± 0.24 | 12.53 CDc ± 0.70 | 16.56 E ± 0.42 | 10.99 BCa ± 0.22 | 13.52 Db ± 0.49 |
| n-6/n-3 | 7.60 ± 0.57 | 6.47 B ± 0.21 | 8.51 a ± 0.23 | 6.86 b ± 0.19 | 8.84 Aa ± 0.20 | 6.38 Bb ± 0.09 | 8.35 a ± 0.26 | 6.90 ± 0.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maděra, P.; Kovářová, M.; Frantík, T.; Filipčík, R.; Novák, J.; Vencl, Š.; Maděrová, L.; Rozkot, M.; Kuchařová, S.; Václavková, E.; et al. Effect of Knotweed in Diet on Physiological Changes in Pig. Agriculture 2021, 11, 169. https://doi.org/10.3390/agriculture11020169
Maděra P, Kovářová M, Frantík T, Filipčík R, Novák J, Vencl Š, Maděrová L, Rozkot M, Kuchařová S, Václavková E, et al. Effect of Knotweed in Diet on Physiological Changes in Pig. Agriculture. 2021; 11(2):169. https://doi.org/10.3390/agriculture11020169
Chicago/Turabian StyleMaděra, Petr, Marcela Kovářová, Tomáš Frantík, Radek Filipčík, Jan Novák, Štěpán Vencl, Lucie Maděrová, Miroslav Rozkot, Stanislava Kuchařová, Eva Václavková, and et al. 2021. "Effect of Knotweed in Diet on Physiological Changes in Pig" Agriculture 11, no. 2: 169. https://doi.org/10.3390/agriculture11020169
APA StyleMaděra, P., Kovářová, M., Frantík, T., Filipčík, R., Novák, J., Vencl, Š., Maděrová, L., Rozkot, M., Kuchařová, S., Václavková, E., Truněčková, J., Volková, J., Nývltová, Z., & Bartoš, M. (2021). Effect of Knotweed in Diet on Physiological Changes in Pig. Agriculture, 11(2), 169. https://doi.org/10.3390/agriculture11020169