
Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Retrieval and Bioinformatic Analysis
2.2. Plant Materials and RNA Extraction
2.3. Expression Analysis
3. Results
3.1. Genome-Wide Identification and Phylogenetic Analysis of PINs in Cucurbitaceae Species
3.2. Cis-Element and Gene Structure Analysis of Watermelon PINs
3.3. In Silico Expression of PINs in Watermelon Tissues
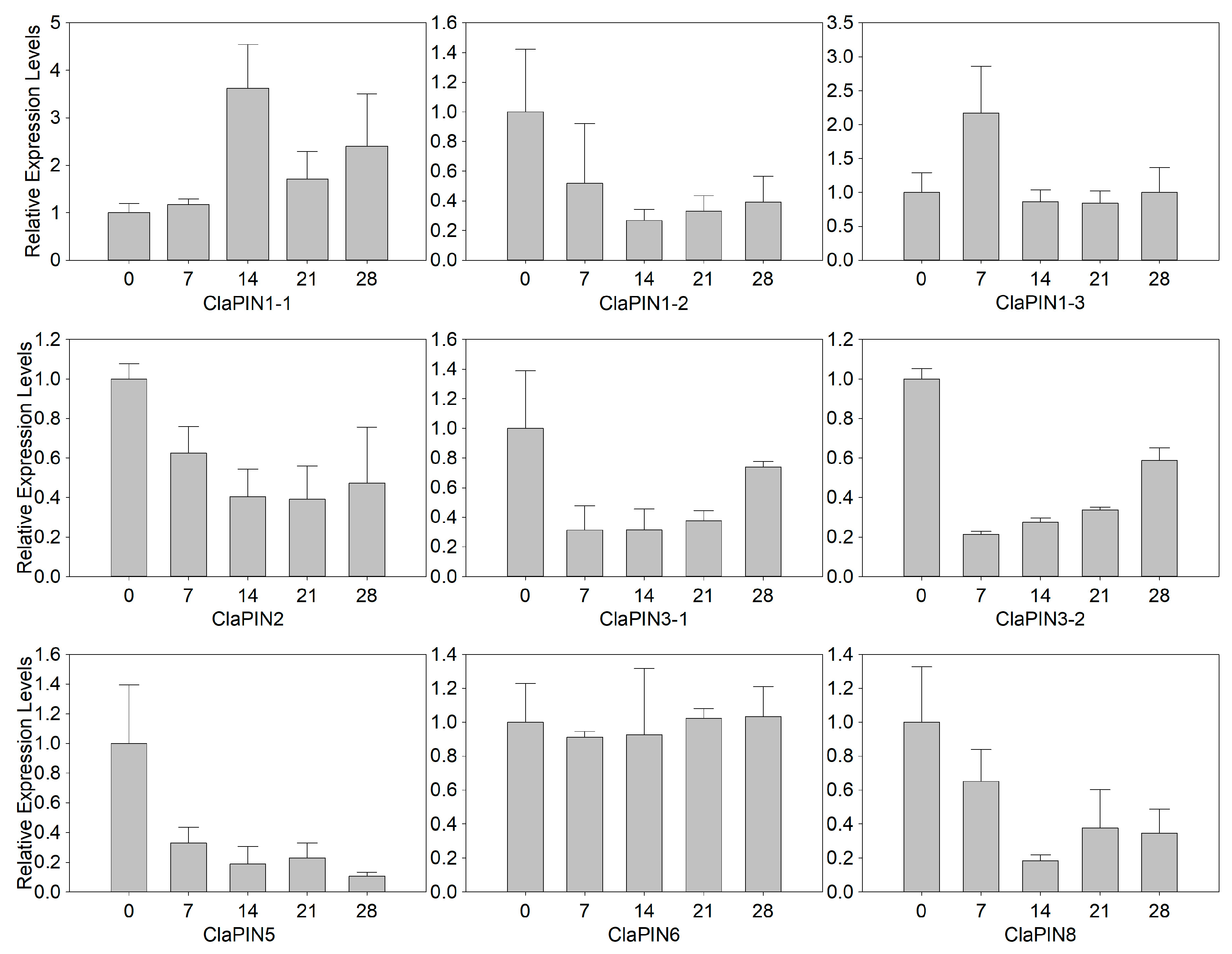
3.4. Expression Profiles of Watermelon PINs during Shoot Regeneration
4. Discussion
4.1. Identification of PINs in Cucurbitaceae Species
4.2. Putative Regulators in ClaPINs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Jiao, Y. Auxin and above-ground meristems. J. Exp. Bot. 2018, 69, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Krecek, P.; Skupa, P.; Libus, J.; Naramoto, S.; Tejos, R.; Friml, J.; Zazímalová, E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol. 2009, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.J.; Luo, J. The PIN-FORMED Auxin Efflux Carriers in Plants. Int. J. Mol. Sci. 2018, 19, 2759. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Wang, B.; Moreno, I.; Duplakova, N.; Simon, S.; Carraro, N.; Reemmer, J.; Pencik, A.; Chen, X.; Tejos, R.; et al. ER-localized auxin transporter PIN8 regulates auxin homeostasis and male gametophyte development in Arabidopsis. Nat. Commun. 2012, 3, 941. [Google Scholar] [CrossRef] [Green Version]
- Mravec, J.; Skupa, P.; Bailly, A.; Hoyerova, K.; Krecek, P.; Bielach, A.; Petrasek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.D. Subcellular homeostasis of phytohormone auxin is mediated by the ER localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Skupa, P.; Viaene, T.; Zwiewka, M.; Tejos, R.; Klima, P.; Carna, M.; Rolcik, J.; De Rycke, R.; Moreno, I.; et al. PIN6 auxin transporter at endoplasmic reticulum and plasma membrane mediates auxin homeostasis and organogenesis in Arabidopsis. New Phytol. 2016, 211, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Rakusová, H.; Abbas, M.; Han, H.; Song, S.; Robert, H.S.; Friml, J. Termination of shoot gravitropic responses by auxin feedback on PIN3 polarity. Curr. Biol. 2016, 26, 3026–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigo, G.; Ayaydin, F.; Tietz, O.; Zsigmond, L.; Kovacs, H.; Pay, A.; Salchert, K.; Darula, Z.; Medzihradszky, K.F.; Szabados, L.; et al. Inactivation of plasma membrane-localized CDPK-RELATED KINASE5 decelerates PIN2 exocytosis and root gravitropic response in Arabidopsis. Plant Cell 2013, 25, 1592–1608. [Google Scholar] [CrossRef] [Green Version]
- Rosquete, M.R.; Waidmann, S.; Kleine, V.J. PIN7 auxin carrier has a preferential role in terminating radial root expansion in Arabidopsis thaliana. Int. J. Mol. Sci. 2018, 19, 1238. [Google Scholar] [CrossRef] [Green Version]
- Xi, W.; Gong, X.; Yang, Q.; Yu, H.; Liou, Y.C. Pin1At regulates PIN1 polar localization and root gravitropism. Nat. Commun. 2016, 7, 10430. [Google Scholar] [CrossRef] [Green Version]
- Haga, K.; Sakai, T. PIN auxin efflux carriers are necessary for pulse-induced but not continuous light-induced phototropism in Arabidopsis. Plant Physiol. 2012, 160, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Zadnikova, P.; Petrasek, J.; Marhavy, P.; Raz, V.; Vandenbussche, F.; Ding, Z.; Schwarzerova, K.; Morita, M.T.; Tasaka, M.; Hejatko, J.; et al. Role of PIN-mediated auxin efflux in apical hook development of Arabidopsis thaliana. Development 2010, 137, 607–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, K.; Ueda, J.; Komaki, M.K.; Bell, C.J.; Shimura, Y. Requirement of the auxin polar transport system in early stages of Arabidopsis floral bud formation. Plant Cell 1991, 3, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Bosco, C.; Dovzhenko, A.; Liu, X.; Woerner, N.; Rensch, T.; Eismann, M.; Eimer, S.; Hegermann, J.; Paponov, I.A.; Ruperti, B.; et al. The endoplasmic reticulum localized PIN8 is a pollen-specific auxin carrier involved in intracellular auxin homeostasis. Plant J. 2012, 71, 860–870. [Google Scholar] [CrossRef]
- Willige, B.C.; Chory, J. A current perspective on the role of AGCVIII kinases in PIN-mediated apical hook development. Front. Plant Sci. 2015, 6, 767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Takahashi, M.; Shibasaki, K.; Wu, S.; Inaba, T.; Tsurumi, S.; Baskin, T.I. Gravitropism of Arabidopsis thaliana roots requires the polarization of PIN2 toward the root tip in meristematic cortical cells. Plant Cell 2010, 22, 1762–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Liu, Y.; Maere, S.; Lee, E.; Van, I.G.; Xie, Z.; Xuan, W.; Lucas, J.; Vassileva, V.; Kitakura, S.; et al. A coherent transcriptional feed-forward motif model for mediating auxin-sensitive PIN3 expression during lateral root development. Nat. Commun. 2015, 6, 8821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friml, J.; Benková, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jürgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Vieten, A.; Vanneste, S.; Wisniewska, J.; Benková, E.; Benjamins, R.; Beeckman, T.; Luschnig, C.; Friml, J. Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression. Development 2005, 132, 4521–4531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations. Available online: Faostat.fao.org (accessed on 6 January 2021).
- Ellis, A.C.; Dudenbostel, T.; Crowe-White, K. Watermelon Juice: A Novel Functional Food to Increase Circulating Lycopene in Older Adult Women. Plant Foods Hum. Nutr. 2019, 74, 200–203. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, S.; Sun, H.; Wang, X.; Wu, S.; Lin, T.; Ren, Y.; Gao, L.; Deng, Y.; Zhang, J.; et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits. Nat. Genet. 2019, 51, 1616–1623. [Google Scholar] [CrossRef]
- Choi, P.S.; Soh, W.Y.; Kim, Y.S.; Yoo, O.J.; Liu, J.R. Genetic transformation and plant regeneration of watermelon using Agrobacterium tumefaciens. Plant Cell Rep. 1994, 13, 344–348. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, L.; Cui, X.; Zhang, J.; Guo, S.; Li, M.; Zhang, H.; Ren, Y.; Gong, G.; Zong, M.; et al. Engineering herbicide-resistant watermelon variety through CRISPR/Cas9-mediated base-editing. Plant Cell Rep. 2018, 37, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Huang, X.; Bao, Y.; Wang, B.; Liu, L.; Dai, L.; Chen, J.; An, X.; Sun, Y.; Peng, D. Genome-wide identification and expression profiling of WUSCHEL-related homeobox (WOX) genes during adventitious shoot regeneration of watermelon (Citrullus lanatus). Acta Physiol. Plant. 2015, 37, 224. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, X.; Bao, Y.; Wang, B.; Zeng, H.; Cheng, W.; Tang, M.; Li, Y.; Ren, J.; Sun, Y. Genome-wide identification of SAUR genes in watermelon (Citrullus lanatus). Physiol. Mol. Biol. Plants 2017, 23, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, D.; Kareem, A.; Durgaprasad, K.; Sreeraj, E.; Sugimoto, K.; Prasad, K. Shoot regeneration: A journey from acquisition of competence to completion. Curr. Opin. Plant Biol. 2018, 41, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef]
- Krogan, N.T.; Marcos, D.; Weiner, A.I.; Berleth, T. The auxin response factor MONOPTEROS controls meristem function and organogenesis in both the shoot and root through the direct regulation of PIN genes. New Phytol. 2016, 212, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Dong, W.; Zhan, Y.; Huang, Z.A.; Li, Z.; Kim, I.S.; Zhang, C. Genome-wide identification and expression analysis of ClLAX, ClPIN and ClABCB genes families in Citrullus lanatus under various abiotic stresses and grafting. BMC Genet. 2017, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Cucurbit Genomics Database. Available online: Cucurbitgenomics.org (accessed on 6 January 2021).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- ProtParam tool. Available online: Web.expasy.org/protparam/ (accessed on 6 January 2021).
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- DNAMAN-Bioinformatics Solutions. Available online: www.lynnon.com (accessed on 6 January 2021).
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools—An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.D.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E.; et al. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Elorduy Vergara, L.; Palme, J.; Marín-de la Rosa, N.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A.; et al. Extensive signal integration by the phytohormone protein network. Nature 2020, 583, 271–276. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID. | CuGenDB Accession | Chromosome Location | Coding Sequence (bp) | Predicted Protein (aa) | Molecular Weight | Theoretical pI | CELLO Localization | |
|---|---|---|---|---|---|---|---|---|
| 97103_v2 | 97103_v1 | |||||||
| ClaPIN1-1 | Cla97C04G072260.1 | Cla012098 | Chr04: 19082426–19084800(−) | 1878 | 625 | 67109.39 | 8.22 | PlasmaMembrane (4.260) |
| ClaPIN1-2 | Cla97C04G077450.1 | Cla018455 | Chr04: 25019277–25022350(+) | 1860 | 619 | 67585.84 | 9.13 | PlasmaMembrane (4.643) |
| ClaPIN1-3 | Cla97C06G123780.1 | Cla018924 | Chr06: 26050523–26053439(+) | 1824 | 607 | 66418.71 | 9.19 | PlasmaMembrane (4.680) |
| ClaPIN2 | Cla97C06G123240.1 | Cla018871 | Chr06: 25556363–25559538(+) | 1938 | 645 | 70749.52 | 9.28 | PlasmaMembrane (4.073) |
| ClaPIN3-1 | Cla97C01G006350.1 | Cla003909 | Chr01: 6303561–6306905(−) | 1926 | 641 | 69902.79 | 7.12 | PlasmaMembrane (4.561) |
| ClaPIN3-2 | Cla97C07G134150.1 | Cla011708-Cla011709 | Chr07: 9679960–9684183(−) | 1872 | 623 | 68190.85 | 8.67 | PlasmaMembrane (4.735) |
| ClaPIN5 | Cla97C11G217020.1 | Cla010530 | Chr11: 20331421–20339597(−) | 1113 | 370 | 40169.63 | 7.61 | PlasmaMembrane (4.639) |
| ClaPIN6 | Na | Cla015026 | Chr09: 9050290–9056689(+) | 1449 | 482 | 53345.31 | 9.25 | PlasmaMembrane (4.712) |
| ClaPIN8 | Cla97C10G197710.1 | Cla017028 | Chr10: 27449992–27452031(−) | 1071 | 356 | 38907.61 | 9.53 | PlasmaMembrane (4.913) |
| CsaPIN1-1 | CsaV3_1G004350.1 | Chr01: 2730713–2733712(−) | 1827 | 608 | 66630.87 | 9.09 | PlasmaMembrane (4.798) | |
| CsaPIN1-2 | CsaV3_1G007160.1 | Chr01: 4542181–4545627(−) | 1857 | 618 | 67443.68 | 9.13 | PlasmaMembrane (4.630) | |
| CsaPIN1-3 | CsaV3_4G029470.1 | Chr04: 19013177–19015651(+) | 1791 | 596 | 63704.54 | 8.77 | PlasmaMembrane (4.476) | |
| CsaPIN2 | CsaV3_1G032010.1 | Chr01: 19030317–19033521(+) | 1938 | 645 | 70831.58 | 9.29 | PlasmaMembrane (4.272) | |
| CsaPIN3-1 | CsaV3_5G013380.1 | Chr05: 9982996–9987048(−) | 1890 | 629 | 68479.02 | 8.5 | PlasmaMembrane (4.767) | |
| CsaPIN3-2 | CsaV3_5G028620.1 | Chr05: 23739663–23744022(−) | 1911 | 636 | 69462.29 | 7.12 | PlasmaMembrane (4.600) | |
| CsaPIN5-1 | CsaV3_2G009610.1 | Chr02: 6112179–6116586(+) | 1116 | 371 | 40371.92 | 7.04 | PlasmaMembrane (4.592) | |
| CsaPIN5-2 | CsaV3_2G009700.1 | Chr02: 6189126–6193716(+) | 1191 | 396 | 43043.53 | 7.57 | PlasmaMembrane (4.916) | |
| CsaPIN8 | CsaV3_3G041710.1 | Chr03: 34023847–34026847(+) | 1071 | 356 | 38980.69 | 9.59 | PlasmaMembrane (4.895) | |
| CmePIN1-1 | MELO3C017357.2.1 | Chr02: 24004740–24008031(+) | 1776 | 591 | 64524.14 | 8.5 | PlasmaMembrane (4.654) | |
| CmePIN1-2 | MELO3C019102.2.1 | Chr08: 13382844–13385789(+) | 1836 | 611 | 65374.38 | 8.75 | PlasmaMembrane (4.526) | |
| CmePIN1-3 | MELO3C002132.2.1 | Chr12: 26099882–26104026(+) | 1929 | 642 | 70303.64 | 9.01 | PlasmaMembrane (4.183) | |
| CmePIN2 | MELO3C017414.2.1 | Chr02: 23469145–23472527(+) | 1872 | 623 | 68443.69 | 9.22 | PlasmaMembrane (3.928) | |
| CmePIN3-1 | MELO3C005326.2.1 | Chr09: 20480725–20484941(−) | 1911 | 636 | 69499.35 | 7.15 | PlasmaMembrane (4.624) | |
| CmePIN3-2 | MELO3C018353.2.1 | Chr10: 18871332–18876126(+) | 1887 | 628 | 68548.23 | 8.69 | PlasmaMembrane (4.729) | |
| CmePIN5 | MELO3C008726.2.1 | Chr05: 18006724–18012271(−) | 1209 | 402 | 43622.16 | 6.23 | PlasmaMembrane (4.858) | |
| CmePIN8 | MELO3C009892.2.1 | Chr04: 28346209–28350422(−) | 1038 | 345 | 37657.14 | 9.54 | PlasmaMembrane (4.939) | |
| Gene ID. | Forward Primers | Reverse Primers | References |
|---|---|---|---|
| ClaPIN1-1 | CTCTGTTTCTGACCTCTACC | TACGGCGAGGGCGGCAAGGA | [32] |
| ClaPIN1-2 | TCTTGCTGTTCTTGCCGTTT | GCTCAGAAATCAGCATTCTG | |
| ClaPIN1-3 | TACATTATCAGACTTTTACC | GTCCAGATACCGAGCACCAC | |
| ClaPIN2 | ACATATACGATGTATTCTCAG | GTGGAAGGAGAGTAATGG | |
| ClaPIN3-1 | ATCATGGAAGGATCTTTACA | AAATCCCAAGAAAGAACAAC | |
| ClaPIN3-2 | ACTGTCTTGACGGCGGTCAT | AATTGTAAGAGCAACAAGCA | |
| ClaPIN5 | CCATGGCCCCATTGTACTTC | AAGACGAGAACGATAACGAT | |
| ClaPIN6 | AAGGTGATGTGTGCGATGGT | TTGCCGGAGAAGAAAACAGC | |
| ClaPIN8 | TTCGTTGTCTGATGTCTATC | TATCAGTTGCAGGGTCATTT | |
| GAPDH | TGGAAGAATCGGTAGGTTGG | CTGTCACTGTTTTTGGCGTC | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, H.; Zhang, N.; Xie, Z.; Deng, S.; Yi, L.; Huang, X. Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus). Agriculture 2021, 11, 447. https://doi.org/10.3390/agriculture11050447
Shang H, Zhang N, Xie Z, Deng S, Yi L, Huang X. Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus). Agriculture. 2021; 11(5):447. https://doi.org/10.3390/agriculture11050447
Chicago/Turabian StyleShang, Huaiguo, Na Zhang, Zhouli Xie, Siyu Deng, Licong Yi, and Xing Huang. 2021. "Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus)" Agriculture 11, no. 5: 447. https://doi.org/10.3390/agriculture11050447
APA StyleShang, H., Zhang, N., Xie, Z., Deng, S., Yi, L., & Huang, X. (2021). Genome-Wide Identification and Expression of the PIN Auxin Efflux Carrier Gene Family in Watermelon (Citrullus lanatus). Agriculture, 11(5), 447. https://doi.org/10.3390/agriculture11050447