
Effect of Spring Frost Damage on Apple Fruit (Malus domestica Borkh.) Inner Quality at Harvest




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction and Determination of Individual Sugars and Organic Acids
2.3. Extraction and Determination of Individual Phenolic Compounds
2.4. Statistics
3. Results
3.1. Sugars and Organic Acids in Apple Flesh
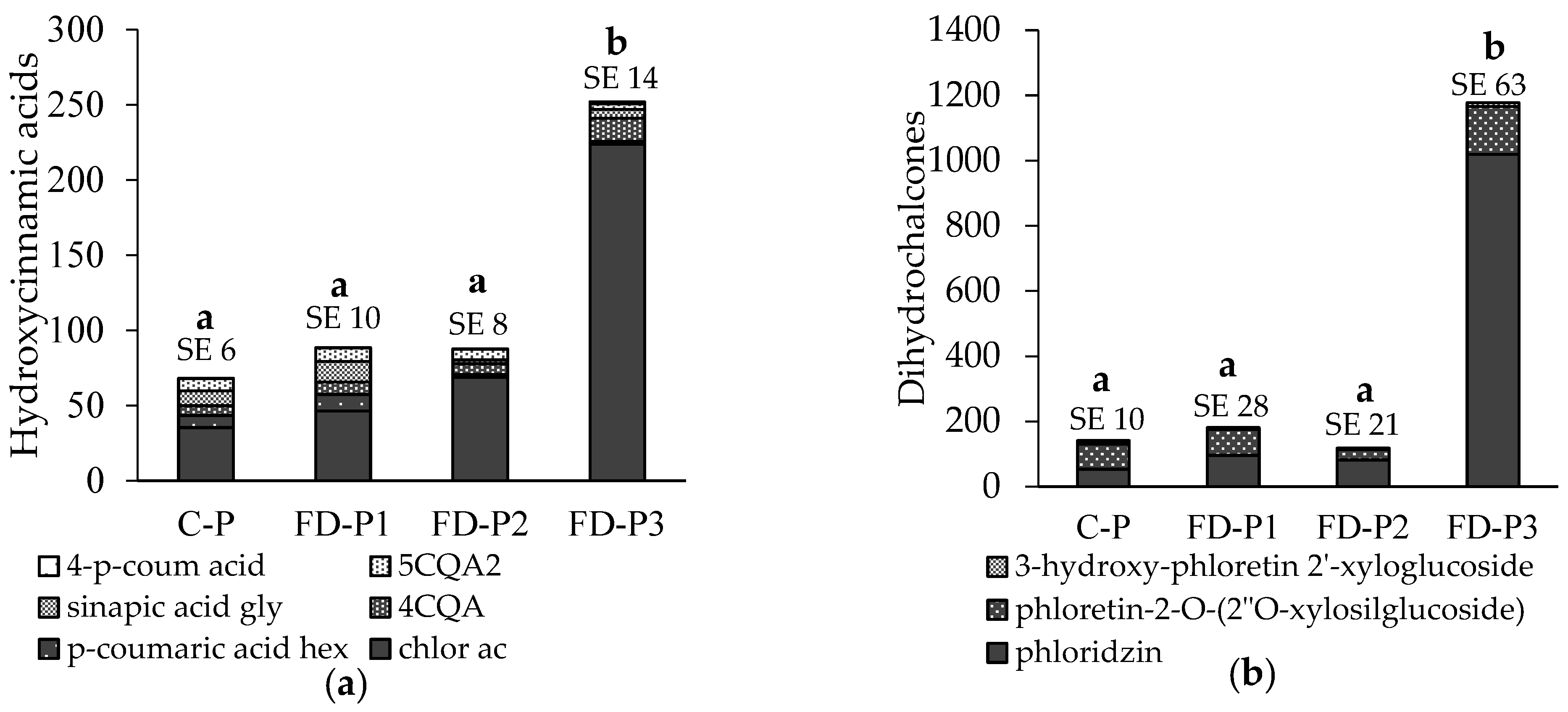
3.2. Phenolic Content in Apple Flesh and Peel
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigo, J. Spring frosts in deciduous fruit trees—Morphological damage and flower hardiness. Sci. Hortic. 2000, 85, 55–173. [Google Scholar] [CrossRef]
- Byers, R.E.; Marini, R.P. Influence of blossom and fruit thinning on peach flower bud tolerance to an early spring freeze. HortScience 1994, 29, 146–148. [Google Scholar] [CrossRef]
- Eccel, E.; Rea, R.; Caffarra, A.; Crisci, A. Risk of spring frost to apple production under future climate scenarios: The role of phenological acclimation. Int. J. Biometeorol. 2009, 53, 273–286. [Google Scholar] [CrossRef]
- Blanke, M.M.; Kunz, A. Effects of climate change on pome fruit phenology and precipitation. Acta Hortic. 2011, 922, 381–386. [Google Scholar] [CrossRef]
- Ginzberg, I.; Stern, R.A. Strengthening fruit-skin resistance to growth strain by application of plant growth regulators. Sci. Hortic. 2016, 198, 150–153. [Google Scholar] [CrossRef]
- Khanal, B.P.; Grimm, E.; Knoche, M. Russeting in apple and pear: A plastic periderm replaces a stiff cuticule. AoB Plants 2013, 5, pls048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musacchi, S.; Serra, S. Apple fruit quality: Overview on pre-harvest factors. Sci. Hortic. 2018, 234, 409–430. [Google Scholar] [CrossRef]
- Busatto, N.; Matsumoto, D.; Tadiello, A.; Vrhovsek, U.; Costa, F. Multifaced analyses disclose the role of fruit size and skin-russeting in the accumulation pattern of phenolic compounds in apple. PLoS ONE 2019, 14, e0219354. [Google Scholar] [CrossRef] [Green Version]
- Hyson, D.A. A Comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Andre, C.M.; Larsen, L.; Burgess, E.J.; Jensen, D.J.; Cooney, J.M.; Evers, D.; Zhang, J.; Perry, N.B.; Laing, W.A. Unusual immuno-modulatory triterpene-caffeates in the skins of russeted varieties of apples and pears. J. Agric. Food Chem. 2013, 61, 2773–2779. [Google Scholar] [CrossRef]
- Faust, M.; Shear, C.B. Russeting of apples, an interpretive review. Hortic. Sci. 1972, 7, 233–235. [Google Scholar]
- Eccher, T.; Hajnaajar, H. Fluctuations of entogenous gibberellin A4 and A7 content in apple fruits with different sensitivity to russet. Acta Hortic. 2006, 727, 537–543. [Google Scholar] [CrossRef]
- Hoffmann, H.; Rath, T. Future bloom and blossom frost risk for Malus domestica considering climate model and impact model uncertainties. PLoS ONE 2013, 8, e75033. [Google Scholar]
- Warrington, I.J.; Fulton TAHalligan, E.A.; de Silva, H.N. Apple Fruit Growth and Maturity are Affected by Early Season Temperatures. J. Am. Soc. Hortic. Sci. 1999, 124, 468–477. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, T.; Ogawa, H.; Fukuda, N.; Mariguchi, T. Changes in the taste and textural attributes of apples in response to climate change. Sci. Rep. 2013, 3, 2418. [Google Scholar] [CrossRef] [Green Version]
- Zupan, A.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R. Sugar and phenol content in apple with or without watercore. J. Sci. Food Agric. 2015, 96, 2845–2850. [Google Scholar] [CrossRef] [PubMed]
- Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R. Parameters of inner quality of the apple scab resistant and susceptible apple cultivars (Malus domestica Borkh.). Sci. Hortic. 2007, 114, 37–44. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, P.; Cheng, L. Developmental changes of carbohydrates, organic acids, amino acids, and phenolic compounds in ‘Honeycrisp’ apple flesh. Food Chem. 2010, 123, 1013–1018. [Google Scholar] [CrossRef]
- Cebulj, A.; Cunja, V.; Mikulic-Petkovsek, M.; Veberic, R. Importance of metabolite distribution in apple fruit. Sci. Hortic. 2017, 214, 214–220. [Google Scholar] [CrossRef]
- Cheng, L.; Zhou, R.; Reidel, E.J.; Sharkey, T.D.; Dandekar, A.M. Antisense inhibition of sorbitol synthesis leads to up-regulation of starch synthesis without altering CO2 assimilation in apple leaves. Planta 2005, 220, 767–776. [Google Scholar] [CrossRef]
- Whetter, J.M.; Taper, C.D. Occurrence of sorbitol (D-glucitol) and certain related sugars in germinating seeds and developing seedlings of Malus. Can. J. Bot. 1966, 44, 51–55. [Google Scholar] [CrossRef]
- Loescher, W.H.; Everard, J.D. Regulation of sugar alcohol biosynthesis. In Photosynthesis. Advances in Photosynthesis and Respiration; Leegood, R.C., Sharkey, T.D., von Caemmerer, S., Eds.; Springer: Dordrecht, The Netherlands, 2000; Volume 9, pp. 275–299. [Google Scholar]
- Li, F.; Lei, H.; Zhao, X.; Shen, X.; Lium, A.; Li, T. Isolation and characterisation of two sorbitol transporter gene promoters in micropropagated apple plants (Malus × domestica) regulated by drought stress. Plant Growth Regul. 2012, 68, 475–482. [Google Scholar] [CrossRef]
- Aprea, E.; Charles, M.; Endrizzi, I.; Corollaro, M.L.; Betta, E.; Biasioli, F.; Gasperi, F. Sweet taste in apples: The role of sorbitol, individual sugars, organic acids and volatile compounds. Sci. Rep. 2017, 7, 44950. [Google Scholar] [CrossRef]
- Fernie, A.R.; Martinoia, E. Malate. Jack of all trades or master of a few? Phytochemistry 2009, 70, 828–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.-G.; Ma, Q.-J.; Sun, C.-H.; Sun, M.-H.; You, C.-X.; Hao, Y.-J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plantarum. 2016, 156, 201–214. [Google Scholar] [CrossRef]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.-G.; Sun, C.-H.; Ma, Q.-J.; You, C.-X.; Cheng, L.; Hao, Y.-J. MdMYB1 regulates anthocyanin and malate accumulation by directly facilitating their transport into vacuoles in apples. Plant Physiol. 2016, 170, 1315–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.-G.; Li, Y.-Y.; Zhang, Q.-Y.; Li, M.; Sun, C.-H.; Yu, J.-Q.; Hao, Y.-J. The R2R3-MYB transcription factor MdMYB 73 is involved in malate accumulation and vacuolar acidification in apple. Plant J. 2017, 91, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Jia, D.; Shen, F.; Wang, Y.; Wu, T.; Xu, X.; Zhang, X.; Han, Z. Apple fruit acidity is genetically diversified by natural variations in three hierarchical epistatic genes: MdSAUR37, MdPP2CH and MdALMTII. Plant J. 2018, 95, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Evans, K.; Guan, Y.; Luby, J.J.; Rosyara, U.R.; Howard, N.P.; Bassil, N.; Bink, M.C.A.M.; van de Weg, W.E.; Peace, C.P. Two large-effect QTLs, Ma and Ma3, determine genetic potential for acidity in apple fruit: Breeding insights from a multi-family study. Tree Genet. Genomes 2019, 15, 18. [Google Scholar] [CrossRef] [Green Version]
- Rymenants, M.; van de Weg, E.; Auwerkerken, A.; De Wit, I.; Czech, A.; Nijland, B.; Heuven, H.; De Storme, N.; Keulemans, W. Detection of QTL for apple fruit acidity and sweetness using sensorial evaluation in multiple pedigreed full-sib families. Tree Genet. Genomes 2020, 16, 71. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules; Elsevier: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar]
- Lattanzio, V. Phenolic compounds: Introduction. In Natural Products; Ramawat, K., Merillon, J.M., Eds.; Springer: Berlin, Germany, 2013; pp. 1543–1580. [Google Scholar]
- Kundu, A.; Vadassery, J. Chlorogenic acid-mediated chemical defence of plants against insect herbivores. Plant Biol. 2019, 21, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Slatnar, A.; Mikulic Petkovsek, M.; Halbwirth, H.; Stampar, F.; Stich, K.; Veberic, R. Enzyme activity of the phenylpropanoid pathway as a response to apple scab infection. Ann. Appl. Biol. 2010, 156, 449–456. [Google Scholar] [CrossRef]
- Zupan, A.; Mikulic-Petkovsek, M.; Cunja, V.; Stampar, F.; Veberic, R. Comparison of phenolic composition of healthy apple tissues and tissues affected by bitter pit. J. Agric. Food Chem. 2013, 61, 12066–12071. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, B.L.; Zhong, G.-Y.; Brown, S.K. Increased phloridzin content associated with russeting in apple (Malus domestica (Suckow) Borkh.) fruit. Genet. Resour. Crop Evol. 2018, 65, 2135–2149. [Google Scholar] [CrossRef]
- Gosch, C.; Halbwirth, H.; Stich, K. Biosythesis, distribution and physiological relevance in plants. Phytochemistry 2010, 71, 838–843. [Google Scholar] [CrossRef]
- Baldi, P.; Moser, M.; Brilli, M.; Vrhovsek, U.; Pindo, M.; Si-Ammour, A. Fine-tuning of the flavonoid and monolignol pathways during apple early fruit development. Planta 2017, 245, 1021–1035. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sucrose | Glucose | Fructose | Sorbitol | Sum of Sugars | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C-F | 41.52 ± 2.17 | a | 6.31 ± 0.34 | a | 46.51 ± 0.83 | a | 0.91 ± 0.08 | a | 95.24 ± 2.55 | a |
| FD-F1 | 35.10 ± 2.00 | a | 6.84 ± 0.52 | a | 52.79 ± 1.76 | b | 0.94 ± 0.06 | a | 95.64 ± 2.95 | a |
| FD-F2 | 38.77 ± 1.63 | a | 6.51 ± 0.23 | a | 55.43 ± 1.17 | b | 1.21 ± 0.04 | b | 101.92 ± 1.25 | a |
| Malic Acid | Citric Acid | Shikimic Acid | Sum of Organic Acids | |||||
|---|---|---|---|---|---|---|---|---|
| C-F | 4.78 ± 0.06 | a | 0.36 ± 0.04 | ab | 0.02 ± 0 | a | 5.09 ± 0.10 | a |
| FD-F1 | 4.42 ± 0.34 | a | 0.35 ± 0.05 | a | 0.02 ± 0 | a | 4.78 ± 0.35 | a |
| FD-F2 | 5.41 ± 0.17 | b | 0.47 ± 0.04 | b | 0.02 ± 0 | a | 5.83 ± 0.23 | b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cebulj, A.; Mikulič-Petkovšek, M.; Veberič, R.; Jakopic, J. Effect of Spring Frost Damage on Apple Fruit (Malus domestica Borkh.) Inner Quality at Harvest. Agriculture 2022, 12, 14. https://doi.org/10.3390/agriculture12010014
Cebulj A, Mikulič-Petkovšek M, Veberič R, Jakopic J. Effect of Spring Frost Damage on Apple Fruit (Malus domestica Borkh.) Inner Quality at Harvest. Agriculture. 2022; 12(1):14. https://doi.org/10.3390/agriculture12010014
Chicago/Turabian StyleCebulj, Anka, Maja Mikulič-Petkovšek, Robert Veberič, and Jerneja Jakopic. 2022. "Effect of Spring Frost Damage on Apple Fruit (Malus domestica Borkh.) Inner Quality at Harvest" Agriculture 12, no. 1: 14. https://doi.org/10.3390/agriculture12010014
APA StyleCebulj, A., Mikulič-Petkovšek, M., Veberič, R., & Jakopic, J. (2022). Effect of Spring Frost Damage on Apple Fruit (Malus domestica Borkh.) Inner Quality at Harvest. Agriculture, 12(1), 14. https://doi.org/10.3390/agriculture12010014