
Broad-Spectrum Resistance to Leaf Rust in the Argentinean Wheat Cultivar “Klein Proteo” Is Controlled by LrKP Located on Chromosome 2BS

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Leaf Rust Inoculation
2.2. Bulked Segregant RNA-Seq (BSR-Seq)
2.3. Development of Molecular Markers
2.4. PCR Validation of Lr Genes in KP
2.5. Gene Expression Analysis
3. Results
3.1. Seedlings of “Klein Proteo” Showed High Resistance to Most of the Pt Pathotypes in China
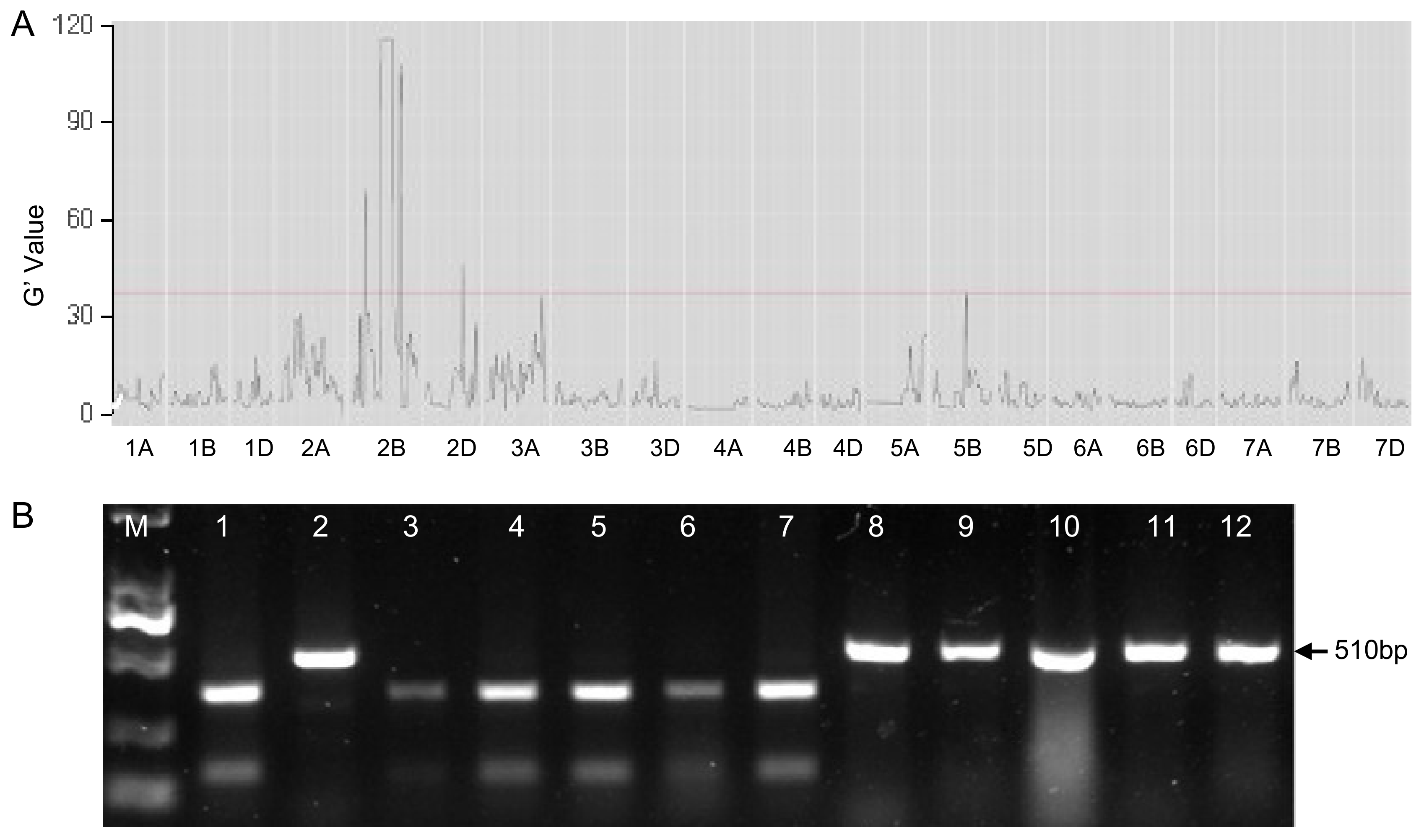
3.2. BSR-seq Analysis on “KP × ZZ5389” F2:3 Lines Revealed a Resistant Locus on Chromosome 2B
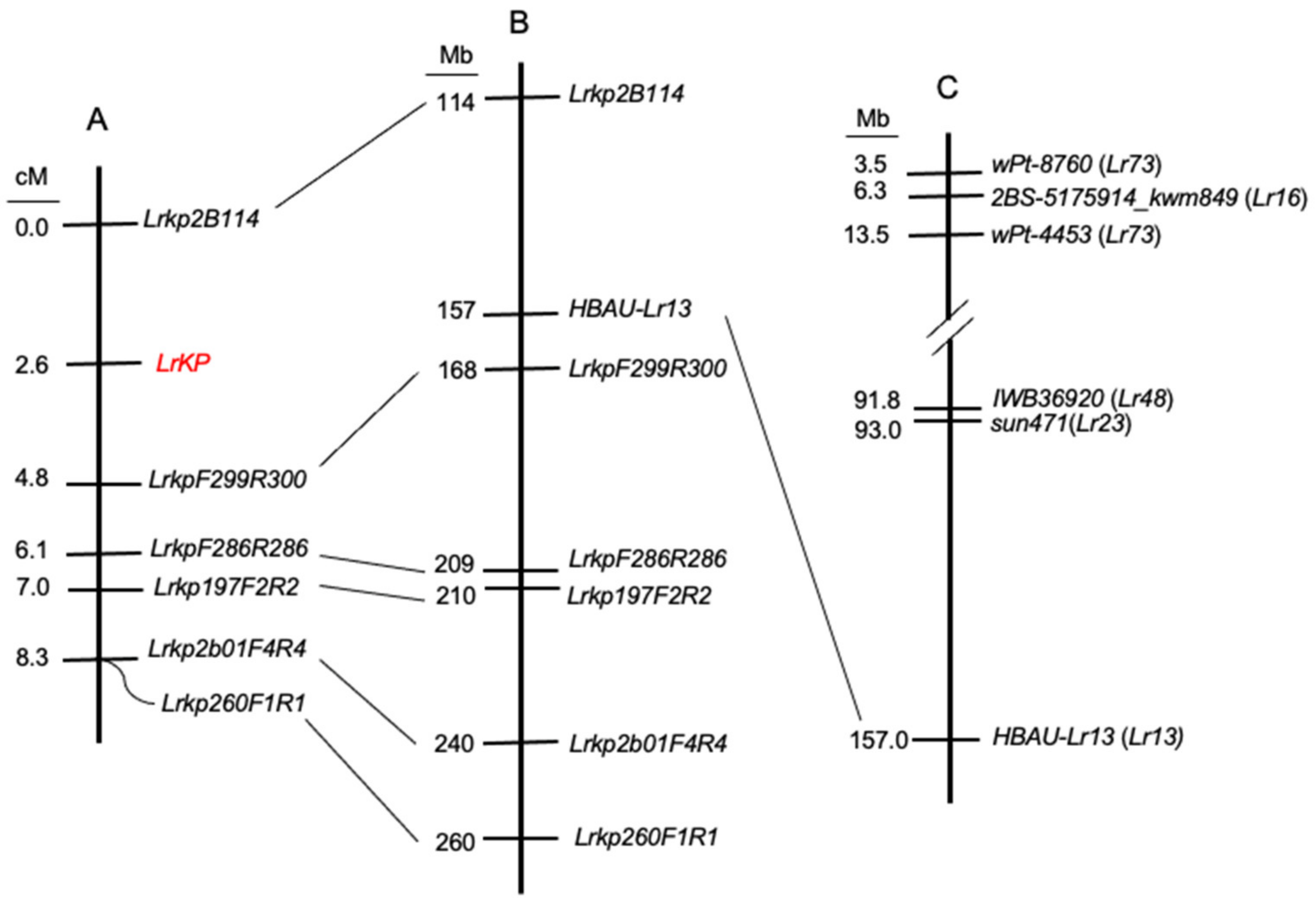
3.3. LrKP Was Mapped to a 114–168 Mb Region on Chromosome 2BS Using the Designed CAPS Markers
3.4. The Previously Cloned Lr13 Gene Located in the LrKP Interval
3.5. Lr13 Was Identified as a Differentially Expressed Gene (DEG) in the LrKP Interval
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bolton, M.; Kolmer, J.; Garvin, D. Wheat leaf rust caused by Puccinia triticina. Mol. Plant Pathol. 2008, 9, 563–575. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Crossa, J.; Yuen, J.; Djurle, A. Effect of leaf rust on grain yield and yield traits of durum wheats with race-specific and slow-rusting resistance to leaf rust. Plant Dis. 2006, 90, 1065–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.A.; Twga, B.; Zhang, P.; Li, Z.F.; Liu, D. Identification of leaf rust resistance genes in common wheat varieties from China and foreign countries. J. Integr. Agric. 2021, 20, 1302–1313. [Google Scholar] [CrossRef]
- Figueroa, M.; Hammond-Kosack, K.E.; Solomon, P.S. A review of wheat diseases-a field perspective. Mol. Plant Pathol. 2018, 19, 1523–1536. [Google Scholar] [CrossRef] [PubMed]
- Feuillet, C.; Travella, S.; Stein, N.; Albar, L.; Nublat, A.; Keller, B. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L.) genome. Proc. Natl. Acad. Sci. USA 2003, 100, 15253–15258. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Brooks, S.A.; Li, W.; Fellers, J.P.; Trick, H.N.; Gill, B.S. Map-based cloning of leaf rust resistance gene Lr21 from the large and polyploid genome of bread wheat. Genetics 2003, 164, 655–664. [Google Scholar] [CrossRef]
- Thind, A.K.; Wicker, T.; Šimková, H.; Fossati, D.; Moullet, O.; Brabant, C.; Vrána, J.; Doležel, J.; Krattinger, S.G. Rapid cloning of genes in hexaploid wheat using cultivar-specific long-range chromosome assembly. Nat. Biotechnol. 2017, 35, 793–796. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Li, M.; Zhang, P.; Yin, G.; Zhang, H.; Gebrewahid, T.W.; Zhang, J.; Dong, L.; Liu, D.; Liu, Z.; et al. High-temperature wheat leaf rust resistance gene Lr13 exhibits pleiotropic effects on hybrid necrosis. Mol. Plant 2021, 14, 1029–1032. [Google Scholar] [CrossRef]
- Lin, G.; Chen, H.; Tian, B.; Sehgal, S.K.; Singh, L.; Xie, J.; Rawat, N.; Juliana, P. Cloning of the broadly effective wheat leaf rust resistance gene Lr42 transferred from Aegilops tauschii. Nat. Commun. 2022, 13, 3044. [Google Scholar] [CrossRef]
- Kolodziej, M.C.; Singla, J.; Sánchez-Martín, J.; Zbinden, H.; Šimková, H.; Karafiátová, M.; Doležel, J.; Gronnier, J.; Poretti, M.; Glauser, G.; et al. A membrane-bound ankyrin repeat protein confers race-specific leaf rust disease resistance in wheat. Nat. Commun. 2021, 12, 956. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Lagudah, E.S.; Spielmeyer, W.; Singh, R.P.; Huerta-Espino, J.; Mcfadden, H.; Bossolini, E.; Selter, L.L.; Keller, B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 2009, 323, 1360–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S.; et al. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef]
- Hewitt, T.; Zhang, J.; Huang, L.; Upadhyaya, N.; Li, J.; Park, R.; Hoxha, S.; McIntosh, R.; Lagudah, E.; Zhang, P. Wheat leaf rust resistance gene Lr13 is a specific Ne2 allele for hybrid necrosis. Mol. Plant 2021, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Kassa, M.T.; You, F.M.; Hiebert, C.W.; Pozniak, C.J.; Fobert, P.R.; Sharpe, A.G.; Menzies, J.G.; Humphreys, D.G.; Harrison, N.R.; Fellers, J.P.; et al. Highly predictive SNP markers for efficient selection of the wheat leaf rust resistance gene Lr16. BMC Plant Biol. 2017, 17, 45. [Google Scholar] [CrossRef] [Green Version]
- Chhetri, M.; Bariana, H.; Wong, D.; Sohail, Y.; Hayden, M.; Bansal, U. Development of robust molecular markers for marker-assisted selection of leaf rust resistance gene Lr23 in common and durum wheat breeding programs. Mol. Breed. 2017, 37, 21. [Google Scholar] [CrossRef]
- Seyfarth, R.; Feuillet, C.; Schachermayr, G.; Winzeler, M.; Keller, B. Development of a molecular marker for the adult plant leaf rust resistance gene Lr35 in wheat. Theor. Appl. Genet. 1999, 99, 554–560. [Google Scholar] [CrossRef]
- Nsabiyera, V.; Qureshi, N.; Bariana, H.S.; Wong, D.; Forrest, K.L.; Hayden, M.J.; Bansal, U.K. Molecular markers for adult plant leaf rust resistance gene Lr48 in wheat. Mol. Breed. 2016, 36, 65. [Google Scholar] [CrossRef]
- Park, R.F.; Mohler, V.; Nazari, K.; Singh, D. Characterisation and mapping of gene Lr73 conferring seedling resistance to Puccinia triticina in common wheat. Theor. Appl. Genet. 2014, 127, 2041–2049. [Google Scholar] [CrossRef]
- Vanzetti, L.S.; Campos, P.; Demichelis, M.; Lombardo, L.A.; Aurelia, P.R.; Vaschetto, L.M.; Bainottl, C.T.; Helguera, M. Identification of leaf rust resistance genes in selected Argentinean bread wheat cultivars by gene postulation and molecular markers. Electron. J. Biotechnol. 2011, 14, 9. [Google Scholar] [CrossRef]
- Cobo, N.; Pflüger, L.; Chen, X.; Dubcovsky, J. Mapping QTL for resistance to new virulent races of wheat stripe rust from two Argentinean wheat cultivars. Crop Sci. 2018, 58, 2470–2483. [Google Scholar] [CrossRef]
- Zhao, J.; Bi, W.; Zhao, S.; Su, J.; Li, M.; Ma, L.; Yu, X.; Wang, X. Wheat apoplast-localized lipid transfer protein TaLTP3 enhances defense responses against Puccinia triticina. Front. Plant Sci. 2021, 12, 771806. [Google Scholar] [CrossRef]
- Roelfs, A.P.; Singh, R.P.; Saari, E.E. Rust Diseases of Wheat: Concepts and Methods of Disease Management; CIMMYT: Batan, Mexico, 1992; p. 43. [Google Scholar]
- Zhou, H.; Xia, X.; He, Z.; Li, X.; Wang, C.; Li, Z.; Liu, D. Molecular mapping of leaf rust resistance gene LrNJ97 in Chinese wheat line Neijiang 977671. Theor. Appl. Genet. 2013, 126, 2141–2147. [Google Scholar] [CrossRef]
- Gebrewahid, T.W.; Zhang, P.; Zhou, Y.; Yan, X.; Xia, X.; He, Z.; Liu, D.; Li, Z. QTL mapping of adult plant resistance to stripe rust and leaf rust in a Fuyu 3/Zhengzhou 5389 wheat population. Ther. Crop J. 2020, 8, 655–665. [Google Scholar] [CrossRef]
- The International Wheat Genome Sequencing Consortium. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ data to high-confidence variant calls: The genome analysis toolkit best practices pipeline. Curr. Bioinform. 2013, 43, 11101–111033. [Google Scholar]
- Mansfeld, B.N.; Grumet, R. QTLseqr: An R package for bulk segregant analysis with next-generation sequencing. Plant Genome 2018, 11, 180006. [Google Scholar] [CrossRef] [Green Version]
- Imerovski, I.; Dedić, B.; Cvejić, S.; Miladinović, D.; Jocić, S.; Owens, G.L.; Tubić, N.K.; Rieseberg, L.H. BSA-seq mapping reveals major QTL for broomrape resistance in four sunflower lines. Mol. Breed. 2019, 39, 41. [Google Scholar] [CrossRef]
- Konieczny, A.; Ausubel, F.M. A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specifc PCR-based markers. Plant J. 1993, 4, 403–410. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; William, M.; Rosewarne, G.; Djurle, A.; Yuen, J. Identification and mapping of Lr3 and a linked leaf rust resistance gene in durum wheat. Crop Sci. 2007, 47, 1459–1466. [Google Scholar] [CrossRef] [Green Version]
- Schachermayr, G.; Feuillet, C.; Keller, B. Molecular markers for the detection of the wheat leaf rust resistance gene Lr10 in diverse genetic backgrounds. Mol. Breed. 1997, 3, 65–74. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Gao, J.; Bi, W.; Zhao, J.; Yu, X.; Li, Z.; Liu, D.; Liu, B.; Wang, X. Genome-wide expression profiling of genes associated with the Lr47-mediated wheat resistance to leaf rust (Puccinia triticina). Int. J. Mol. Sci. 2019, 20, 4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, I.S.; Sharma, A.; Kaur, S.; Nahar, N.; Bhardwaj, S.C.; Sharma, T.R.; Chhuneja, P. Comparative temporal transcriptome profiling of wheat near isogenic line carrying Lr57 under compatible and incompatible interactions. Front. Plant Sci. 2016, 7, 1943. [Google Scholar] [CrossRef] [Green Version]
- Haggag, M.E.A.; Dyck, P.L. The inheritance of leaf rust resistance in four common wheat varieties possessing genes at or near the Lr3 locus. Can. J. Genet. Cytol. 1973, 15, 127–134. [Google Scholar] [CrossRef]
- Sacco, F.; Suárez, E.Y.; Naranjo, T. Mapping of the leaf rust resistance gene Lr3 on chromosome 6B of Sinvalocho Ma wheat. Genome 1998, 41, 686–690. [Google Scholar] [CrossRef]
- Künzel, G.; Korzun, L.; Meister, A. Cytologically integrated physical Restriction Fragment Length Polymorphism maps for the barley genome based on translocation breakpoints. Genetics 2000, 154, 397–412. [Google Scholar] [CrossRef]
- Gebrewahid, T.W.; Yao, Z.J.; Yan, X.C.; Gao, P.; Li, Z.F. Identification of leaf rust resistance genes in Chinese common wheat cultivars. Plant Dis. 2017, 101, 1729–1737. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pt Pathotype | PHSS-1 | PHSS-2 | PHQS | FHSS | PHTS-1 | KHSS | PHTS-2 | TGTS | FHJQ | NHJS |
| Klein Proteo | 1 | 1 | 2 | 1 | 1 | 1+ | 1 | 1 | 1 | ; |
| Lr3a-NIL | 4 | 4 | 3 | 4 | 3+ | 4 | 4 | 3+ | 2 | 3 |
| Lr3bg-NIL | 3 | 3 | 3 | 4 | 3+ | 4 | 4 | 3 | 1+ | 3 |
| Lr3ka-NIL | 3+ | 3+ | 3 | 3+ | 3 | 3+ | 4 | 2 | 2 | 1 |
| Lr10-NIL | 3 | 3+ | 3 | 3 | 3+ | 4 | 4 | 4 | 3 | 3 |
| ZhengZhou 5389 | 4 | 4 | 3+ | 3+ | 4 | 3+ | 4 | 4 | 3+ | 3+ |
| Lr13-NIL | 3+ | 4 | 3+ | 4 | 3+ | 4 | 3 | 3+ | 3+ | 3 |
| Lr16-NIL | 3+ | 4 | 4 | 4 | 3 | 4 | 4 | 4 | 4 | 3 |
| Lr23-NIL | 3 | 3+ | 3 | 3+ | 3+ | 3+ | 4 | 4 | 4 | 3 |
| ZhouMai 22 (LrZH22/Lr13) | 3+ | 2 | 2 | 3 | 2 | 1 | ; | 2 | 4 | 3 |
| PtPathotype | FRJS | FGBS | PGJS | FHSS | THSP | SHJT | PHJS | FHGS | PHTT | FHGQ |
| Klein Proteo | ; | ; | ; | ; | 1 | 1 | ; | 1 | ; | ; |
| Lr3a-NIL | 3+ | 3+ | 3 | 3+ | 2 | 3 | 3 | 3 | 3 | 3 |
| Lr3bg-NIL | 3 | 3+ | 3 | 3 | 2 | 3 | 3 | 3 | 3 | 3+ |
| Lr3ka-NIL | 2 | 2 | 3c | 3 | 1 | 1 | 1 | 2 | 3 | 2 |
| Lr10-NIL | 3 | 3+ | 3 | 2 | 3 | 3+ | 3 | 3 | 3 | 3 |
| ZhengZhou 5389 | 3+ | 3+ | 4 | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ | 3+ |
| Lr13-NIL | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Lr16-NIL | 3 | 3+ | 3 | 3+ | 3+ | 3 | 3 | 3+ | 3 | 3+ |
| Lr23-NIL | 3 | 3+ | 3 | 2 | 2 | 1 | 3 | 3 | 3 | 3 |
| ZhouMai 22 (LrZH22/Lr13) | 1 | 2 | 1 | 3 | 3 | 3 | 1 | 2 | 2 | 1 |
| CAPS Markers | Primer Sequence (5′-3′) | SNP Location | Expected Size a | Annealing Temperature | Restriction Enzyme |
|---|---|---|---|---|---|
| Lrkp2B114 | CAAACCCTCACCTTGGAAGC | 114 Mb | 1021 bp | 58 °C | AatII |
| GCCCTGGAGGTTTTCACGA | |||||
| LrkpF299R300 | GTGCAGACGTTAGAGCGTAG | 168 Mb | 337 bp | 55 °C | TaqI |
| TGATGTACATTTTTGTGGGGAAT | |||||
| LrkpF286R286 | ATCCGCAGCAAGCACATAC | 209 Mb | 510 bp | 55 °C | MboII |
| ACACAAAGAGATTAGGGCGTGT | |||||
| Lrkp197F2R2 | AACTTCATGTACGCCCTGT | 210 Mb | 921 bp | 52 °C | NdeI |
| GTTGGTCACCTAAACTGCC | |||||
| Lrkp2b01F4R4 | ATGAACCCTCTCTGTGTTTGAGC | 240 Mb | 760 bp | 60 °C | MspI |
| TACTGTGCCGCAGTGTCGC | |||||
| Lrkp260F1R1 | GACACCTTAGCAGCTCCCTA | 260 Mb | 679 bp | 55 °C | HindIII |
| ATGTTTGCTACTTCTTCCGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, S.; Pang, S.; Li, H.; Yang, J.; Yu, H.; Chen, S.; Wang, X. Broad-Spectrum Resistance to Leaf Rust in the Argentinean Wheat Cultivar “Klein Proteo” Is Controlled by LrKP Located on Chromosome 2BS. Agriculture 2022, 12, 1836. https://doi.org/10.3390/agriculture12111836
Bai S, Pang S, Li H, Yang J, Yu H, Chen S, Wang X. Broad-Spectrum Resistance to Leaf Rust in the Argentinean Wheat Cultivar “Klein Proteo” Is Controlled by LrKP Located on Chromosome 2BS. Agriculture. 2022; 12(11):1836. https://doi.org/10.3390/agriculture12111836
Chicago/Turabian StyleBai, Shengsheng, Shuyong Pang, Hongna Li, Jinwei Yang, Haitao Yu, Shisheng Chen, and Xiaodong Wang. 2022. "Broad-Spectrum Resistance to Leaf Rust in the Argentinean Wheat Cultivar “Klein Proteo” Is Controlled by LrKP Located on Chromosome 2BS" Agriculture 12, no. 11: 1836. https://doi.org/10.3390/agriculture12111836
APA StyleBai, S., Pang, S., Li, H., Yang, J., Yu, H., Chen, S., & Wang, X. (2022). Broad-Spectrum Resistance to Leaf Rust in the Argentinean Wheat Cultivar “Klein Proteo” Is Controlled by LrKP Located on Chromosome 2BS. Agriculture, 12(11), 1836. https://doi.org/10.3390/agriculture12111836